姜黃素聯合KLF8基因siRNA調控JAK2/STAT3信號通路對乳腺癌細胞生長抑制作用的研究
2019-02-22 00:44:20李慶輝張成娟
安徽醫科大學學報 2019年1期
李 新 ,牛 冰 ,李慶輝 ,張成娟
Krüppel樣轉錄因子8(KLF8)是Krüppel樣轉錄因子(Krüppel like transcription factor,KLFs)家族成員之一,是表達廣泛的一個Krüppel樣轉錄因子,參與細胞的增殖、分化、凋亡、遷移、細胞外基質形成等多種生物學過程[1],研究[2-3]顯示,包括乳腺癌在內的多種腫瘤中KLF8呈現出過表達,而抑制KLF8后可降低腫瘤細胞生長,提示抑制乳腺癌中KLF8的表達可能影響其發生發展,但目前研究尚不清楚。
姜黃素(Curcumin)是姜黃的根莖中提取出來的一種酚性色素,具有明顯的抗腫瘤作用,有研究[4]顯示,姜黃素可明顯抑制多種腫瘤的生長及誘導凋亡。抑制KLF8基因表達與姜黃素聯合用于乳腺癌是否產生協同治療作用尚不清楚。因此,該研究通過RNA干擾抑制KLF8基因表達及姜黃素單獨或共同處理乳腺癌細胞,檢測對癌細胞活力和凋亡的影響,并進一步研究對其生物學特性影響的可能分子機制。
1 材料與方法
1.1主要試劑和儀器小牛血清及RPMI1640培養基購自美國Gibco公司;姜黃素購自美國Sigma公司;AG490購自美國Biosource公司;細胞計數試劑盒(cell counting kit-8,CCK-8)試劑盒和二喹啉甲酸(BCA)試劑盒購自上海碧云天公司;LipofectamineTM2000轉染試劑盒、Annexin V-FITC/PI細胞凋亡試劑盒購自美國Invitrogen公司;KLF8、磷酸化的蛋白酪氨酸激酶2(phosphorylated Janus kinase2,p-JAK2)、磷酸化的信號轉導與轉錄因子3(phosphorylated signal transducers and activators of transcription 3,p-STAT3)、細胞周期素D1(Cyclin D1)和B細胞淋巴瘤/白血病-2(B cell lymphoma/lewkmia-2,Bcl-2)抗體購自美國Abcam公司;酶標儀及流式細胞儀均購自美國Bio-Rad公司。
1.2細胞及其培養人乳腺癌MCF-7細胞購自中國科學院上海細胞庫;MCF-7細胞為貼壁細胞,細胞在含有小牛血清及雙抗的RPMI1640培養基中, 5% CO2、37 ℃飽和濕度培養箱中傳代培養。實驗選擇生長至對數期的細胞。
1.3細胞轉染MCF-7細胞進行常規消化后收集細胞,接種細胞于6孔板,轉染前1 d更換培養液(含有血清,不含抗生素),待細胞達70%~90%的生長密度時通過脂質體LipofectamineTM2000轉染試劑盒將合成的干擾KLF8表達的siRNA(KLF8-siRNA)轉染細胞,同時轉染無干擾作用的siRNA作為陰性對照組,并設置只加入脂質體的為空白對照組,轉染過程嚴格參照試劑盒說明,收集轉染48 h的細胞用于實驗研究。
1.4轉染KLF8-siRNA的MCF-7細胞KLF8的表達通過Western blot檢測轉染KLF8-siRNA的MCF-7細胞KLF8的表達。簡要步驟如下:PBS洗滌轉染48 h的細胞,離心后棄掉上清,加入全細胞蛋白裂解液于冰上反應30 min,離心收集上清液,上清液即為細胞總蛋白。BCA試劑盒對蛋白進行定量,蛋白經100 ℃變性,取等量變性蛋白進行SDS-PAGE分離,電泳結束后切下含有目的蛋白的膠通過電轉移至PVDF膜,5%脫脂奶粉在室溫條件下封閉轉好的PVDF膜2 h,封閉后置入適當比例的KLF8和GAPDH一抗溶液中,4 ℃緩慢搖動過夜,洗膜,加入二抗,室溫反應2 h,ECL發光后暗室中將X光片曝光,沖洗X光片,拍照掃描,成像分析軟件分析KLF8的相對表達量。
1.5CCK-8法檢測姜黃素聯合KLF8基因siRNA對MCF-7細胞活力的影響以每孔100 μl(約1×104個細胞)接種生長至對數期的MCF-7細胞于96孔板,用于調零組細胞的僅僅加入細胞培養液,細胞長滿50%~80%孔底時參照上述方法進行轉染,并隨機分為陰性對照組(轉染無干擾作用的siRNA)、KLF8-siRNA(轉染合成的干擾KLF8表達的siRNA)、姜黃素組(40 μmol/L姜黃素處理細胞)和KLF8-siRNA+姜黃素組(在轉染KLF8-siRNA的基礎上加入姜黃素),每組設置5個平行孔,培養48 h后,加入10 μl CCK-8試劑至每孔中,利用調零組調零,酶標儀測定吸光度值(optical density,OD),以OD值反映細胞活力,可以間接反映出細胞的增殖能力。實驗重復3次。
1.6流式細胞儀檢測姜黃素聯合KLF8基因siRNA對MCF-7細胞凋亡的影響分組及處理方法同1.5,PBS洗滌各組細胞,Binding緩沖液重懸細胞,利用Annexin V-FITC和PI熒光染色,室溫避光反應15 min,流式細胞儀進行檢測。實驗重復3次。
1.7姜黃素聯合KLF8基因siRNA抑制JAK2/STAT3信號通路對乳腺癌細胞活力及凋亡的影響AG490作為JAK2/STAT3信號通路抑制劑,加入濃度為20 μmol/L,通過CCK-8法及流式細胞儀檢測用KLF8-siRNA+姜黃素+AG490處理細胞相對于KLF8-siRNA+姜黃素處理的細胞活力及凋亡率情況,Western blot檢測p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表達,方法同1.4。

2 結果
2.1KLF8-siRNA轉染MCF-7細胞后KLF8的表達KLF8-siRNA轉染MCF-7細胞后KLF8的表達結果見圖1。與空白對照組(0.311±0.035)比較,KLF8-siRNA組(0.102±0.010)KLF8的表達顯著降低(P<0.05),而陰性對照組(0.302±0.032)和空白對照組KLF8的表達差異無統計學意義(P>0.05)。

圖1 KLF8-siRNA轉染MCF-7細胞后KLF8的表達
2.2姜黃素聯合KLF8-siRNA對MCF-7細胞活力的影響CCK-8檢測結果顯示:KLF8-siRNA組(0.562±0.048)和姜黃素組(0.626±0.052)的細胞活力均顯著低于陰性對照組(0.873±0.074)(P<0.05),高于KLF8-siRNA+姜黃素組(0.385±0.041)(P<0.05)。
2.3姜黃素聯合KLF8-siRNA對MCF-7細胞凋亡的影響流式細胞儀檢測結果顯示:KLF8-siRNA組(12.12±1.02)%和姜黃素組(10.77±0.96)%的細胞凋亡率均顯著高于陰性對照組(1.56±0.16)%(P<0.05),低于KLF8-siRNA+姜黃素組(18.79±1.35)%(P<0.05)。見圖2。
2.4姜黃素聯合KLF8-siRNA對MCF-7細胞p-JAK2、p-STAT3、CyclinD1和Bcl-2表達的影響Western blot檢測結果顯示:KLF8-siRNA組和姜黃素組Cyclin D1、Bcl-2、p-JAK2和p-STAT3的蛋白表達均顯著低于陰性對照組(KLF8-siRNA組:P<0.05;姜黃素組:P<0.05),高于KLF8-siRNA+姜黃素組(KLF8-siRNA組:P<0.05;姜黃素組P<0.05)。四組間JAK2和STAT3的蛋白表達差異無統計學意義。見表1、圖3。
2.5姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號對MCF-7細胞活力及凋亡的影響姜黃素、KLF8-siRNA及JAK2/STAT3信號抑制劑AG490共同處理細胞后,細胞OD值及凋亡率檢測結果顯示:與KLF8-siRNA+姜黃素組[(0.422±0.043)、(16.68±1.21)%]比較,KLF8-siRNA+姜黃素+AG490組OD值(0.301±0.032)顯著降低(P<0.05),凋亡率顯著升高[(21.15±1.54)%](P<0.05)。見圖4。

表1 姜黃素聯合KLF8-siRNA對MCF-7細胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表達的影響
與陰性對照組比較:*P<0.05;與KLF8-siRNA+姜黃素組比較:#P<0.05

圖2 姜黃素聯合KLF8-siRNA對MCF-7細胞凋亡的影響

圖3 姜黃素聯合KLF8-siRNA對MCF-7細胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表達的影響
A:陰性對照組;B:KLF8-siRNA組;C:姜黃素組;D:KLF8-siRNA+姜黃素組
2.6姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號對MCF-7細胞p-JAK2、p-STAT3、CyclinD1和Bcl-2表達的影響姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號后MCF-7細胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表達結果見表2、圖5。與KLF8-siRNA+姜黃素組比較,KLF8-siRNA+姜黃素+AG490組p-JAK2、p-STAT3、Cyclin D1和Bcl-2的蛋白表達均顯著降低(P<0.05)。

表2 姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號對MCF-7細胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表達的影響
與KLF8-siRNA+姜黃素組比較:*P<0.05
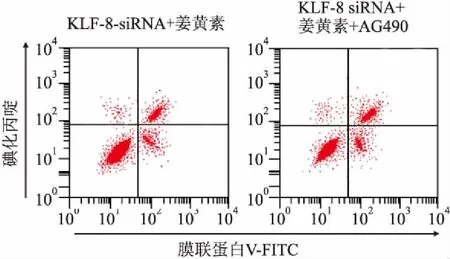
圖4 姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號對MCF-7細胞活力及凋亡的影響
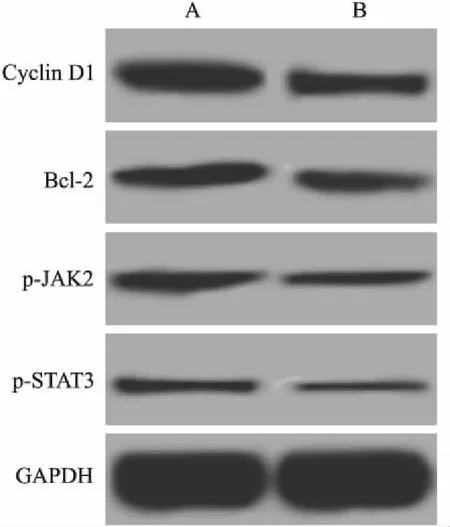
圖5 姜黃素聯合KLF8-siRNA抑制JAK2/STAT3信號對MCF-7細胞p-JAK2、p-STAT3、Cyclin D1和Bcl-2表達的影響
A:KLF8-siRNA+姜黃素組;B:KLF8-siRNA+姜黃素+AG490組
3 討論
研究[5]顯示,在多種腫瘤中,KLFs家族的多個成員與癌基因相關。KLF8是KLFs家族成員之一,在上皮細胞-間充質轉化(epithelial cell mesenchyme transformation,EMT)、細胞致癌性分化、細胞侵襲和遷移、周期循環等過程中均有重要作用,且在多種腫瘤中過表達,其過表達與疾病不良預后及發生相關[6]。也有研究[7]指出抑制KLF8的表達可降低腫瘤的發生發展,如抑制腎癌細胞KLF8表達可降低癌細胞增殖、侵襲及誘導細胞凋亡。KLF8在乳腺癌中的研究相對較少,研究[8]顯示,KLF8在乳腺癌中表達上調,可通過活化基質金屬蛋白9促進乳腺癌的侵襲及轉移。本研究旨在抑制KLF8對乳腺癌增殖凋亡的影響。我國中藥資源豐富,中藥提取物在腫瘤治療的研究受到廣泛關注,姜黃素是姜黃的根莖中提取出來的一種酚性色素,目前在腫瘤中的作用有大量研究,研究[9]顯示,姜黃素可降低乳腺癌的增殖、侵襲、遷移,阻滯細胞周期和誘導細胞的凋亡,但聯合使用姜黃素和KLF8的siRNA是否能更有效地治療乳腺癌還未清楚。
鑒于有研究[2]已證實乳腺癌中KLF8存在高表達,本研究檢測抑制KLF8表達對乳腺癌細胞的影響。由于RNA干擾技術能在基因的轉錄和翻譯水平及染色質水平調節基因的表達,且表現出很強的序列特異性、有效性,在多種腫瘤中研究基因功能時被應用[10],因此本研究也選用RNA干擾技術抑制乳腺癌中KLF8的表達。將KLF8的siRNA和姜黃素共同處理乳腺癌細胞,結果顯示KLF8-siRNA和姜黃素均能明顯抑制乳腺癌細胞活力和誘導細胞凋亡,這提示KLF8基因和姜黃素能協同用于乳腺癌的防治。信號轉導與轉錄因子3(signal transducers and activators of transcription 3, STAT3)是STATs家族的一員,與細胞增殖、凋亡等生物學特性密切相關,在多種腫瘤中出現異常活化,STAT3的異常活化可引起細胞異常增殖分化,且凋亡受到抑制,目前已被確認為癌基因,STAT3引起癌變的機制主要是通過激活CyclinD1、Bcl-2、Survivin等靶基因的一些產物表達實現[11]。CyclinD1是一個細胞周期核因子,可促進細胞的增殖[12]。Bcl-2是Bcl-2家族成員之一,抑制其表達可誘導細胞的凋亡[13]。JAK2處在STAT3上游,JAK2激酶抑制劑AG490可抑制JAK2/STAT3信號通路,從而使細胞增殖受到抑制,AG490可抑制包括乳腺癌在內的多種腫瘤的發生發展[14]。本研究顯示,姜黃素和KLF8的siRNA均能下調p-JAK2和p-STAT3及靶基因CyclinD1和Bcl-2表達,兩者聯合下調更明顯,抑制JAK2/STAT3信號通路后,相對于聯合使用姜黃素和KLF8的siRNA對細胞的抑制作用和凋亡促進作用更明顯。
綜上所述,下調KLF8基因表達和姜黃素均能通過抑制JAK2/STAT3信號通路降低乳腺癌細胞活力,誘導細胞凋亡,兩者聯合對細胞活力及凋亡的影響作用更強。該研究可能為乳腺癌的治療提供了新的途徑,值得進一步深入探討。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中國生殖健康(2019年2期)2019-08-23 08:11:42
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
