不同生境下景寧玉蘭灌叢萌株形態及其生殖特征
2019-01-18 05:51:46杜有新吳偉建劉躍鈞王軍峰季志利章偉民何小勇
生態學報 2018年23期
杜有新,吳偉建,劉躍鈞,王軍峰,季志利,章偉民,何小勇,*
1 麗水市林業科學研究院,麗水 323000 2 松陽縣林業局,松陽 323400 3 麗水蓮都生態林場,麗水 323000
景寧玉蘭(MagnoliasinostellataP. L. Chiu et Z. H. Chen) 屬浙江特有種[1- 2],為國家二級保護的瀕危物種[3- 4],具有較高的科學研究和園林觀賞價值[5]。景寧玉蘭集中分布于浙江南部的麗水市景寧畬族自治縣草魚塘林場、松陽縣牛頭山地區、蓮都區峰源鄉和溫州市樂清雁蕩山,其原生環境相似,由于人類活動的干擾及自然環境變化導致的生境破碎化,現代分布格局呈分散性特征[6],主要生長于海拔900m以上的落葉闊葉林、杉木林、黃山松林、林緣溝邊和灌叢中[7]。
景寧玉蘭為淺根性落葉灌木,喜陰涼濕潤、水分充足且土壤肥沃的生境,對環境變化較為敏感[7- 8]。在自然環境下,景寧玉蘭自然結實率不到1%[9],但其萌生性強,莖稈基部具有很強的萌芽能力[2],實生苗定植后往往萌發數根萌蘗[10],根頸萌枝[11]和殘樁萌枝[12]現象在原生環境下普遍存在,這也是其對資源狀況及環境變化的一種適應性響應[13-15]。之前對景寧玉蘭生態生物學特性[2]、種群分布[6- 7]、花粉活力[8- 9]、無性繁殖[10]及屬內親緣關系[16]等方面開展了較多研究,但未見有關不同生境下景寧玉蘭萌枝形態及生殖特征方面的報道。萌生是植物抵抗不利環境的一種營養繁殖方式一,是面臨較大強度干擾及實生更新受到限制情況下的一種重要自然更新方式[17],研究萌枝形態及其生殖特征可以深入了解植物在不利環境下的生長發育狀況及其應對策略。本文通過調查分析不同生境下景寧玉蘭萌枝形態及開花結實狀況,以期了解景寧玉蘭自然更新特征及其與生境的關系,為科學保護合理利用其種質資源提供參考。
1 研究地概況
本研究選取景寧玉蘭代表性的3個分布點即麗水市松陽縣大東壩鎮燈塔村牛頭山、麗水市蓮都區峰源鄉尤源村和葑垟村,3個分布點相距20—60km。分布點均處于山體北坡,含蓋了景寧玉蘭自然生長的各種生境類型,除了松陽牛頭山海拔720m處生長著3叢外,景寧玉蘭主要分布于海拔950—1300m之間。景寧玉蘭在浙南山區的分布區位于27°30′—28°26′N,119°49′—120°05′E之間的北坡[6],土壤為山地黃壤和黃棕壤,土層厚度30—120cm,pH值4.8—5.5[2]。其中在峰源鄉的自然分布區位于28°05′—28°14′N,119°40′—119°51′E之間,年均氣溫處于12.0—15.5℃,海拔950—1100m,年均相對濕度87%[18]。牛頭山分布區處于28°15′—28°17′N,119°32′—119°35′E之間,年均氣溫處于11.5—16.5℃,海拔700—1285m,年均相對濕度87%。研究樣地的地理信息及植被概況見表1。
2 研究方法
2.1 樣地設置
于2016年2月,根據景寧玉蘭自然生長的生境條件,把景寧玉蘭生境分為5種類型,分別為灌叢(SH)、黃山松(Pinustaiwanensis)林(PTF)、落葉闊葉林(DBF)、杉木(Cunninghamialanceolata)林(CLF)及林緣溝邊(FE),其中林緣是指樹林邊緣5m內區域,距離樹林邊緣10m視為林內。根據景寧玉蘭分布狀況,在每種生境上分別設置1—3個典型樣地,樣地面積10m × 20m,每個樣地劃分為8個5m × 5m的小樣方,尤源村共設置了4個樣地;葑垟村灌叢生境設置3個10m × 20m樣地,而林緣生境的景寧玉蘭居群主要分布于長為100余m的毛竹林和杉木林的林緣溝邊,設置2個面積5m × 40m樣地,每個樣地劃分為8個5m × 5m的小樣方(表1)。在松陽縣牛頭山地區,景寧玉蘭分布于海拔716—1300m之間,結合景寧玉蘭種群數量,在海拔720、1100m和1250m處分別設置1、2和3個樣地。海拔720m處只發現相距不到5m的3叢,直到海拔1030m后才再次出現景寧玉蘭植株,另外景寧玉蘭在海拔1000—1200m的落葉闊葉林中只零散分布著9叢,因此,在720m處只設置了1個樣地,在落葉闊葉林生境設置2個樣地,牛頭山上共設置6個樣地(表1)。

表1 景寧玉蘭群落樣地概況(均值±標準差)
SH,灌叢,Shrubberies; PTF,臺灣松林,Pinustaiwanensisforest;DBF,落葉闊葉林,Deciduous broad-leaved forest;CLF,杉木林,Cunninghamialanceolataforest;FE,林緣,Forest edges
2.2 調查方法
由于灌叢年齡結構的確定受到方法上的限制,通常采用立木的大小徑級代替不同的年齡結構,但至今也沒有景寧玉蘭徑級與樹齡關系的資料。景寧玉蘭屬世代重疊及生殖連續的多年生植物,筆者根據實際調查和相關文獻[19],以景寧玉蘭每叢最大萌枝作為母株,按其基徑(BD)大小劃分為5個年齡級,分別為I:BD<1.5cm,II:1.5≤BD<3cm,III:3≤BD<4.5cm,IV:4.5≤BD<6cm,V級:BD≥6cm。本研究樣地上的景寧玉蘭均為天然更新苗,根據上述標準,所有樣地景寧玉蘭種群年齡均處于II至IV之間。
調查統計樣地內景寧玉蘭株(叢)數及其冠幅,所有從母株根頸部或其地表根出萌枝均視為其萌枝,觀測每叢所有萌枝的基徑和高度等信息,記錄伴生植物種類,同時記錄樣地的經緯度、海拔、坡向、坡度等生態因子及干擾情況。景寧玉蘭原生地存在從母株根部萌生的根出條萌枝和機會萌枝[11]現象,在實際調查中把與母株相連的萌枝均視為母株萌枝,不計為獨立的植株,野外調查中沒發現萌生起源的植株。在每個樣地內選取2—3叢代表性成年(III級)植株,在其冠層上部標記3根主枝,用直尺和游標卡尺測量各級枝的基徑和枝長。按Strahler法確定分枝序列[20],冠層最外層的第一小枝為第一級,兩個以上第一級相遇即為第二級,兩個以上第二級相遇后則為第三級,依此類推。光照強度采用浙江托普儀器公司的農業環境檢測儀(型號DJL- 18)測定。
于2016年2—3月,對每個樣地景寧玉蘭植株及每叢萌枝全部編號掛牌,于2016年和2017年的2—3月份,每叢開花的萌枝計為生殖萌枝,花期調查每根生殖萌枝花量,統計每叢花量及每個樣地開花叢數。于2016年和2017年的8—9月,在景寧玉蘭果實成熟期,對每個樣地景寧玉蘭植株,根據花期調查時每個樣地上的編號植株,逐叢逐根萌枝進行結實情況調查,對采集的果實進行編號,用直尺和游標卡尺測量每個果實的長度和直徑,果實開裂后記錄每個果實種子粒數,統計生殖萌枝結實率和每叢結實率。
2.3 數據處理與分析
萌枝總分枝率(Overall bifurcation ratio,OBR)和逐級分枝率(Stepwise bifurcation ratio,SBR)分別采用如下公式[20]計算:總分枝率OBR=(NT-Ns)/(NT-N1),逐級分支率SBRi:(i+1)=Ni/N(i+1),式中NT為所有枝級中的枝條總和,NS為最高級枝條數,N1為第一級的枝條數,Ni和Ni+1分別為第i級和第i+1級的枝條數。生殖萌枝花量為每根生殖萌枝著花的數量,每叢生殖萌枝比例=每叢開花的萌枝數/每叢萌枝總數;生殖萌枝結實率=每根生殖萌枝果實數量/每根生殖萌枝花量總數,每叢結實率=每叢果實總數/每叢花量總數。
把松陽縣牛頭山的景寧玉蘭居群劃分為3個海拔梯度,即海拔700—1000m、1000—1200m和>1200m。分析海拔高度對景寧玉蘭形態及生殖性狀的影響。選取海拔950—1200m處的灌叢、落葉闊葉林、杉木林和林緣等4種生境,分析不同生境類型下景寧玉蘭形態及生殖性狀差異。通過單因素方差分析(ANOVA)和多重比較(LSD)檢驗不同植被類型和不同海拔高度間景寧玉蘭萌枝形態及生殖性狀的顯著性差異,采用Pearson相關性分析檢驗各性狀之間的相關性特征。所有數據均在Excel 2003和SPSS 17.0軟件上進行。
3 結果與分析
3.1 不同生境景寧玉蘭萌枝形態特征
景寧玉蘭萌枝形態特征及分枝率在不同生境之間存在一定差異(表2)。黃山松林下的景寧玉蘭密度最高,每公頃平均達732叢,顯著高于落葉闊葉林和杉木林(P<0.05),而且落葉闊葉林下景寧玉蘭萌枝基徑顯著小于其他林分(P<0.05)。在分枝特征方面,黃山松和落葉闊葉林下景寧玉蘭一級枝基徑顯著小于其他3種生境(P<0.05),黃山松林下景寧玉蘭一級枝長度即年生長量顯著低于其他4種生境(P<0.05)。灌叢中景寧玉蘭的總分枝率顯著高于杉木林落葉闊葉林和黃山松林(P<0.05),灌叢和林緣生境上的景寧玉蘭逐步分枝率均處于較高水平,也許光照條件不受限制,高分枝率有利于拓展更大的生存空間。
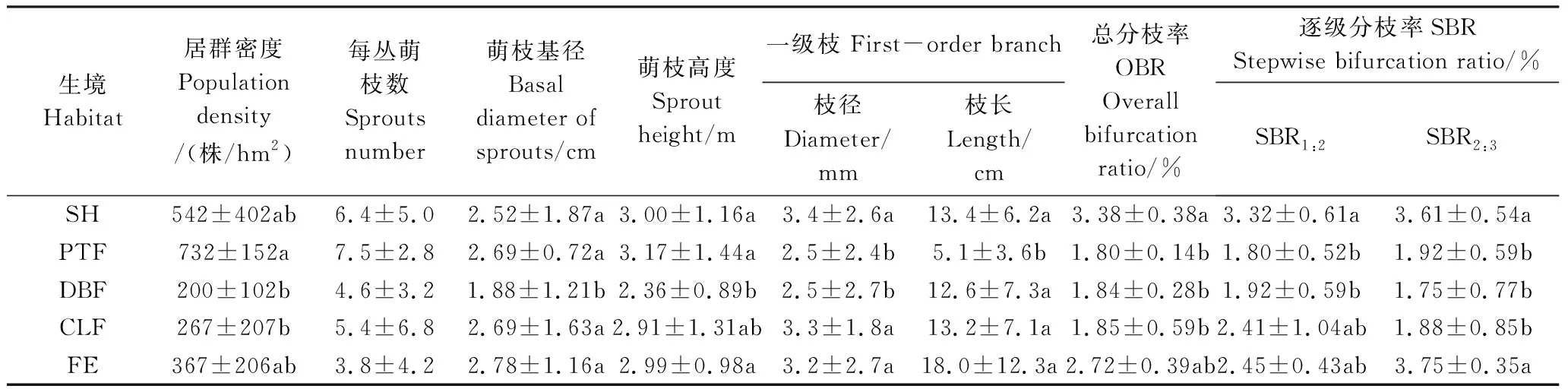
表2 不同生境下景寧玉蘭形態特征 (均值±標準差)
同列中不同小寫字母者表示差異顯著水平(P< 0.05)
3.2 不同生境景寧玉蘭生殖特征
不同生境下景寧玉蘭平均生殖萌枝比例及生殖萌枝開花數量存在差異(表3)。其中黃山松林、落葉闊葉林和林緣的景寧玉蘭生殖萌枝花量顯著低于杉木林(P<0.05),灌叢與其他生境沒有差異。黃山松林下景寧玉蘭生殖萌枝比例顯著低于灌叢、杉木林和林緣(P<0.05),落葉闊葉林下景寧玉蘭生殖萌枝比例顯著低于林緣。景寧玉蘭萌枝生殖性狀除落葉闊葉林外,其他4種生境上的景寧玉蘭生殖萌枝結實率沒有差異。黃山松林下的景寧玉蘭果實大小、單果種子數及每叢結實率最高(P<0.05),落葉闊葉林下景寧玉蘭沒有結實植株,其他3種生境沒有差異。景寧玉蘭生殖特征也許與其受到的光照資源及花期氣溫密切相關,有待深入研究。

表3 不同生境下景寧玉蘭生殖特征(均值±標準差)
3.3 萌枝性狀相關性分析
通過萌枝基徑與高度之間及其與萌枝花量的相關分析表明,萌枝基徑與高度具有極顯著正相關關系(P<0.01),萌枝基徑(P<0.01)和高度(P<0.01)與萌枝花量均有極顯著正相關關系(圖1)。每叢最大萌枝基徑與高度之間具極顯著正相關關系(P<0.01),最大萌枝基徑與每叢萌枝數具有顯著正相關關系(P<0.05),而最大萌枝高度與每叢萌枝數沒有相關關系(圖1)。通過生殖萌枝和每叢花量及其與結實率之間相關分析表明,生殖萌枝花量與萌枝結實率沒有關系(P>0.05),每叢花量與每叢結實率之間也沒有關系(P>0.05)。結果說明萌枝大小與其高度密切相關,并影響著景寧玉蘭的開花數量,但每叢開花數量與結實狀況沒有顯著相關關系。

圖1 萌枝形態和生殖性狀的相關性Fig.1 Relationship between morphological and reproductive characters of sprouts
3.4 萌枝性狀的變異性分析
生境類型和海拔高度顯著影響著景寧玉蘭萌枝的基徑、高度和一級枝長度(P<0.05),海拔對一級枝粗度和長度具有顯著影響(P<0.05),但生境類型和海拔高度對每叢萌枝數及分枝率均沒有影響(表4)。生境類型和海拔高度對生殖萌枝花量、生殖萌枝比率及生殖萌枝結實率具有顯著影響(P<0.05)。說明景寧玉蘭萌枝形態及其生殖生物學性狀與植株所處的生境條件密切相關。

表4 生境類型和海拔對景寧玉蘭萌枝形態及生殖萌枝特征的影響
*和**分別表示顯著性差異(P<0.05)和極顯著性差異(P<0.01)
4 討論
植物的萌生能力受內在生物學特性及微生境、資源水平和干擾狀況等外部因子的綜合影響[21]。景寧玉蘭屬萌生能力較強的密集型生長類型[10],以根頸萌生類型為主[2, 14]。萌生能力與母株的大小存在一定的相關關系,但萌枝數量與母株高度沒有相關關系(圖1 E—1F),這與萌生能力很強的亞熱帶喬木樹種米心水青岡(Fagusengleriana)有點不同,米心水青岡的萌生能力與母株的大小以及萌枝數量與母株高度均呈正相關關系[22]。與溫帶落葉樹種連香樹(Cercidiphyllumjaponicum)也不同,連香樹萌枝數與立地條件和根系暴露程度有關[23],而景寧玉蘭植株萌枝數量在不同生境之間也沒有差異,如在人類活動較為頻繁的尤源村灌叢和沒有干擾的牛頭山黃山松林中,景寧玉蘭每叢的萌枝數量沒有差異,但不同生境之間的萌枝大小及高度存在差異(表2),說明景寧玉蘭的萌枝能力很可能主要由其生物學特性控制[11],而萌枝大小及高度差異可能是萌枝間以及個體間光照及土壤養分資源的競爭結果[14,24],有待深入研究。
萌枝形態特征能較好地反映植物對資源與環境變化的適應狀況[20]。雖然適度低溫和陰濕環境是玉蘭亞屬較為理想的生存環境[25],但景寧玉蘭在不同生境上,因為所處的微生境和面臨的限制因素各不相同,分枝構型在不同生境上的表現有較大差異。對林下植物而言,有機物質積累與其接受的光資源密切相關,光資源條件是影響植物萌蘗和增粗的主要因素[15],如黃山松和闊葉林下生境,由于上層喬木樹種遮蔽,具有林下光照較少、種內種間競爭較為激烈,景寧玉蘭可能通過低分枝率(表2)以增加有效光合作用面積。海拔通過環境條件的變化間接影響著植物生長[26],中低海拔的闊葉林和杉木林下的景寧玉蘭一級枝粗度和長度顯著高于黃山松林下,可能與土壤有機質和堿解氮含量有關[27]。植物群落及土壤環境對景寧玉蘭生長及枝型特征均有影響,雖然不同生境上植被群落性質不同[6]和缺乏完整的土壤環境數據,但由于林緣生境的灌木覆蓋度較低和灌從生境上沒有上層林木遮蔽(表1),景寧玉蘭可獲得較充分的光照和空間資源[20],有利于其一級枝生長和提高逐步分枝率,增強其與伴生樹種的生存競爭力 。
研究結果表明,景寧玉蘭生殖萌枝的花量均與萌枝大小和高度呈正相關關系(圖1),但生殖萌枝比例及其花量與每叢最大萌枝的高度沒有關系(P>0.05),說明植物開花是以植物體內有機物質的積累為基礎,但個體生殖成功與否還受到花期環境條件及傳粉昆蟲活動狀況的綜合影響[24,28]。本研究發現,景寧玉蘭植株的生殖萌枝花量及其比率與海拔高度有關(表4),隨海拔升高而逐漸下降,如從低海拔的杉木林、落葉闊葉林到高海拔的黃山松林,景寧玉蘭植株的生殖萌枝花量及其比率呈逐漸下降的趨勢;但其結實狀況卻呈現與開花性狀不同的現象,如高海拔黃山松林下景寧玉蘭生殖萌枝果實數、單果種子數和每叢平均結實率均明顯高于中海拔的落葉闊葉林和杉木林(表3),可能黃山松林下景寧玉蘭早春開花時,伴生樹種金縷梅同期開花,在吸引傳粉昆蟲方面發揮了積極作用,說明環境條件是影響景寧玉蘭生殖成功與否的重要因素[8,29]。不同植被類型對景寧玉蘭的生殖萌枝開花及結實情況也有一定影響(表4),如景寧玉蘭在杉木林下的生殖萌枝花量顯著高于林緣,林緣生殖枝比例顯著高于落葉闊葉林,落葉闊葉林下景寧玉蘭沒有發現結實現象(表3)。景寧玉蘭屬先花后葉植物,雖然個體光合有機產物的積累是影響個體生殖構件發育的直接因素,但花果期敗育狀況與環境條件關系密切[24],有機物質分配策略與雌雄花異熟的生物學特性[29-30]和傳粉昆蟲多寡及其訪花效率均是影響生殖成功的主要因素[31]。
植物體是作為一個整體對外部環境變化做出反應,由于環境變化導致的某個性狀變化可能引起其他性狀的相應變化,進而影響著植物體對變化環境的整體響應。本研究只是分析比較了不同生境上景寧玉蘭萌枝的形態及生殖生物學性狀,今后應重點關注微生境變化及傳粉昆蟲活動對景寧玉蘭生態生理及生殖發育方面的影響,探討影響景寧玉蘭生殖成功率的關鍵因素及其影響機制,深入了解景寧玉蘭的生態適應性和種群健康維持機制。
5 結論
(1)不同生境下景寧玉蘭萌枝形態特征存在差異。景寧玉蘭萌生能力與母株的大小存在一定關系,但萌枝大小和高度與生境更為密切相關。其枝系特征在不同生境上的表現也有較大差異,落葉闊葉林下景寧玉蘭萌枝大小及分枝率最低。
(2)不同生境下景寧玉蘭萌枝生殖特征存在差異。景寧玉蘭生殖萌枝的花量均與萌枝大小及其高度呈正相關關系,生殖萌枝的花量與其結實率沒有關系。落葉闊葉林下景寧玉蘭生殖枝花量及其結實率均最低,雖然黃山松林下景寧玉蘭生殖枝花量較低,但其萌枝結實率、單果種子數及每叢結實率最高。
(3)生境條件對景寧玉蘭萌枝形態及其生殖特征存在顯著影響。生境類型和海拔高度對景寧玉蘭萌枝基徑、高度、一級枝長度、生殖萌枝花量、生殖萌枝比率及生殖萌枝結實率均存在顯著影響。
致謝:松陽縣林業局景寧玉蘭小區護林員雷國興和麗水蓮都生態林場江麗榮參加部分野外工作,特此致謝。
