氮沉降增加情景下植物-土壤-微生物交互對自然生態系統土壤有機碳的調控研究進展
2019-01-18 05:36:10程淑蘭方華軍于光夏曹子鋮
生態學報 2018年23期
程淑蘭,方華軍,*,徐 夢,耿 靜,何 舜,于光夏,曹子鋮
1 中國科學院地理科學與資源研究所/生態系統觀測與模擬重點實驗室,北京 100101 2 中國科學院大學資源與環境學院,北京 100049
當前全球碳循環研究中的一個關鍵科學問題是“失蹤碳匯”(missing C sink)的分布與驅動機制[1]。過去150年,人類活動導致大氣氮沉降增加了3倍[2],顯著提高了受氮限制的陸地生態系統碳儲量,是正確解釋“失蹤碳匯”的重要途徑[3]。然而,目前有關氮沉降驅動陸地生態系統固碳的效率以及時空格局還存在很大的不確定性,不同學者估計的“氮促碳匯”變化范圍為16—400 kgC/kgN[4- 11]。相對于植被碳匯,有關氮沉降增加情景下土壤碳儲量的演變方向也存在分歧,包括增加[12]、降低[13]和不變[14]等3種結論。增氮引起土壤碳匯增量變化范圍為0—70 kgC/kgN[7, 15- 17],同樣存在很大的不確定性。
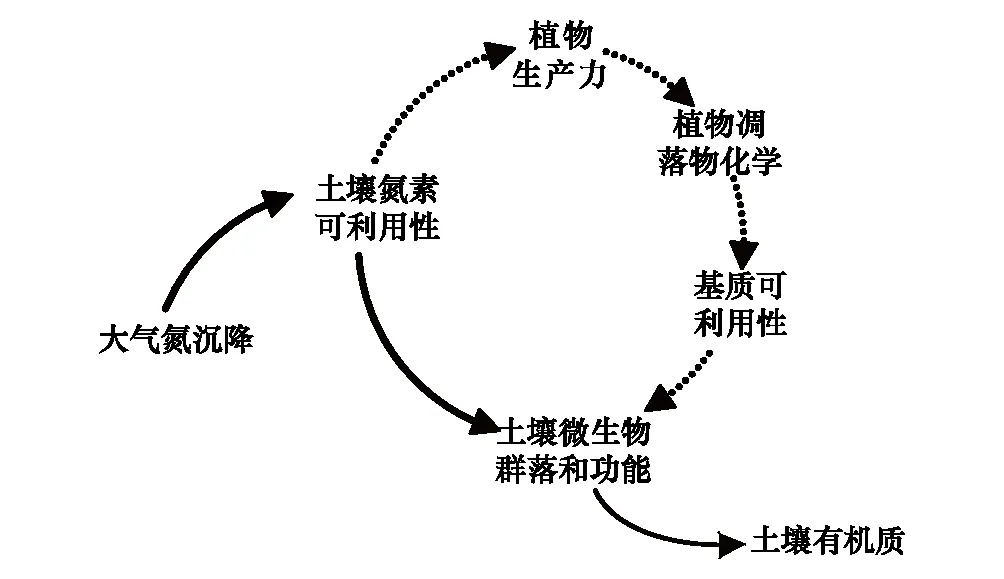
圖1 氮沉降對自然生態系統植物和微生物群落的影響 Fig.1 Effects of nitrogen deposition on plant and microbial communities in natural terrestrial ecosystems實線為直接影響,虛線為間接影響
大氣氮沉降輸入提高生態系統氮素的可利用性,直接改變土壤微生物群落的數量和組成,抑制根系和凋落物分解的速率和程度[18],進而增加或降低土壤有機質(SOM)的儲量和穩定性[19];另外,大氣氮沉降輸入提高植被生產力,改變凋落物產量和生化屬性[20],通過底物可利用性來調節土壤微生物群落的結構和功能[21],間接影響SOM的累積速率(圖1)。土壤有機碳(SOC)庫對增氮的響應取決于生態系統類型[22]、土壤深度[23]、SOC組成[24]、施氮類型和劑量[25]等要素。就植物-土壤-微生物群落系統而言,增氮會自上而下改變SOC的周轉過程與穩定性,理論上會產生以下4種影響:①外源性氮輸入會改變植物同化碳的再分配,根際沉積碳的數量和質量的變化進而影響活躍微生物群落組成[26];②氮素有效性增加會抑制微生物胞外酶活性,降低微生物群落獲取碳源的能力,提高其碳素利用效率[27];③氮素富集會改變微生物群落之間的交互作用和競爭關系,進而改變分解微生物群落的組成[28];④氮素富集會降低凋落物的分解速率和程度,增加類木質素化合物的氧化程度和穩定性,進而促進SOM的積累[29]。然而,上述4個科學假設缺乏系統的實驗驗證,其內在的微生物分子生態學機理未得到很好的理解,也沒有被融入到當前主流的生物地球化學模型之中[30]。
過去30年,盡管自然陸地生態系統對大氣氮沉降增加的響應研究得到了充分的重視,但是主要側重于生態系統碳氮轉化過程與土壤碳儲量動態,缺乏植物-土壤-微生物群落的交互作用及其調控機制的研究,對SOC積累與損耗的微生物學機理研究也不夠系統深入。探討植物-土壤-微生物群落三者交互作用及其對土壤碳儲量動態的調控作用,研究有助于完善陸地生態系統碳-氮循環耦合模型,深入認識SOC截存與損耗的機理,有效降低區域陸地“氮促碳匯”評估的不確定性,并可為陸地生態系統應對全球變化提供科學依據。鑒于此,本文以地下碳循環過程為主線,分別綜述氮富集對植物地下碳分配、SOC激發效應、微生物群落碳代謝過程的影響,深入分析SOM化學穩定性與微生物群落動態的關系。論述過程中我們試圖標注各個領域的薄弱環節,明確未來可能的研究重點,期望起到拋磚引玉的效果。
1 增氮對植物地下碳分配的影響
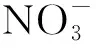
2 氮素有效性對SOC激發效應的影響
增加凋落物的輸入會降低土壤碳庫,該現象稱為“激發效應”[47]。激發效應是由于外源底物的添加激活土壤中的微生物,促進它們對SOM的分解。根據激發方向可分為正激發和負激發效應,根據有無有機質參與可分為表觀激發和真實激發[48]。當添加的外源底物遠低于微生物量時,由于微生物自身內部代謝和周轉導致CO2釋放的過程,并不涉及SOM周轉,因此表觀激發對土壤碳庫沒有影響。但是當添加的底物劑量遠高于微生物量時,微生物優先利用外源底物的能量,降低對SOM的分解,因此會出現負激發。隨后,當外源底物的能量耗盡之后,促使被激活的微生物分解SOM來獲取所需的養分和能量,從而產生正激發。基于全球培養實驗的集成分析,發現正激發效應可促使有機質分解增加 3.8倍,而負激發效應導致SOM分解減少50%[49]。激發效應的產生機制主要包括:①共代謝理論[50];②底物偏好理論[51];③氮挖掘理論[52];④化學計量學理論[53]等。正激發產生機理以微生物激活作用、微生物競爭與演替、養分挖掘和化學計量學為主,而負激發產生機理以底物偏好利用和毒害作用為主。微生物的種類和大小[51]、植物類型和物候[54]、土壤動物對微生物的取食[55- 56],均會影響激發效應的強度。非生物因素如底物質量[57]、數量[58]和土壤屬性(溫度、水分、土層、養分狀態)也會影響微生物群落對底物的利用,進而影響激發效應的強度和方向[59-62]。
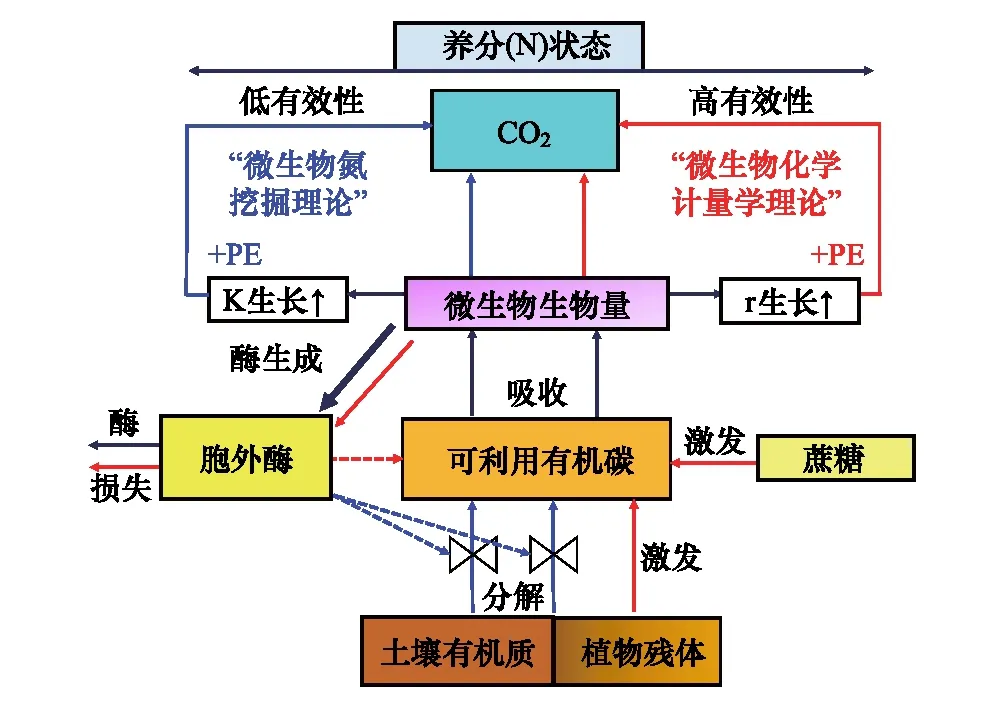
圖2 養分挖掘理論與化學計量學理論關系[53] Fig.2 The relationship between nutrient mining theory and stoichiometry theory[53]PE: 激發效應priming effect

3 增氮對微生物群落碳代謝的影響
微生物碳利用效率(CUE)是指分配給微生物生長所用有機碳與微生物群落組成所吸收的有機碳之比,它是微生物群落碳代謝的一個重要的綜合指標[67]。理論上,微生物CUE被熱力學限制在0.88[68],即微生物吸收的碳用于生長的部分不會超過88%,因為微生物至少需要12%的碳用于呼吸產生能量以維持其自身的生物量。然而,由于環境條件的限制,微生物生長和生物量的維持需要超過其最小的能量投入,土壤微生物CUE未達到理論上的最大值[69]。土壤生源要素化學計量學、底物質量、微生物群落組成均會影響微生物CUE。在養分限制條件下(高碳/養分比),微生物被迫分配較多的碳和能量以獲取所需養分,土壤呼吸CO2排放較高,導致微生物CUE較低[67, 69]。當外源性底物的C∶N∶P比與微生物生物量C∶N∶P比(60∶7∶1—42∶6∶1)差異較大時,微生物必須分配更多能量生產胞外酶來獲取缺失元素以維持其生物量;同時更多同化碳被用于微生物呼吸,導致微生物CUE下降[67, 69]。因此,養分富集農田土壤的微生物(尤其是細菌)生長效率普遍高于受氮限制的森林和草地土壤[70]。此外,碳和養分的可利用性也會影響微生物生物量的周轉[71]。微生物CUE和土壤微生物生物量周轉時間可能受有機碳濃度和質量的影響。降解復雜化合物需要多種胞外酶,而酶的合成需要消耗大量的能量和氮,所以降解復雜底物的微生物CUE較低[72]。由于微生物通過不同的代謝路徑來同化不同類型底物,導致單位質量的同化碳具有不同的呼吸速率[73]。因此,植物類型、微生物群落組成以及不同來源的凋落物也會間接地影響微生物CUE,真菌的CUE一般高于細菌[74]。目前測定微生物CUE的方法主要是基于13C標記底物,然而不同底物的微生物CUE差異巨大,該方法混淆了微生物碳利用效率與特定底物的利用效率[75]。最近,Spohn等[76]建立了基于18O-H2O標記技術計算微生物CUE的新方法,根據18O-DNA的增加量來計算土壤微生物的生長速率,再結合基礎呼吸速率和微生物數量,可計算出微生物CUE和周轉時間。
野外觀測和模型模擬結果表明,施氮降低微生物活性、SOM分解和CO2排放[77- 80]。施氮可能通過改變微生物群落活性和組成[81- 82]以及底物的化學性質(如可分解性)來抑制土壤微生物呼吸,與微生物CUE、胞外酶活性以及微生物生物量的變化有關[27]。其理論假設是:(1) 氮限制條件下,增氮會導致更多的碳向微生物分配,而不是通過呼吸和胞外酶損失,導致微生物CUE增加,呼吸降低[69],這可能與微生物群落組成向高CUE轉變有關[83]。(2)增氮抑制了降解復雜底物(如木質素)的氧化酶活性。例如,增氮降低了北美溫帶森林土壤酚氧化酶活性[22]、木質素降解相關的功能基因豐度和基因表達[84]。(3)增氮通過減少分解微生物的生物量而降低呼吸[33]。增氮導致土壤酸化,增加Ca2+、Mg2+等陽離子的溶解與流失,土壤溶液中Al3+濃度增加,對微生物產生毒害作用[85]。此外,土壤酸化也會抑制微生物胞外酶活性,金屬離子的鍵合增加有機質中碳的穩定性,降低微生物獲取碳的能力,進而降低微生物生物量和分解速率[86- 87]。盡管已經對氮素富集條件下土壤微生物碳的代謝過程和響應格局做了一些探討,但是氮素類型和劑量如何差異性地影響微生物碳代謝功能還不得而知;由于缺乏合適的測定方法,氮沉降增加情景下自然陸地生態系統土壤微生物碳利用效率及微生物生物量的周轉時間尚未得到準確的量化。
4 SOM化學穩定性與微生物群落動態的關系

有機質化學結構的變化也會影響土壤微生物的底物利用,進而影響微生物群落的活性和組成,反之亦然。長期施氮會顯著降低森林、草地等自然生態系統土壤微生物生物量[34],抑制腐生菌分泌木質纖維素水解酶,降低腐生微生物群落獲取碳源(如纖維素、半纖維素)的能力[93];此外,氮沉降輸入還會抑制白腐菌產生酚氧化酶,進而抑制木質素溶解酶生成,促進耐分解有機質的積累[94]。利用功能基因定量PCR和高通量測序技術,Hassett等[95]和Edwards等[84]研究發現,施氮傾向于降低美國密歇根糖楓林土壤擔子菌漆酶基因lcc的拷貝數(lcc編碼酚氧化酶),改變真菌群落組成(擔子菌OTUs/子囊菌OTUs增加),并且隨著施氮時間的延長其抑制效應更加顯著。同時,施氮傾向于降低凋落物分解速率和程度,增加類木質素化合物的氧化度和穩定性,進而促進SOM的積累[29]。潛在的生物化學機理是:高劑量的無機氮會抑制某些擔子菌木質素水解酶的合成,以及編碼酚氧化酶、錳過氧化物酶和木質素過氧化物酶的真菌基因轉錄下調,導致木質素降解下降[96]。木質纖維素水解酶基因轉錄下調會導致凋落物分解下降和SOM積累[97]。總體上,過去對氮沉降增加情景下森林土壤真菌功能基因表達和氧化酶活性演變研究較為深入,但是有關草地土壤細菌和真菌群落組成的變化如何代謝有機底物的機制仍不清楚,土壤微生物群落組成、SOM化學結構的變化與SOC積累之間的耦聯關系尚不清晰。
5 存在問題與展望
目前關于外源性氮輸入對自然生態系統土壤碳動態的影響已開展了一些探索,但總體上還不夠深入,存在許多問題亟待解決。首先,植物根系碳輸入對增氮的響應呈現拋物線變化,取決于生態系統氮素有效性、施氮的類型與持續時間。總的來說,施氮降低根系的生長與周轉,但是根際沉積物對增氮的響應及其固碳效應十分復雜,與植被-菌根類型、生態化學計量學等要素有關,未形成統一的研究結論。其次,關于激發效應的3個階段、4種驅動機制及影響因素分析較為深入,認識到養分尤其是氮素強烈影響激發效應的方向及強度,但是氮素類型、土層深度及其交互作用對激發效應的貢獻并不清楚。再次,微生物CUE是微生物群落碳代謝的關鍵表征,受底物生態化學計量學驅動,能夠很好地解釋氮沉降增加情景下土壤碳的積累與損耗過程;由于缺乏適宜的測定方法,目前還難以準確量化土壤微生物CUE及微生物生物量的周轉時間。最后,SOM化學質量與土壤微生物群落活性、組成之間存在復雜的交互作用,增氮顯著抑制貧氮生態系統土壤真菌群落及其胞外酶活性,對細菌群落組成的影響結論多樣,有關微生物群落代謝底物、SOC積累的生物化學機制尚不清楚。

圖3 氮沉降增加情景下植物-土壤-微生物交互作用對土壤有機碳動態的調控機制Fig.3 Mechanisms responsible for the regulation of plant-soil-microbe interactions to soil organic carbon dynamics under elevated nitrogen deposition
6 結論
外源性氮輸入通過影響微生物群落活性和組成直接影響SOM分解,或通過改變底物數量和質量間接影響SOC的分解和累積過程。總體上,增氮傾向于促進地上生物量,抑制地下碳分配和根系周轉。氮素有效性強烈影響SOM激發效應的方向和強度,取決于微生物生長的r-K策略和生態化學計量學特征。微生物CUE是衡量微生物群落碳代謝的重要表征,能夠很好地解釋土壤碳的積累與損耗過程,目前缺乏準確測定CUE的方法,穩定性碳氧同位素示蹤是解決該問題較為理想的手段。SOM化學結構與微生物群落活性、組成緊密耦合在一起,增氮抑制真菌群落及氧化酶活性,對細菌群落組成的影響尚未形成定論,需要將有機質化學與分子生物學、宏基因組學方法緊密結合起來,深入地揭示增氮引起土壤碳積累與損耗的生物化學與微生物學機理。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
核科學與工程(2021年4期)2022-01-12 06:30:26
當代陜西(2021年2期)2021-03-29 07:41:24
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學物理·高中(2016年12期)2017-04-22 11:53:03
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
