荒漠齒肋赤蘚(Syntrichia caninervis)非結構性碳水化合物含量對植株脫水的響應
2019-01-18 05:36:16李永剛張元明
生態學報 2018年23期
李永剛,張元明,*
1 中國科學院新疆生態與地理研究所干旱區生物地理與生物資源重點實驗室, 烏魯木齊 830000 2 中國科學院大學, 北京 100049
生物土壤結皮(Biological Soil Crusts, BSCs)是由隱花植物如藍藻、荒漠藻、地衣、苔蘚和土壤中微生物,以及相關的其他生物體通過菌絲體、假根和分泌物等與土壤表層顆粒膠結形成的十分復雜的復合體,是干旱半干旱荒漠地表景觀的重要組成之一[1]。生物土壤結皮對荒漠的穩定和土壤微環境的改善具有重要作用[2-3]。據生物土壤結皮的演替順序,大致可分為藻結皮、地衣結皮和苔蘚結皮,而苔蘚結皮既是生物結皮演替的最高階段也是結皮生物量的最主要貢獻者,對土壤水分的維持和群落的演替的具有重要影響[4]。苔蘚植物作為植物演替的先鋒物種,能夠在極端高、低溫、干燥、強輻射等條件下生存,因此具有很強的抗脅迫特性[5]。為此,諸多研究對其在脅迫條件下的形態特征[6- 11]、生理生化特性[12- 19]、分子表達等方面做了詳細研究[20- 22]。然而,對蘚類植物體內的非結構性碳水化合物變化的相關研究還相對較少。
非結構性碳水化合物包括可溶性非結構性碳水化合物和不溶性非結構性碳水化合物[23],可溶性非結構性碳水化合物又包括蔗糖、葡萄糖、果糖和山梨醇等。不可溶性非結構碳水化合物主要是指淀粉[23-24]。可溶性糖是植物生長發育的主要物質,能夠為植物的生長發育提供能量和中間產物;其中蔗糖是植物體內主要的運輸物質,并且可以作為信號物質用來調節植物細胞內基因的表達,對細胞的代謝進行調節;果聚糖是植物營養組織主要的暫貯形式,能夠短時間貯藏有機物供給營養組織;然而淀粉是植物的長期貯存物質[25]。非結構性碳水化合物既是植物光合作用的直接儲存物質,也是植物生長和發育的重要調節物質,能夠隨環境(如水分、溫度、大氣、二氧化碳和土壤養分)變化而變化[26-28]。非結構性碳水化合物含量的變化極有可能是植物對環境變化產生的信號,從而調節體內基因的表達,有效的減緩環境脅迫對植物體造成的損傷,進而調節植物在環境改變時的生長發育。例如,劉香萍等[29]對紫花苜蓿的抗寒抗旱研究中發現,秋季紫花苜蓿主根內的可溶性碳水化合物的變化對抗寒性和再生率有著重要作用。許志信和白永飛[25]對草原碳水化合物的儲藏部位以及對植物不同生長發育時期植物非結構性碳水化合物儲存的位置和含量的變化與環境相關。對于耐旱植物非結構性碳水化合物的研究表明,耐旱植物體內非結構性碳水化合物含量的變化和環境與水分脅迫改變有關[5]。
干旱半干旱地區降水較少,且多以脈沖形式降雨[30],荒漠蘚類植物易受復水-脫水循環的影響,對水分變化十分敏感的荒漠蘚類植物,在形態和生理上必然會表現出一定的響應特征。然而,有關荒漠蘚類植物體內非結構性碳水化合物的響應特征尚無相關報道。在古爾班通古特沙漠腹地和南緣年降雨量不同,南緣降雨量較高,苔蘚植物對水分變化極為敏感,降雨量的不同對苔蘚非結構性碳水化合物的影響鮮有報道。因此,我們選取古爾班通古特沙漠優勢蘚類植物齒肋赤蘚(Syntrichiacaninervis),對其在復水后脫水48 h內齒肋赤蘚非結構性碳水化合物的變化進行研究,以期了解齒肋赤蘚在脈沖式降雨后非結構性碳水化合物變化規律,為深入理解荒漠蘚類植物對干旱環境的適應機制提供資料。
1 材料與方法
1.1 研究區概況
研究區位于新疆準噶爾盆地中部的古爾班通古特沙漠(44°11′—46°20′ N, 84°31′—90°00′ E, 海拔300—600 m)。古爾班你通古特沙漠面積為4.88×104km2,是中國最大的固定和半固定的沙漠。該沙漠的基地是第四世紀疏松沖積物,主要有0.1—0.25 mm粒級的沙粒組成。由于受到喜馬拉雅山脈的阻隔作用,來自印度洋西南季風的水汽未能進入到內陸,導致該沙漠屬于典型的溫帶大陸性氣候。年均降水量僅70—160 mm,潛在年蒸發量在2606.6 mm左右,年均溫7.26℃,冬季平均氣溫在-16.8℃。該沙漠在冬季具有穩定的積雪覆蓋,平均積雪厚度達到20 cm左右,占全年降水量的25%以上[31-32]。
該沙漠植被以白梭梭(Haloxylonpersicum) 和梭梭(Haloxylonammodendron)等小半喬木為建群種。沙丘中下部有蛇麻黃(Ephedradistachya)、淡枝沙拐棗(Calligonumleucocladum)等灌木廣泛分布[33],春季融雪使得短命類短命植物及一年生草本廣泛發育。此外,該沙漠地表生物土壤結皮發育良好,主要包括藻結皮,地衣結皮和苔蘚結皮3種類型。藻結皮主要分布在沙壟中上部,地衣和苔蘚結皮主要分布在下坡和丘間低地,尤其在蛇麻黃、沙拐棗等灌叢下發育良好[33]。古爾班通古特沙漠南緣地貌類型以南北走向的樹枝狀沙壟為主,表現為固定、半固定沙丘,其間廣泛分布著由梭梭、生物結皮、以及其他沙生植物組成的小喬木群落,梭梭是建群種。平均氣溫7.19℃,極端最高氣溫41.5℃。多年平均降水量128.6 mm,潛在蒸發量為1746.4 mm。春夏降雨量約占總降水量的2/3,秋季干燥,冬季積雪厚度一般在20 cm以上,積雪覆蓋持續100—160 d,春季的覆雪融水和春夏季的降雨是維持本區植被生存的主要水分來源[34]。古爾班通古特沙漠腹地風沙地貌類型以固定、半固定沙壟為主,沙壟頂部普遍存在10—40 m寬度不等的流動帶[35],平均氣溫5—5.7℃,極端最高氣溫40℃以上, 極端最低氣溫小于-40℃,年降雨量70—160 mm,年蒸發量2000—2800 mm,冬季穩定積雪厚度在20 cm左右。沙漠腹地較沙漠南緣更為干旱。沙漠南緣齒肋赤蘚蓋度較高于腹地。
1.2 研究對象
齒肋赤蘚(S.caninervis)是古爾班通古特沙漠苔蘚結皮的優勢種,屬于叢蘚科赤蘚屬,具有良好的耐旱性和變水性。在高溫干燥的情況下,通常失水并呈現出黑色或黑褐色,處于休眠狀態。復水后快速變為綠色,并在短時間內恢復光合作用[36]。
1.3 樣品的采集及處理
樣品采集于古爾班通古特沙漠腹地(45°24′N,87°31′E,海拔478 m)和南緣(44°36′N,88°15′E,海拔478 m)兩個區域,在區域內隨機選擇發育良好的齒肋赤蘚結皮,用自制PVC管(內徑10 cm,高5 cm)垂直插入沙土中,小心取得未擾動的苔蘚結皮樣品,每個區域取40個,封底后迅速運至實驗室。由于6 mm和10 mm降雨后苔蘚含水量無差異,且6 mm降雨時苔蘚光合色素含量最高,能夠恢復其生理活性[36-37],因此齒肋赤蘚復水時加入6 mm降雨量的蒸餾水,通過PVC管的面積大小計算每個PVC管中應加入蒸餾水的量為47.1 mL(計算公式: PVC管的面積A=πr2,降雨量:V=A×h,r=5 cm,h=0.6 cm,計算后V=47.1,即47.1 mL)。用小型噴壺向PVC管中的齒肋赤蘚表面噴灑47.1 mL的蒸餾水。為研究齒肋赤蘚在脫水過程中不同時間的非結構碳水化合物含量的變化,以0 h(噴水前)為對照,在噴水后1、2、4、8、16、24 h和48 h進行取樣,每個處理取5個重復。在不同時間節點,用刀片快速切取苔蘚莖葉樣品,過篩3次以去除樣品中夾雜的沙粒,稱取150 mg鮮重(0.0001 g精度天平)樣品,用錫箔紙包裹后用液氮快速冷凍,然后放置于-80℃冰箱備用。
1.4 含水量的測定
分別對齒肋赤蘚復水前(0 h)和脫水不同階段(1、2、4、8、16、24 h和48 h)的含水量進行測定,并在不同時間點稱取0.5 g地上部分鮮樣,將稱好的鮮樣放進烘箱先105℃殺青,然后65℃下48 h烘干、稱干重,含水量=(鮮重-干重)/鮮重。
1.5 非結構性碳水化合物的測定
樣品的非結構性碳水化合物測定參照上海植物生理學會(1985)著《植物生理實驗手冊》。將樣品用粉碎機粉碎,加7 mL 70%乙醇提取10 min,4℃下8000g離心10 min,收集上清液于100 mL容量瓶中,反復3次,合并上清液,用70%乙醇定容,即為可溶性糖提取液。
1.5.1 可溶性總糖含量測定
取1 mL可溶性糖提取液,加入5 mL蒽酮試劑,90℃水浴鍋中反應15 min,待冷卻后于620 nm下讀取吸光度值[38]。根據標準曲線可以計算其濃度,為保證不同含水量間濃度的比較,文中濃度均計算為mg/g干重。
1.5.2 蔗糖含量測定
取1 mL可溶性糖提取液,加入7.6 mol/L KOH 0.1 mL,沸水浴10 min,冷卻后加入5 mL蒽酮試劑,90℃保溫15 min,待冷卻后于620 nm下讀取吸光度值[36]。根據標準曲線可以計算其濃度。
1.5.3 果糖含量測定
取1 mL可溶性糖提取液,加入5 mL蒽酮試劑,25℃反應90 min后,于620 nm下比色,根據果糖標準曲線,計算植物組織提取液中的果糖含量。
1.5.4 淀粉含量的測定
采用高氯酸法,用上述離心后的沉淀物測定植物樣品中的淀粉含量,沉淀物加入30% (v/v)的高氯酸8.5 mL,靜置過夜,然后于80℃水浴中提取10 min,冷卻,4000 r/min離心10 min,取上清液定容到100 mL的容量瓶中,取1 mL提取液,于620 nm下測量吸光度值。
1.6 數據處理
利用SPSS 22.0對所測得的數據進行正態性和方差齊性檢驗,并對不同時間點的齒肋赤蘚可溶性總糖、蔗糖、果糖和淀粉等含量進行單因素方差分析和多重比較,用ORIGIN 9.0作圖。
2 結果分析
2.1 脫水過程中齒肋赤蘚含水量隨時間的變化特征
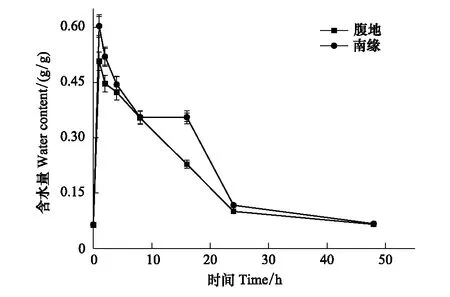
圖1 復水后齒肋赤蘚含水量隨時間的變化 Fig.1 Changes of water content in S. caninervis shoots after rehydration
由圖1可知,植株含水量在復水后1 h內急劇上升并達到最大,采自不同區域的苔蘚植株含水量分別從0.0635 mg/g和0.0645 mg/g上升到0.5112 mg/g和0.6023 mg/g。隨后在1到2 h呈現加速下降趨勢,下降速度分別為0.0617 mg g-1h-1和0.0793 mg g-1h-1,隨后在2—24 h之間含水量繼續緩慢下降,下降速率分別為0.0158 mg g-1h-1和0.0185 mg g-1h-1。不同區域對脫水過程植物含水量具有相似變化,脫水1、2、4、8 h和16 h與復水前相比差異極顯著(P<0.01),脫水24 h與復水前差異顯著(P<0.05),脫水48 h后與復水前含水量無差異(P=0.426)。復水前不同區域齒肋赤蘚植物含水量無差異,復水后在最大含水量時出現極顯著性 (P<0.05) 差異,在脫水48 h后含水量在不同區域上無顯著差異。
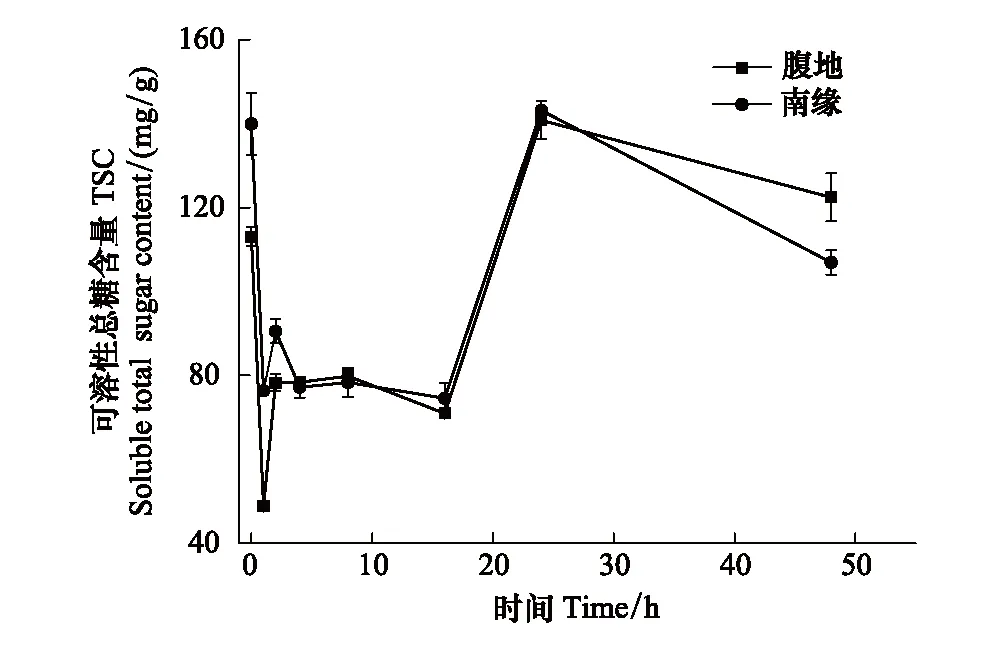
圖2 可溶性糖隨脫水時間的變化Fig.2 The TSC changes with time after the rain
2.2 脫水過程中齒肋赤蘚植株可溶性總糖、蔗糖和果糖的變化特征
脫水過程不同區域齒肋赤蘚植株可溶性總糖含量隨時間呈現下降、穩定、上升、下降四個階段的變化。復水后1 h齒肋赤蘚植株含水量達到最大值(圖1),從圖2中可知,在復水后1 h內齒肋赤蘚可溶性總糖含量快速下降,腹地和南緣的齒肋赤蘚可溶性總糖含量下降速率分別為64.275 mg g-1h-1和63.573 mg g-1h-1;其中腹地的齒肋赤蘚可溶性總糖含量下降到最低值,南緣的齒肋赤蘚可溶性總糖含量下降未達到整個脫水過程的最低值,且兩個地區齒肋赤蘚可溶性總糖含量存在顯著性差異(P<0.05)。不同區域的齒肋赤蘚蔗糖與果糖含量的變化趨勢與可溶性總糖相似,腹地中齒肋赤蘚植株蔗糖和果糖含量達到最小值,南緣齒肋赤蘚植株蔗糖和果糖含量未達到最小值,且蔗糖和果糖含量分別在不同區域之間存在顯著的差異(圖3,圖4)。
脫水過程中隨著含水量的逐漸降低,在2—16 h期間,沙漠腹地齒肋赤蘚在2—8 h無顯著變化,在8—16 h可溶性總糖含量出現下降趨勢,脫水8 h與16 h時齒肋赤蘚植株可溶性總糖含量出現顯著性差異(P<0.05),沙漠南緣齒肋赤蘚可溶性糖含量在2—4 h有下降的趨勢,脫水2 h與4 h齒肋赤蘚植株可溶性總糖含量存在顯著性差異,4 h后趨于穩定,且在脫水16 h時可溶性糖含量達到最小值。然而在脫水2—16 h期間不同區域齒肋赤蘚蔗糖和果糖含量也出現不同的變化趨勢,其中沙漠腹地齒肋赤蘚蔗糖含量出現先降低后穩定的變化,果糖含量表現出先升高后降低的趨勢,沙漠南緣齒肋赤蘚蔗糖含量變化趨勢與沙漠腹地不同,表現出先降低再升高最后趨于穩定,且在脫水4 h時達到蔗糖含量的最低值,果糖含量與腹地齒肋赤蘚果糖含量變化趨勢相反,出現先降低后穩定,且在脫水16 h時果糖含量達到最低值。
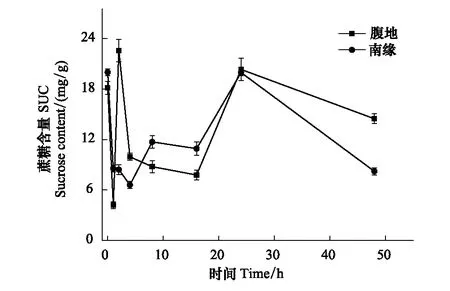
圖3 蔗糖含量隨脫水時間的變化Fig.3 The SUC changes with time
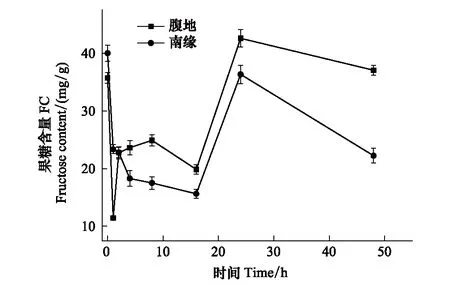
圖4 果糖含量隨脫水時間的變化Fig.4 The FC changes with time
脫水16 h,沙漠腹地和南緣齒肋赤蘚含水量分別下降到22.83%和23.35%。此后,在脫水16—24 h期間不同區域齒肋赤蘚可溶性總糖含量出現極顯著上升,腹地和南緣齒肋赤蘚可溶性總糖含量的上升速率分別是8.729 mg g-1h-1和8.36 mg g-1h-1,脫水24 h時齒肋赤蘚可溶性總糖,蔗糖含量在兩個區域無顯著差異(P=0.651),但果糖含量在兩個區域之間存在顯著的差異(P<0.05)。
脫水24 h,齒肋赤蘚含水量分別下降到10%和11%,脫水48 h后不同區域齒肋赤蘚含水量與復水前含水量無差異(腹地P=0.672和南緣P=0.426),脫水24—48 h齒肋赤蘚可溶性總糖含量再次緩慢下降,速率分別是0.67 mg g-1h-1和1.46 mg g-1h-1(圖2)。下降后齒肋赤蘚可溶性總糖和蔗糖含量在腹地和南緣兩個地區間存在顯著的不同(P<0.05),南緣齒肋赤蘚含量在脫水結束后可溶性總糖,蔗糖,果糖含量顯著低于復水前它們的含量。果糖的含量在不同區域之間存在極顯著的差異(P<0.01)。在含水量最高的時候蔗糖含量達到最低值,隨后再2—16 h趨于穩定狀態,16—24 h出現較為迅速的積累,24—48 h出現最為緩慢的下降。
2.3 脫水過程中淀粉在齒肋赤蘚植株中淀粉含量的變化特征。
從圖5中我們可以看出兩地點的苔蘚的淀粉含量的變化趨勢完全相同,復水前不同區域齒肋赤蘚淀粉含量存在顯著性差異,在復水后1 h內淀粉含量出現極顯著的下降,下降速率分別為腹地和南緣分別為37.859 mg g-1h-1和55.305 mg g-1h-1,此時兩個區域齒肋赤蘚淀粉含量達到最小值;脫水2 h時淀粉含量顯著增加,隨后在脫水2—16 h期間淀粉含量緩慢降低,腹地和南緣下降速度分別為1.55 mg g-1h-1和1.75 mg g-1h-1;脫水16—24 h淀粉含量顯著增加,增長速率分別是腹地5.24 mg g-1h-1和南緣7.63 mg g-1h-1,且在24 h時淀粉含量達到最高;在24—48 h,沙漠腹地齒肋赤蘚淀粉含量趨于穩定,沙漠南緣齒肋赤蘚含量出現緩慢的下降,下降速率為1.22 mg g-1h-1,在脫水24 h和48 h 之間存在顯著性差異。在脫水48 h后齒肋赤蘚的淀粉含量在南緣仍顯著高于腹地。脫水過程中南緣齒肋赤蘚淀粉含量在各個時段都顯著高于腹地齒肋赤蘚淀粉含量。
2.4 脫水過程中齒肋赤蘚植株非結構性碳水化合物含量的變化特征。
非結構性碳水化合物包括淀粉和可溶性糖,脫水過程中非結構性碳水化合物的變化趨勢與淀粉相似。如圖6所示脫水1 h非結構性碳水化合物含量出現極顯著降低,腹地和南緣的下降速率分別為102.13 mg g-1h-1和118.88 mg g-1h-1,兩個區域的齒肋赤蘚非結構性碳水化合物含量下降至最低;在脫水1—2 h齒肋赤蘚植株非結構性碳水化合物含量顯著的增加,增加速率分別為腹地73.825 mg g-1h-1和南緣60.545 mg g-1h-1;2—16 h處于穩定階段;16—24 h出現顯著的增加,腹地和南緣增加速率分別為13.94 mg g-1h-1和15.99 mg g-1h-1,在24 h非結構性碳水化合物含量達到最高;24—48 h非結構性碳水化合物緩慢下降,腹地和南緣的下降速率分別為1.34 mg g-1h-1和2.69 mg g-1h-1。在脫水48 h期間,沙漠南緣的齒肋赤蘚非結構性碳水化合物含量顯著高于腹地。
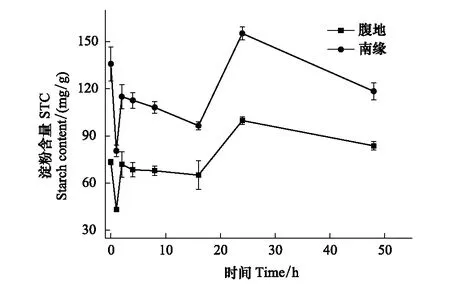
圖5 淀粉含量隨脫水時間的變化Fig.5 The STC changes with time
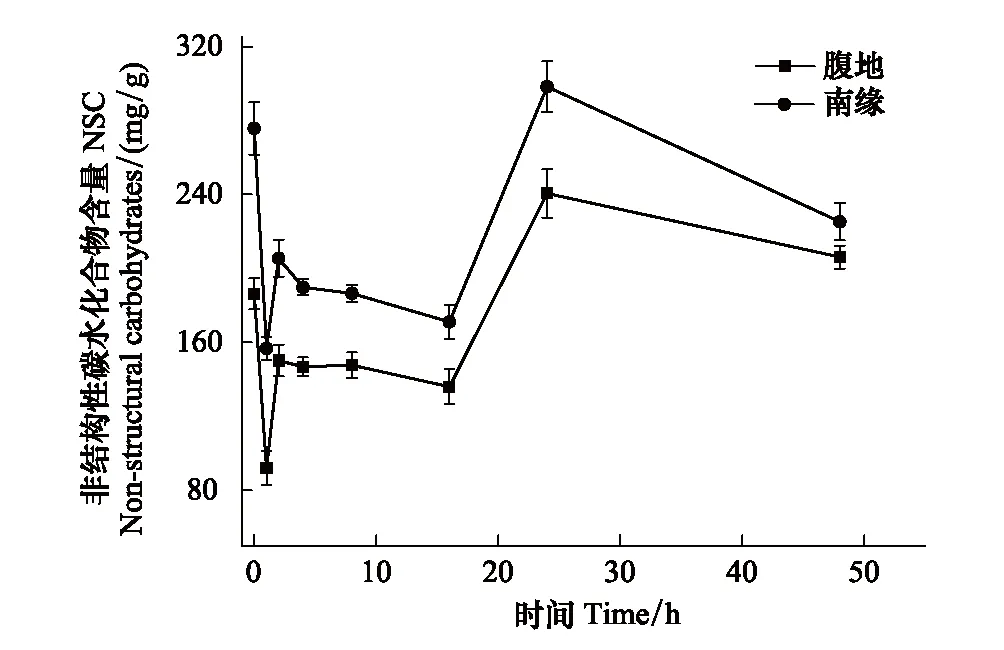
圖6 非結構性碳水化合物隨時間的變化 Fig.6 The changes in Non-structural carbohydrates with time increase
3 討論
3.1 脫水過程中齒肋赤蘚非結構性碳水化合物變化與水分脅迫的關系
高等植物碳水化合物代謝對干旱脅迫十分敏感,水分的脅迫會直接影響植物碳水化合物代謝,從而引起植物生長和代謝的改變[39]。齒肋赤蘚復水前處于休眠狀態,休眠期齒肋赤蘚具有較高的非結構性碳水化合物含量,能為復水后齒肋赤蘚的生長提供碳源,而且能為植物光合作用不足時起到緩沖作用,這與Hoch對森林生態系統樹木的研究結果相似[40]。復水后齒肋赤蘚迅速恢復活性,非結構性碳水化合物迅速作為碳源供齒肋赤蘚生長和代謝,非結構性碳水化合物轉化為代謝物質供齒肋赤蘚的生長,降低非結構性碳水化合物含量維持細胞吸水調節光合作用。脫水過程后期齒肋赤蘚受到干旱脅迫時,非結構性碳水化合物含量迅速增加,這一現象與小麥受到干旱脅迫,小麥葉片細胞可溶性糖迅速積累,維持滲透壓、降低水勢、減少葉片水相似[41-42]。齒肋赤蘚在受到干旱脅迫時過河作用降低,可溶性糖和淀粉的含量明顯上升,用于提高齒肋赤蘚滲透壓以及儲存碳源,隨著脫水過程的持續,齒肋赤蘚將受到極端干旱的脅迫,與維管植物不同齒肋赤蘚可以直接進入休眠狀態,維持較高的可溶性糖和淀粉的含量[43]。
3.2 脫水過程中齒肋赤蘚非結構性碳水化合物的積累與光合作用的關系
前期研究發現齒肋赤蘚在干旱情況下,往往處于休眠狀態,葉片背卷,緊貼于莖,呈黑褐色;當環境中有可利用的水分時,齒肋赤蘚能夠快速恢復其細胞膜的選擇吸收功能,迅速吸水展開葉片,恢復其正常的生理代謝功能。蘚類植物能夠在30—90 s內完成復水過程,數分鐘內激活其光合活性[44-45]。我們研究發現齒肋赤蘚在復水1 h后含水量達到最大值,且蔗糖、果糖、淀粉含量出現極顯著的下降,說明齒肋赤蘚非結構性碳水化合物的積累不在脫水早期。復水后光合作用能在40 min中內恢復到原來的90%以上,在光合作用未完全恢復時,呼吸消耗大量碳水化合物,保證其正常代謝功能。含水量在20%—70%之間,暗呼吸隨水分含量的下降而下降,并且一般認為呼吸速率恢復快于凈光合速率[45-46]。在1—16 h蔗糖果糖含量上升后處于一個相對穩定非結構性碳水化合物含量。1—2 h蘚類植物隨著光合作用的恢復,產生大量糖等有機物,此時可溶性糖和淀粉的含量增加。隨后可溶性總糖和淀粉含量處于相對穩定時期。隨著脫水過程光合色素含量在脫水2 h后迅速增加,脫水6 h達到最大,脫水10 h至完全干燥時,光合色素含量緩慢降低[47-48],同時含水量在20%—70%之間,光合作用不斷減弱,呼吸作用和光合作用變化趨勢相似,也呈現下降趨勢[49]。光合作用和呼吸作用同時下降可能導致可溶性糖和淀粉的含量處于相對穩定的狀態。在16 h后可溶性總糖和淀粉出現顯著性積累,然后緩慢降低,表明齒肋赤蘚非結構性碳水化合物的積累主要發生在16 h至24 h之間,16 h時齒肋赤蘚含水量下降到20%,齒肋赤蘚受到干旱脅迫,非結構性碳水化合物含量增加。國春暉[50]研究也發現脫水過程中刺葉赤蘚葉片可溶性糖持續積累,脫水24 h 時葉片可溶性糖含量已接近未脫水時的4倍。非結構性碳水化合物是植物典型的滲透調節物質,非結構化合物含量的改變可以直接影響植物滲透調節。植物細胞通過滲透調節能增加細胞溶質濃度,降低滲透勢,保持膨壓,緩和脫水脅迫,有助于光合作用、氣孔運動和細胞生長等生理過程正常進行[51-53]。徐杰等[54]也認為蘚類植物體內可溶性糖含量能夠對干旱脅迫產生應激反應,它能有效地保護細胞膜和蛋白質的結構,使生物體在許多異常情況下,如高溫、脫水、冷凍時仍保持細胞內濕潤,防止細胞因失水而造成細胞內養分的損失和細胞損傷,從而使這些生物具有較強的抗旱、抗寒的能力。在24 h后齒肋赤蘚非結構性碳水化合物出現緩慢的下降,主要因為齒肋赤蘚受到嚴重的干旱脅迫,光合作用降低,部分非結構性碳水化合物轉化為保護蛋白和酶以適應對干旱的脅迫的響應。同時,可溶性總糖作為滲透調節物質抵御齒肋赤蘚受到干旱脅迫,并維持植物細胞的完整性。齒肋赤蘚在脫水過程中光合作用的恢復與光合產物的積累不是同步進行,光合產物的積累顯著遲于光合作用的恢復。這種機制的發生可能存在以下原因:(1)復水后光合作用的恢復較呼吸作用慢,復水后齒肋赤蘚恢復生理特性,需要消耗大量儲存的非結構性碳水化合物;(2)含水量在20%以上,齒肋赤蘚未受到干旱脅迫時,隨含水量的降低光合和呼吸作用都呈現下降,光合產物部分被呼吸作用消耗,部分可以轉化為結構性物質,有助于齒肋赤蘚的生長;(3)含水量下降至20%后,齒肋赤蘚受到干旱脅迫后,呼吸作用消耗部分光合產物,光合產物不再向結構性物質轉化,光合產物大量積累;(4)含水量持續下降至復水前水平,齒肋赤蘚受到嚴重的干旱脅迫,為適應脅迫,需將部分非結構性碳水化合物轉化為防止膜損傷和細胞器損傷的物質。
3.3 不同區域齒肋赤蘚非結構性碳水化合物的含量
脫水過程中兩個區域的齒肋赤蘚非結構性碳水化合物含量的變化趨勢相似。復水前、復水后和脫水過程中沙漠南緣齒肋赤蘚植株淀粉含量顯著高于沙漠腹地。齒肋赤蘚非結構性碳水化合物含量與淀粉含量相似,沙漠南緣的齒肋赤蘚非結構性碳水化合物含量顯著高于沙漠腹地。結果說明不同地區齒肋赤蘚非結構性碳水化合物含量顯著不同。沙漠腹地和沙漠南緣其土壤含水量的研究發現土壤表層含水量較低[55-57],齒肋赤蘚主要在土壤表面,沒有維管組織和根,因此齒肋赤蘚的生長主要受到降雨的影響。沙漠腹地降水的年、月、季的分布與沙漠周邊地區基本一致,10年的平均降水量123.4 mm,比沙漠周邊同期平均降水量偏少32.3%[58]。南緣的非結構性碳水化合物含量顯著高于沙漠腹地,可能由于降雨量在沙漠南緣高于沙漠腹地,齒肋赤蘚具有較好的生長,同時具有較高的非結構性碳水化合物來含量抵御高頻次的小降雨事件。復水前沙漠南緣可溶性總糖、蔗糖和果糖含量高于沙漠腹地,經過復水后的脫水過程沙漠南緣齒肋赤蘚可溶性總糖、蔗糖和果糖含量低于沙漠腹地。說明兩個地區的齒肋赤蘚可溶性糖對相同復水條件的脫水過程響應不同。沙漠腹地降雨量低,對干旱脅迫具有較強的適應能力,因此在脫水結束后中沙漠腹地的可溶性糖含量與復水前無差異。這與干旱脅迫后,抗旱性強的小麥比抗旱性弱的小麥能保持較高的可溶性總糖含量,有較強的光合及干物質積累能力[42,59-60]。淀粉含量在兩個地區保持相對穩定復水前和脫水后沒有顯著的變化,主要由于淀粉作為能量儲存物質,在受到干旱脅迫后,植物會積累大量的淀粉含量,為進入休眠和再恢復生長和代謝提供物質能量基礎[43]。然而非結構性碳水化合物含量的變化表現出復水前南緣齒肋赤蘚非結構性碳水化合物含量顯著高于腹地齒肋赤蘚,脫水過程結束后則表現為兩個地區齒肋赤蘚非結構性碳水化合物含量無顯著性差異,主要由于南緣齒肋赤蘚可溶性總糖含量出現明顯的下降。因此不同地區齒肋赤蘚非結構性碳水化合物的顯著差異和對脫水過程響應的顯著不同,可能是齒肋赤蘚對環境因素的長期適應的結果。
4 結論
(1)齒肋赤蘚復水后,非結構性碳水化合物在脫水過程中主要出現4個階段:1 h內非結構性碳水化合物含量顯著降低,2—16 h非結構性碳水化合物含量沒有顯著變化,16—24 h非結構碳水化合物含量顯著增加,24—48 h非結構性碳水化合物含量緩慢下降至復水前水平。
(2)古爾班通古特沙漠南緣齒肋赤蘚非結構性碳水化合物含量顯著高于沙漠腹地。古爾班通古特沙漠不同區域齒肋赤蘚非結構碳水化合物含量顯著不同。
(3)古爾班通古特沙漠腹地與南緣齒肋赤蘚的非結構性碳水化合物對脫水過程具有不同的響應,主要表現在兩個地區齒肋赤蘚可溶性糖含量變化的顯著不同。
致謝:感謝尹本豐博士和朱秉堅博士對野外采樣和室內測定非結構性碳水化合物含量實驗的幫助。
