基于轉(zhuǎn)錄組信息的黑果枸杞WD40蛋白質(zhì)家族分析
2019-02-21 11:12:50陳建偉王翠平喬改霞
核農(nóng)學(xué)報(bào) 2019年3期
嚴(yán) 莉 陳建偉 王翠平,? 仝 倩 王 晨 喬改霞 李 健
(1寧夏大學(xué)生命科學(xué)學(xué)院,寧夏 銀川 750021;2寧夏林業(yè)研究院種苗生物工程國(guó)家重點(diǎn)實(shí)驗(yàn)室,寧夏 銀川 750004)
WD40 蛋白質(zhì)又稱WD40 重復(fù)體,廣泛存在于真核生物中,在原核生物中極少見[1]。WD40 蛋白質(zhì)中的WD40 基序高度保守,通常約含40 ~60 個(gè)氨基酸殘基,在其N 末端含有甘氨酸-組氨酸(GH)二肽,C 末端含有色氨酸-天冬氨酸(WD)二肽,中間被可變區(qū)域隔開[2-5]。WD40 基序可作為蛋白質(zhì)-蛋白質(zhì)及蛋白質(zhì)-核酸相互作用的支架[6],多數(shù)WD40 蛋白質(zhì)包含至少由4 個(gè)WD40 基序折疊形成的β 螺旋槳結(jié)構(gòu),并通過該結(jié)構(gòu)發(fā)揮作用[7]。
研究表明,WD40 蛋白質(zhì)廣泛存在于細(xì)胞質(zhì)和核質(zhì)中,參與多種重要的細(xì)胞過程[8],包括信號(hào)轉(zhuǎn)導(dǎo)[9]、基因轉(zhuǎn)錄調(diào)控[10]、細(xì)胞骨架動(dòng)力學(xué)[11]、膜泡運(yùn)輸[12]、DNA 損傷及修復(fù)[13]、細(xì)胞死亡和細(xì)胞周期調(diào)控[14]等。目前對(duì)植物WD40 蛋白質(zhì)的研究主要集中于模式植物擬南芥,如Lee 等[15]通過酵母雙雜交實(shí)驗(yàn)分離得到擬南芥WD40 蛋白質(zhì)AtSeh1,分析其結(jié)構(gòu)發(fā)現(xiàn)AtSeh1 含有4 個(gè)WD40 基序,為COPⅡ囊泡的重要組成部分,可通過控制動(dòng)力蛋白AtDRP2A(控制從反面高爾基網(wǎng)絡(luò)到中央液泡的蛋白運(yùn)輸)來調(diào)控膜泡運(yùn)輸;Gachomo等[16]通過反向遺傳和蛋白質(zhì)建模分離出擬南芥WD40 蛋白質(zhì)家族成員GIGANTUS1(GTS1),并發(fā)現(xiàn)其可通過調(diào)控細(xì)胞中核糖體的結(jié)構(gòu)、活動(dòng)和生物遺傳來調(diào)節(jié)植物的生長(zhǎng)發(fā)育;Cheng 等[17]研究發(fā)現(xiàn)擬南芥GSK3 樣激酶可直接調(diào)控MYB-bHLH-WD40 轉(zhuǎn)錄因子復(fù)合體的表達(dá)來調(diào)控植物體內(nèi)油菜素內(nèi)酯的含量進(jìn)而調(diào)控植物根毛區(qū)細(xì)胞的發(fā)育;Beris 等[18]通過RNA 干擾介導(dǎo)基因沉默表達(dá)的方法分離出擬南芥WD40 蛋白質(zhì)AtULCS1,發(fā)現(xiàn)其通過與植物泛素環(huán)連接酶亞基DDB1α 相互作用導(dǎo)致某種特異酶的降解,來抑制花藥細(xì)胞次生壁的形成。
諸多研究學(xué)者對(duì)其他生物WD40 蛋白質(zhì)也進(jìn)行了相關(guān)研究,如Ge 等[19]發(fā)現(xiàn)煙草NtTTG1 和NtTTG2 可分別與誘導(dǎo)蛋白ParA1、參與生長(zhǎng)素信號(hào)轉(zhuǎn)導(dǎo)途徑的3種蛋白質(zhì)(NtARF8、NtARF17 和NtARF19)共同作用來調(diào)控?zé)煵莸纳L(zhǎng)發(fā)育及對(duì)抵御病原菌的浸入,說明WD40 蛋白質(zhì)可與多種蛋白質(zhì)相互作用促進(jìn)植物的生長(zhǎng)發(fā)育;Lee 等[20]分離出一種歐洲油菜WD40 蛋白質(zhì)BnSWD1,發(fā)現(xiàn)其在鹽脅迫、脫落酸、茉莉酸甲酯及水楊酸處理下的表達(dá)水平較對(duì)照顯著升高,推測(cè)該基因可能依賴脫落酸或其他信號(hào)來響應(yīng)鹽脅迫,為研究WD40 蛋白質(zhì)響應(yīng)非生物脅迫提供一種新思路;Guerriero 等[21]在子囊菌中分離出WD40 蛋白質(zhì)編碼基因WDR,分析后發(fā)現(xiàn)其是真菌幾丁質(zhì)生物合成基因簇中的保守成員,并參與了細(xì)胞壁的形成調(diào)控,Beris等[18]在擬南芥中也得出類似結(jié)果,說明在植物和真菌中WD40 蛋白質(zhì)功能具有保守性。
黑果枸杞(Lycium ruthenicum Murr.)為西北干旱半干旱地區(qū)特色灌木樹種,其果實(shí)中花青素含量豐富,經(jīng)濟(jì)價(jià)值較高[22-23]。目前關(guān)于黑果枸杞的研究報(bào)道主要集中于對(duì)其活性成分、代謝途徑、耐鹽性及遺傳轉(zhuǎn)化研究4 個(gè)方面,對(duì)黑果枸杞類黃酮及花青素合成途徑關(guān)鍵基因的研究尚不常見。本研究通過對(duì)黑果枸杞青色期、轉(zhuǎn)色期和成熟期3 個(gè)發(fā)育時(shí)期的果實(shí)進(jìn)行轉(zhuǎn)錄組測(cè)序,根據(jù)測(cè)序結(jié)果篩選黑果枸杞WD40 蛋白質(zhì)家族成員,對(duì)其進(jìn)行結(jié)構(gòu)域組成、亞細(xì)胞定位、進(jìn)化關(guān)系分析,并對(duì)WD40 蛋白質(zhì)編碼基因成員在果實(shí)生長(zhǎng)發(fā)育3 個(gè)時(shí)期的表達(dá)變化進(jìn)行分析,以期為黑果枸杞中WD40 蛋白質(zhì)的功能研究奠定一定的基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料及處理
2016年7月12日在寧夏林業(yè)研究院枸杞資源試驗(yàn)田選取生長(zhǎng)健康、長(zhǎng)勢(shì)良好的黑果枸杞植株(組培苗扦插繁育)并標(biāo)記,開花后25 d 開始采摘青果期果實(shí),40 d 后采摘轉(zhuǎn)色期果實(shí),58 d 后采摘成熟期果實(shí)(各生育期取樣果實(shí)大小、飽滿度一致)。以3 株黑果枸杞的果實(shí)為一個(gè)重復(fù),設(shè)3 次生物學(xué)重復(fù),取樣后立即用液氮速凍,-80℃保存?zhèn)溆谩?/p>
1.2 試驗(yàn)方法
1.2.1 果實(shí)總RNA 提取、文庫(kù)構(gòu)建及轉(zhuǎn)錄組測(cè)序 利用天根生化科技(北京)有限公司提供的植物總RNA 提取試劑盒(RNA Prep Pure Plant Kit)分別提取黑果枸杞青色期(G1 ~G3)、轉(zhuǎn)色期(C1 ~C3)和成熟期(B1~B3)果實(shí)的總RNA,采用NanoDrop-2000 核酸檢測(cè)儀(上海Thermo Scientific 有限公司)檢測(cè)RNA 純度(OD260/OD280),通過Qubit? 1.0 熒光儀(上海恒斐生物科技有限公司)對(duì)RNA 濃度進(jìn)行精確定量,然后用Agilent 2100 生物分析儀(安捷倫,美國(guó))精確檢測(cè)RNA 的完整性,最后以1%的瓊脂糖凝膠電泳。樣品檢測(cè)合格后送至北京諾禾致源科技股份有限公司進(jìn)行轉(zhuǎn)錄組測(cè)序,后續(xù)分析基于此測(cè)序結(jié)果數(shù)據(jù)。
1.2.2 黑果枸杞和擬南芥WD40 蛋白質(zhì)編碼基因的篩選 基于轉(zhuǎn)錄組測(cè)序數(shù)據(jù),在Nr、Nt、Pfam、KOG/COG、Swiss-prot、KEGG 及GO 7 個(gè)數(shù)據(jù)庫(kù)進(jìn)行注釋,初步注釋到136 個(gè)WD40 基因,將這些基因編碼的蛋白質(zhì)逐個(gè)進(jìn)行NCBI Blastp 和SMART 預(yù)測(cè),去除重復(fù)序列或短序列后,最終得到77 個(gè)WD40 基因。擬南芥WD40 蛋白質(zhì)序列從TAIR(http:/ /www.arabidopsis.org/)數(shù)據(jù)庫(kù)中下載獲得。
1.2.3 黑果枸杞WD40 蛋白質(zhì)家族結(jié)構(gòu)域、亞細(xì)胞定位及進(jìn)化關(guān)系分析 對(duì)初步注釋得到的136 個(gè)黑果枸杞WD40 蛋白質(zhì)逐個(gè)進(jìn)行NCBI Blastp 和SMART 預(yù)測(cè),得到每個(gè)WD40 蛋白質(zhì)的結(jié)構(gòu)域組成,根據(jù)其所含結(jié)構(gòu)域分析其結(jié)構(gòu)特征;利用在線軟件Prot Comp 9.0(http:/ /linux.softberry.com)對(duì)77 個(gè)候選WD40 基因家族各成員的氨基酸序列進(jìn)行亞細(xì)胞定位預(yù)測(cè)分析;利用MEGA6.0 軟件對(duì)77 個(gè)黑果枸杞WD40 蛋白質(zhì)和部分功能明確的擬南芥WD40 蛋白質(zhì)(共34 個(gè),包括僅含WD40 基序的WD40 蛋白質(zhì)13 個(gè),同時(shí)包含WD40 基序和其他結(jié)構(gòu)域的WD40 蛋白質(zhì)21 個(gè))進(jìn)行序列比對(duì),并用鄰接(Neighbor-Joining)法構(gòu)建系統(tǒng)進(jìn)化樹,進(jìn)行Bootstrap 測(cè)試,重復(fù)設(shè)置為1 000,其他參數(shù)為默認(rèn)設(shè)置。
1.2.4 黑果枸杞WD40 基因的表達(dá)模式分析 黑果枸杞果實(shí)轉(zhuǎn)錄組測(cè)序文庫(kù)中各基因片段表達(dá)量的計(jì)算采用RPKM(reads per kb per million) 法[24]。采 用Heml 軟件對(duì)77 個(gè)黑果枸杞WD40 基因在果實(shí)發(fā)育各生育期的表達(dá)數(shù)據(jù)進(jìn)行分層聚類分析,得到黑果枸杞WD40 基因家族成員隨果實(shí)發(fā)育的表達(dá)模式。
2 結(jié)果與分析
2.1 黑果枸杞果實(shí)總RNA 提取、文庫(kù)構(gòu)建及轉(zhuǎn)錄組測(cè)序結(jié)果分析
由圖1 可知,9 種RNA 樣品(G1、G2、G3、C1、C2、C3、B1、B2 和B3)的28S 和18S 條帶清晰明亮,且28S和18S 條帶的亮度比接近2 ∶1,各條帶均沒有拖尾現(xiàn)象,各RNA 樣品無(wú)DNA 和蛋白污染,說明9 個(gè)樣品RNA 的質(zhì)量和完整性較好,可用于后續(xù)研究。
轉(zhuǎn)錄組測(cè)序結(jié)果表明,共得到194 385 條基因序列,其中有138 322 個(gè)基因被注釋到Nr、Nt、Pfam、KOG/COG、Swiss-prot、KEGG 和GO 功能數(shù)據(jù)庫(kù)中,占基因總數(shù)的71.15%,被同時(shí)注釋到7 大數(shù)據(jù)庫(kù)的基因序列有18 088 條。基因進(jìn)行KO 注釋后對(duì)參與的KEGG 代謝通路進(jìn)行分類分析,發(fā)現(xiàn)這些基因參與的KEGG 代謝通路共分為5 個(gè)分支,即細(xì)胞過程(cellular process)、 環(huán) 境 信 息 處 理(environmental information processing)、 遺 傳 信 息 處 理( genetic information processing)、代謝(metabolism)和有機(jī)系統(tǒng)(organismal system)。
2.2 黑果枸杞WD40 蛋白質(zhì)編碼基因的篩選與蛋白質(zhì)序列的分類分析
對(duì)所得77 條黑果枸杞WD40 蛋白質(zhì)序列進(jìn)行批量SMART 預(yù)測(cè),根據(jù)其所包含的結(jié)構(gòu)域類型將其分為僅含WD40 基序的WD40 蛋白質(zhì)和同時(shí)含有其他結(jié)構(gòu)域的WD40 蛋白質(zhì),最終鑒定出僅含WD40 基序的黑果枸杞WD40 蛋白質(zhì)有38 個(gè),包括含1 個(gè)WD40 基序的蛋白質(zhì)1 個(gè),含2 個(gè)WD40 基序的蛋白質(zhì)有3 個(gè),含3 個(gè)WD40 基序的蛋白質(zhì)2 個(gè),含4 個(gè)WD40 基序的蛋白質(zhì)6 個(gè),含5 個(gè)WD40 基序的蛋白質(zhì)3 個(gè),含6個(gè)WD40 基序的蛋白質(zhì)7 個(gè),含7 個(gè)WD40 基序的蛋白質(zhì)11 個(gè),含8 個(gè)WD40 基序的蛋白質(zhì)2 個(gè),含11 個(gè)WD40 基序的蛋白質(zhì)2 個(gè),含13 個(gè)WD40 基序的蛋白質(zhì)1 個(gè)。鑒定出同時(shí)含有WD40 基序和其他結(jié)構(gòu)域的WD40 蛋白質(zhì)共39 個(gè),這些蛋白質(zhì)共包含25 種結(jié)構(gòu)域類型(圖2)。
2.3 黑果枸杞WD40 蛋白質(zhì)的亞細(xì)胞定位分析
由圖3 可知,共有42 個(gè)WD40 蛋白質(zhì)定位于細(xì)胞質(zhì)中,占鑒定出的WD40 蛋白質(zhì)總數(shù)的54.55%;24 個(gè)蛋白質(zhì)定位于細(xì)胞核中,占鑒定出的WD40 蛋白質(zhì)總數(shù)的31.17%;2 個(gè)蛋白質(zhì)定位于細(xì)胞膜上,1 個(gè)蛋白質(zhì)定位于細(xì)胞壁上,3 個(gè)蛋白質(zhì)定位于線粒體上,5 個(gè)蛋白質(zhì)定位于內(nèi)質(zhì)網(wǎng)上。其中,定位于細(xì)胞核及細(xì)胞質(zhì)上的WD40 蛋白質(zhì)占鑒定出的總WD40 蛋白質(zhì)的85.71%,說明WD40 蛋白質(zhì)主要位于細(xì)胞核及細(xì)胞質(zhì)中。此外,還有11 個(gè)WD40 蛋白質(zhì)分別定位于細(xì)胞膜、細(xì)胞壁、內(nèi)質(zhì)網(wǎng)及線粒體上,說明WD40 蛋白質(zhì)在黑果枸杞中分布范圍廣,有利于其行使多種功能。
2.4 黑果枸杞與擬南芥WD40 蛋白質(zhì)家族的比較分析
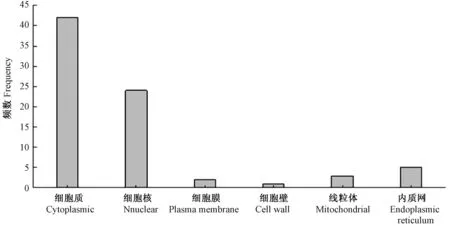
圖3 黑果枸杞WD40 蛋白質(zhì)家族的亞細(xì)胞定位預(yù)測(cè)Fig.3 Sub-cellular localization of WD40 protein family in L.ruthenicum Murr.
利用MEGA5.0 軟件構(gòu)建黑果枸杞和擬南芥WD40 蛋白家族的系統(tǒng)進(jìn)化樹,根據(jù)進(jìn)化樹(圖4)的拓?fù)浣Y(jié)構(gòu)對(duì)兩個(gè)物種的WD40 蛋白進(jìn)行聚類分析,共分為五個(gè)亞家族,第Ⅰ個(gè)亞家族包含17 個(gè)WD40 蛋白質(zhì),其中黑果枸杞WD40 蛋白質(zhì)Cluster-26021.51455和擬南芥WD40 蛋白質(zhì)AT2G19430.1 處于同一進(jìn)化亞枝;第Ⅱ亞家族包含18 個(gè)WD40 蛋白質(zhì),其中黑果枸杞WD40 蛋白質(zhì)Cluster-26021.93652 與擬南芥WD40 蛋白質(zhì)AT1G65580.1 處于同一進(jìn)化亞枝上;第Ⅲ亞家族包含14 個(gè)WD40 蛋白質(zhì),其中黑果枸杞WD40 蛋 白 質(zhì) Cluster - 26021.10572、 Cluster -26021.24543 和Cluster-26021.71813 分別與擬南芥WD40 蛋白質(zhì)AT1G61720.1 和AT5G24520.1 位于同一進(jìn)化亞枝;第Ⅳ亞家族包含26 個(gè)WD40 蛋白質(zhì),其中黑果枸杞WD40 蛋白質(zhì)Cluster-26021.77781 與擬南芥WD40 蛋白質(zhì)AT2G41500.1 位于同一進(jìn)化亞枝;第Ⅴ亞家族包含38 個(gè)WD40 蛋白質(zhì),為進(jìn)化樹中WD40 成員最多的亞家族,其中黑果枸杞WD40 蛋白質(zhì)Cluster-26021.46098 和Cluster-26021.86872 均與擬南芥AT4G09820.1 位于同一進(jìn)化亞枝。根據(jù)擬南芥數(shù) 據(jù) 庫(kù)(TAIR: http:/ /www.arabidopsis.org/) 中WD40 蛋白質(zhì)的功能注釋,擬南芥WD40 蛋白質(zhì)AT1G61720.1、AT5G24520.1 和AT4G09820.1 均參與了類黃酮及花青素的代謝調(diào)控,而黑果枸杞WD40 蛋白質(zhì)Cluster - 26021.105724、Cluster - 26021.24543、Cluster-26021.46098 和Cluster-26021.86872 與其進(jìn)化關(guān)系較近,可能參與了黑果枸杞中花青素的代謝調(diào)控。
2.5 黑果枸杞WD40 蛋白質(zhì)編碼基因在果實(shí)不同發(fā)育期的表達(dá)模式分析
由圖5 可知,隨著果實(shí)的發(fā)育成熟,黑果枸杞各WD40 蛋白質(zhì)編碼基因的表達(dá)模式存在差異,共有17個(gè)WD40 蛋白質(zhì)編碼基因隨果實(shí)的發(fā)育成熟表達(dá)上調(diào),其中Cluster-2917.0、Cluster-26021.67536、Cluster-11034.0、Cluster-20812.0 和Cluster-24260.0 基因的上調(diào)表達(dá)較明顯;共有21 個(gè)WD40 蛋白質(zhì)編碼基因在果實(shí)發(fā)育的青色期到轉(zhuǎn)色期表達(dá)上調(diào),而在果實(shí)發(fā)育的轉(zhuǎn)色期到成熟期表達(dá)下調(diào);共有13 個(gè)WD40 編碼基因隨果實(shí)的發(fā)育成熟表達(dá)下調(diào),其中Cluster-17972.0和Cluster-26021.148900 基因的表達(dá)下調(diào)較明顯;共有26 個(gè)WD40 蛋白質(zhì)編碼基因的表達(dá)在果實(shí)發(fā)育的青色期到轉(zhuǎn)色期、轉(zhuǎn)色期到成熟期均呈先降低后增加的趨勢(shì)。
3 討論
由于不同WD40 蛋白質(zhì)所含WD40 基序數(shù)目不同,且部分蛋白質(zhì)同時(shí)含有其他重要結(jié)構(gòu)域,導(dǎo)致WD40 蛋白質(zhì)結(jié)構(gòu)和功能的多樣化。Ouyang 等[25]對(duì)水稻OsWD40 基因進(jìn)行系統(tǒng)鑒定與共表達(dá)分析,根據(jù)水稻W(wǎng)D40 蛋白質(zhì)所含結(jié)構(gòu)域?qū)⑺網(wǎng)D40 蛋白質(zhì)分為11 類,包括僅含WD40 基序的WD40 蛋白質(zhì)145個(gè),同時(shí)包含LisH 結(jié)構(gòu)域、Utp12、Utp13、Utp15 和Utp21 結(jié)構(gòu)域、WDAD 或COPI 結(jié)構(gòu)域、NLE 結(jié)構(gòu)域、熱激蛋白重復(fù)體、BEACH 結(jié)構(gòu)域及BCAS3 結(jié)構(gòu)域及FBOX/U-BOX 結(jié)構(gòu)域的WD40 蛋白質(zhì)55 個(gè);Mishra等[9]在谷子全基因組中鑒定得到225 個(gè)SiWD40 基因,根據(jù)谷子WD40 蛋白質(zhì)所含結(jié)構(gòu)域?qū)⒐茸覹D40蛋白質(zhì)分為12 類,包括僅含WD40 基序的WD40 蛋白質(zhì)146 個(gè),同時(shí)包含其他結(jié)構(gòu)域的WD40 蛋白質(zhì)79個(gè)。本研究結(jié)果表明,黑果枸杞WD40 蛋白質(zhì)包括僅含WD40 基序的WD40 蛋白質(zhì)有38 個(gè),同時(shí)含其他結(jié)構(gòu)域的WD40 蛋白質(zhì)39 個(gè),這與前人研究結(jié)果一致,說明WD40 蛋白質(zhì)的結(jié)構(gòu)在物種間具有高度保守性。
大量研究表明,基因表達(dá)模式與基因功能密切相關(guān),如An 等[26]通過檢測(cè)蘋果WD40 蛋白質(zhì)編碼基因MdTTG1 在其根、葉、花及果皮中的表達(dá),發(fā)現(xiàn)其在蘋果紅色果皮中表達(dá)最高,將蘋果的WD40 蛋白質(zhì)編碼基因MdTTG1 在擬南芥中異位表達(dá),發(fā)現(xiàn)其可提高擬南芥花青素的積累;而Zhu 等[27]發(fā)現(xiàn)煙草WD40 蛋白質(zhì)編碼基因NtTTG2 表達(dá)沉默后,煙草花中的花青素含量顯著降低。本研究通過對(duì)黑果枸杞WD40 蛋白質(zhì)編碼基因在果實(shí)發(fā)育青色期、轉(zhuǎn)色期、成熟期的表達(dá)作分層聚類分析發(fā)現(xiàn),黑果枸杞WD40 蛋白質(zhì)編碼基因Cluster-2917.0 和Cluster-20812.0 的表達(dá)隨著果實(shí)發(fā)育不斷升高,且在成熟期較轉(zhuǎn)色期分別提高33 倍和10 倍以上,推測(cè)這2 個(gè)WD40 蛋白質(zhì)編碼基因可能正向調(diào)控黑果枸杞果實(shí)的顏色變化及花青素積累;由黑果枸杞與擬南芥WD40 蛋白質(zhì)共建的進(jìn)化樹得出,黑果枸杞WD40 蛋白質(zhì)Cluster-26021.105724、Cluster-26021.24543、Cluster-26021.46098 和Cluster-26021.86872 與擬南芥中參與花青素調(diào)控的WD40 蛋白質(zhì)進(jìn)化關(guān)系接近,同時(shí)結(jié)合黑果枸杞WD40 基因隨果實(shí)發(fā)育的表達(dá)模式中這些基因的表達(dá)趨勢(shì),進(jìn)一步證實(shí)了這些基因可能為花青素合成調(diào)控的候選基因。本試驗(yàn)結(jié)果為枸杞中WD40 蛋白質(zhì)編碼基因的分離和研究提供了一定的理論基礎(chǔ)。
4 結(jié)論
本研究基于黑果枸杞不同發(fā)育時(shí)期果實(shí)轉(zhuǎn)錄組測(cè)序結(jié)果,篩選注釋出77 個(gè)黑果枸杞WD40 蛋白質(zhì)編碼基因;分析其結(jié)構(gòu)域和結(jié)構(gòu)組成得出黑果枸杞WD40蛋白質(zhì)家族成員結(jié)構(gòu)保守;亞細(xì)胞預(yù)測(cè)結(jié)果顯示,黑果枸杞WD40 蛋白質(zhì)主要分布于細(xì)胞核及細(xì)胞質(zhì)中,其家族成員隨果實(shí)發(fā)育呈現(xiàn)不同的表達(dá)模式。本研究結(jié)果為枸杞中WD40 蛋白質(zhì)編碼基因的研究提供了一定的理論依據(jù)。下一步將通過實(shí)時(shí)熒光定量PCR 對(duì)WD40 蛋白質(zhì)編碼基因的表達(dá)進(jìn)行驗(yàn)證,并挑選隨果實(shí)顏色變化表達(dá)差異顯著的WD40 蛋白質(zhì)編碼基因,并對(duì)其功能進(jìn)行研究。
