γ-氨基丁酸對高溫脅迫下黑麥草光合特性及碳水化合物代謝的影響
2019-02-25 01:57:14,,
草業學報 2019年2期
關鍵詞:植物
,,
(1.北部灣大學資源與環境學院,廣西 欽州535011;2.湖南芷蘭生態環境建設有限公司, 湖南 長沙410000;3.湖南農業大學農學院草業科學系,湖南 長沙 410128)
草坪和草坪草在現代城市發展和人們日常生活中發揮著越來越重要的作用。溫度是影響草坪草生長發育的重要環境因子,在南方亞熱帶過渡性氣候區6-8月份氣溫長期高達35~38 ℃, 而對于大多數的冷季型草坪草種和品種來說都存在難以越夏的問題。多年生黑麥草(Loliumperenne)是庭園和城市園林綠化以及足球場運動草坪建植中常用的冷季型草坪草種,因其具有草坪質量優良、分蘗性能好、成坪迅速等優點而常用作草坪建植的先鋒草種。黑麥草的最適生長溫度為15~25 ℃,當氣溫長時間高于35 ℃時草坪的生長發育受阻而造成草坪質量下降甚至死亡[1-2]。
光合作用是植物對外界環境條件最為敏感的生理反應之一。光合作用的各個環節包括光能的吸收、傳遞和轉換等光反應過程、CO2氣體交換及固定和還原等暗反應過程常受到高溫脅迫的影響[3]。高溫脅迫下植物光合作用的下降,既有氣孔因素的限制,同時也受高溫脅迫下植物細胞內PSⅡ電子傳遞和反應中心結構等受損、Rubisco羧化效率和RuBP再生速率下降等非氣孔因素的限制[4]。糖是植物生長發育過程中的重要能量提供者,同時也對植物的抗逆性具有重要調控作用。植物體內非結構性糖既是植物細胞內代謝的重要底物,也是光合與生長利用之間主要的中間貯存產物[5]。可溶性糖不僅可作為滲透調節物質,還可以作為抗氧化劑以維持和保護逆境條件下植物細胞膜結構的完整性[6]。高溫脅迫下植物葉片光合能力降低,光合產物減少,從而糖代謝發生改變,而蔗糖合成酶(sucrose synthase,SS)、蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)和轉化酶(invertase,INV)是糖代謝過程中的關鍵酶,對細胞內蔗糖、葡萄糖和果糖的合成與分解起著關鍵性的作用[7]。
γ-氨基丁酸(γ-aminobutyric acid,GABA)是一種在動植物體內均有分布的四碳非蛋白質氨基酸,可作為外源小分子物質參與植物的生長發育和逆境脅迫調控[8]。當植物受到水分、溫度、鹽堿等一些逆境脅迫時,植物體內GABA的含量大量增加而抗逆性提高[9]。外源GABA 既可作為臨時氮庫,還可以清除活性氧和自由基、調節細胞內的pH和滲透壓等作用以提高植物的抗逆性[10]。已有研究表明,外源GABA 能夠提高番茄(Lycopersiconesculentum)[10-11]、網紋甜瓜(Cucumismelo)[12]、小麥(Triticumaestivum)[13]、玉米(Zeamay)[14]、白三葉(Trifoliumrepens)[15]、黃瓜(Cucumissativus)[16]等植物的抗旱、耐鹽、耐寒等抗逆能力。Li 等[17]研究發現外源施用GABA可通過調控氨基酸、有機酸的代謝水平而提高冷季型草坪草匍匐剪股穎(Agrostisstolonifera)的耐旱和耐熱能力,但是關于外源GABA 能否通過調控光合和葉綠素熒光特性及碳水化合物代謝水平來提高黑麥草耐高溫能力的研究還未見報道。因此,本研究在前期試驗的基礎上,以耐熱性弱的黑麥草品種為試驗材料,通過噴施和澆灌外源GABA,研究了外源GABA對高溫脅迫下黑麥草的生長、光合作用、葉綠素熒光特性、碳水化合物含量及其代謝關鍵酶基因表達的影響,為解析外源GABA增強植物的抗逆性和多年生黑麥草的抗逆機理提供理論基礎。
1 材料與方法
1.1 植物材料
采用耐熱性弱的多年生黑麥草品種‘頂峰’(L.perennecv. Pinnacle)為材料[18]于2017年9-11月進行試驗。
1.2 試驗設計與處理
黑麥草品種‘頂峰’種子經發芽試驗確定發芽率,然后將種子播種于裝有栽培基質的塑料花盆中(上口徑15 cm×下口徑10 cm×高15 cm),培養基質為泥炭土和河砂(1∶1,體積比),然后置于湖南農業大學教學科研試驗溫室中培養。幼苗長至高約8 cm時進行修剪,修剪高度保持約6 cm,每周每盆澆一次1/2濃度的Hoagland營養液,每次澆200 mL。
將在溫室中培養至冠層和根系基本建成(約45 d)的各盆栽材料轉移至光照培養箱中進行預培養,培養條件設為晝/夜溫度為25 ℃/20 ℃,65%~75%的相對濕度,12 h光照時間和400 μmol·m-2·s-1的光照強度。前期預試驗結果發現5 mmol·L-1GABA對黑麥草緩解高溫脅迫具有最好的效果。盆栽材料在生長箱預培養3 d后,用5 mmol·L-1的GABA溶液每盆噴施100 mL并同時澆灌150 mL,2 d后開始進行高溫處理。對照晝夜溫度設為25 ℃/20 ℃,高溫處理晝夜溫度設為35 ℃/30 ℃,其余培養條件跟預培養相同。試驗完全隨機設計,每個處理設4次重復(4盆)。隔天澆水一次以保持土壤濕潤。在高溫脅迫處理后的第4、12、24 h和第5、15天取倒數第3~5片功能葉片,液氮速凍后保存于-80 ℃冰箱用于基因表達分析;在高溫處理后的第5 和15天進行冠層高度、葉片電導率、葉綠素熒光和光合作用測定,同時取倒數第3~5片功能葉片,液氮速凍后保存于-80 ℃冰箱用于丙二醛和碳水化合物測定分析。
1.3 測定項目與方法
1)冠層高度:在高溫處理前和高溫處理的第5和15天用直尺測定各盆栽材料的冠層高度(cm)。
2)細胞膜穩定性:用電導率表示,根據Hu等[19]的方法進行測定。
3)丙二醛含量(MDA):采用pH 7.8的磷酸緩沖液提取,然后根據Hu等[19]的方法用分光光度計(UV7500,上海)進行測定。
4)葉綠素含量:葉片經二甲基亞砜在室溫避光條件下浸提后采用分光光度計法進行測定[20]。
5)葉綠素熒光:葉片經暗適應30 min后,采用德國WALZ公司生產的PAM 2500葉綠素儀進行最大光化學效率(Fv/Fm)、光化學猝滅系數(qP)、非光化學淬滅系數(NPQ)、光合電子傳遞速率(ETR)、PSⅡ實際光化學效率(ΦPSⅡ)等葉綠素熒光參數的測定。
6)光合作用:參考Hu 等[21]的方法,在上午9: 00-11: 00,采用Li-6400光合儀直接測定葉片凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr)和CO2響應曲線(Pn-CO2);測定光合作用時設定光強為600 μmol·m-2·s-1,CO2濃度為400 μmol·mol-1,溫度為25 ℃;用直線回歸法求得Pn-CO2響應曲線的初始斜率,即為CO2羧化效率(CE), 根據Pn-Ci曲線計算氣孔限制度(Ls):Ls=100%×(Aca-Aci)/Aca(ca 為大氣CO2濃度),同時用Photosynthesis Assistant軟件計算最大光合速率(Amax)、CO2補償點(CP)、Rubisco最大羧化速率(Vcmax)和RuBP再生速率(Jmax)。
7)碳水化合物測定:樣品烘干后研磨過0.425 mm篩,用92%的乙醇提取后按王正文[22]的方法用HPLC測定果糖、葡萄糖、蔗糖和淀粉的含量。
8)基因表達分析:總RNA采用Trizol試劑盒參考說明書進行提取,經Nanodrop 2000檢測濃度和純度以及用1%瓊脂糖凝膠電泳檢測完整性后,根據Fermentas cDNA synthesis 試劑盒說明書進行反轉錄合成cDNA。參考TaKara 公司的SYBR System操作手冊,采用ABI7500實時熒光定量PCR儀進行PCR擴增。每個處理設3個重復。基因特異性引物序列參考表1進行合成[23],以YT521-B基因為內參。根據得到的Ct值,利用2-ΔΔCT法,分別計算目標基因在不同處理下的相對表達量。
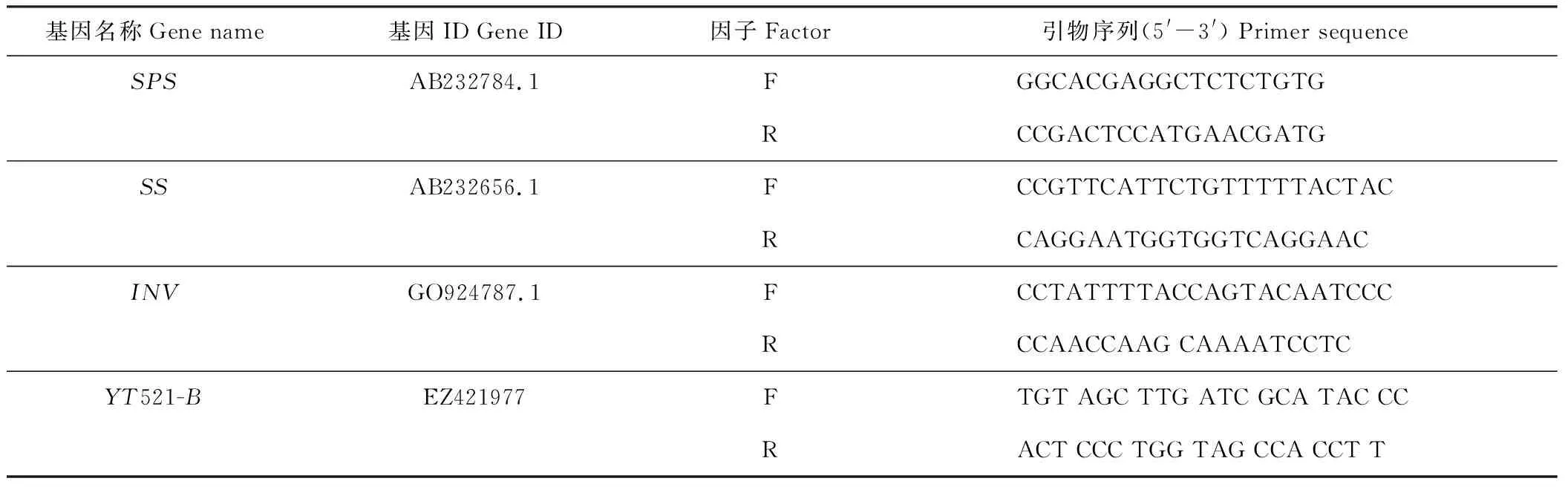
表1 試驗中所用基因及其引物序列Table 1 Genes and primers used in the experiment
SPS: 蔗糖磷酸合成酶Sucrose phosphate synthase;SS: 蔗糖合成酶Sucrose synthetase; INV: 轉化酶Invertase; invertin;YT521-B: 看家基因House-keeping gene;F: 正面Forward; R: 反向Reverse.
1.4 數據處理與分析
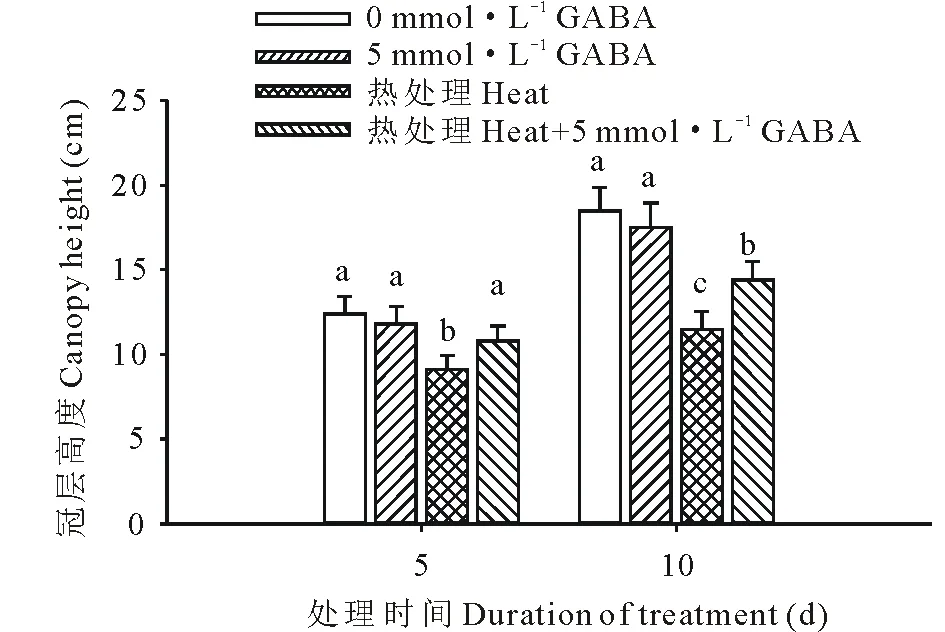
圖1 外源GABA對高溫脅迫下黑麥草冠層高度的影響Fig.1 Effect of exogenous GABA on the canopy height in perennial ryegrass under heat stress 圖中豎線表示平均值±標準誤,不同小寫字母表示在同一處理時間內不同處理間差異顯著(P<0.05),下同。Vertical bars indicate means±standard error (SE). Different lowercase letters indicate significant difference among treatment at a given day of treatment, the same below.
采用SAS 軟件進行數據統計分析,然后采用Duncan’s多重比較檢測處理間的差異顯著性(P<0.05),采用Sigmaplot 12.0作圖。
2 結果與分析
2.1 GABA對高溫脅迫下黑麥草冠層高度的影響
由圖1可以看出,高溫脅迫下黑麥草生長顯著受到抑制,冠層高度在高溫處理的第5 和15天時比對照低,分別為27%和38%。但是外源施用GABA顯著緩解了高溫脅迫對黑麥草生長的抑制,在高溫處理第5 和15天時施用GABA的處理冠層高度比未施GABA的處理分別了提高了16%和20%。
2.2 GABA對高溫脅迫下黑麥草葉片電導率的影響
由圖2可知,在高溫脅迫下黑麥草葉片的相對電導率顯著升高,在脅迫的第5和15天時電導率分別提高了31%和78%;但是高溫脅迫下外源施用GABA后,黑麥草葉片電導率顯著降低,與高溫脅迫下未施用GABA的處理相比,在高溫處理的第5和15 天分別下降了30%和28%。
2.3 GABA對高溫脅迫下黑麥草葉綠素含量的影響
從圖3可以看出,高溫脅迫下黑麥草葉片的葉綠素含量顯著下降,與對照相比,在脅迫處理的第5和15天時的含量分別下降了9%和37%;外源施用GABA后則顯著緩解了因高溫而造成的葉綠素含量的下降,在高溫處理后的第5和15天,施用外源GABA的處理比未施GABA的處理葉綠素含量則分別提高了9%和23%。
2.4 GABA對高溫脅迫下黑麥草葉片葉綠素熒光參數的影響
從表2可以看出,正常溫度下施用外源GABA短期內(5 d)提高了黑麥草的ETR,但對Fv/Fm、ΦPSⅡ、qP和NPQ無顯著影響,但較長時間(15 d)后對ETR、Fv/Fm、ΦPSⅡ、qP和NPQ均無顯著影響。短時間高溫(5 d)對黑麥草葉片的Fv/Fm無顯著影響,較長時間高溫(15 d)下黑麥草Fv/Fm顯著下降;高溫脅迫下黑麥草葉片的ETR、qP和ΦPSⅡ顯著降低,而NPQ顯著升高;但高溫脅迫下施用外源GABA不論是在短期(5 d)還是較長時間(15 d)內均顯著提高了黑麥草葉片的Fv/Fm、ETR、qP和ΦPSⅡ,而降低了NPQ;與高溫下未施用GABA相比,在處理5 d時,ETR、qP和ΦPSⅡ分別提高了26%,9%和13%,而NPQ則降低了25%;但到高溫處理15 d,Fv/Fm、ETR、qP和ΦPSⅡ則分別提高了17%,50%,21%和18%,而NPQ則降低了32%。
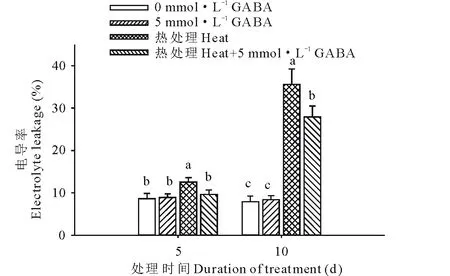
圖2 外源GABA對高溫脅迫下黑麥草葉片電導率的影響Fig.2 Effect of exogenous GABA on the leaf electrolyte leakage in perennial ryegrass under heat stress
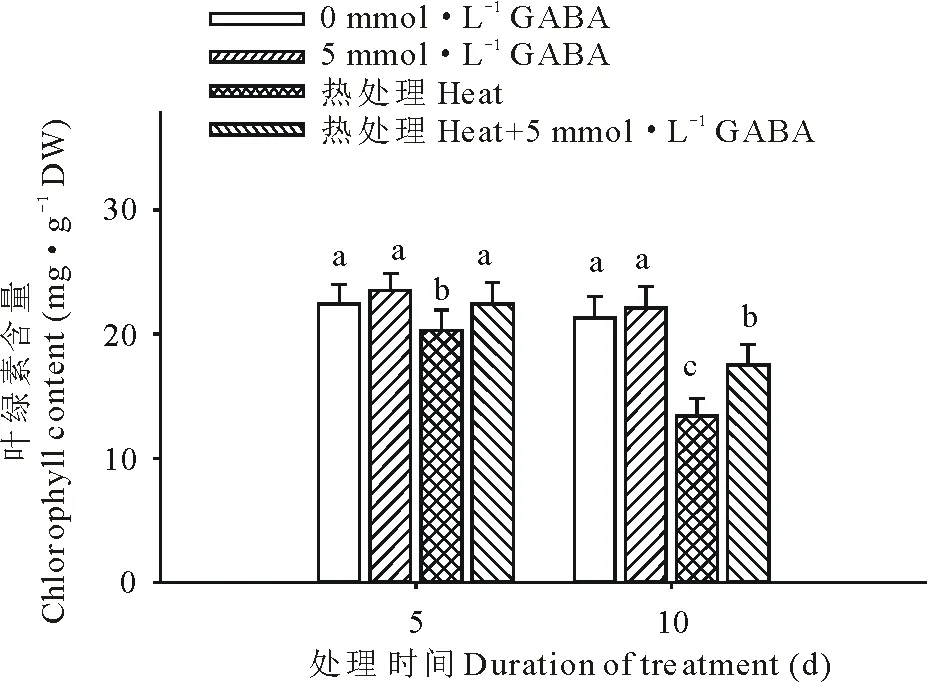
圖3 外源GABA對高溫脅迫下黑麥草葉綠素含量的影響Fig.3 Effect of exogenous GABA on the leaf chlorophyll content in perennial ryegrass under heat stress
注:同列中不同小寫字母表示同一處理時間不同處理間在P<0.05水平上差異顯著,下同。
Note: Means in a column marked with different lower-case letters indicate significant difference among treatment at a given day of treatment atP<0.05 level, the same below.
2.5 GABA對高溫脅迫下黑麥草葉片光合參數的影響
由表3可知,施用外源GABA對正常溫度下黑麥草的Pn、Gs、Ci和Tr均無顯著影響。高溫脅迫下除了較長時間(15 d)處理下Ci外,黑麥草葉片的Pn、Gs和Tr顯著降低,在處理5 d時,分別降至對照的64%,52%和67%,到處理15 d時,則分別僅為對照的35%,37%和55%;高溫下黑麥草葉片的Ci在短期內(5 d)顯著降低,而較長時間(15 d)處理下則與對照沒有顯著性差異。高溫脅迫下施用外源GABA顯著提高了黑麥草葉片的Pn、Gs和Tr,與未施GABA的處理相比,在5 d時分別提高了31%,45%和18%,在處理15 d時則分別提高了73%,45%和34%;與高溫脅迫下未施GABA的處理相比,施用外源GABA在5 d時顯著提高了黑麥草葉片的Ci,而在處理15 d時則顯著降低了葉片Ci。
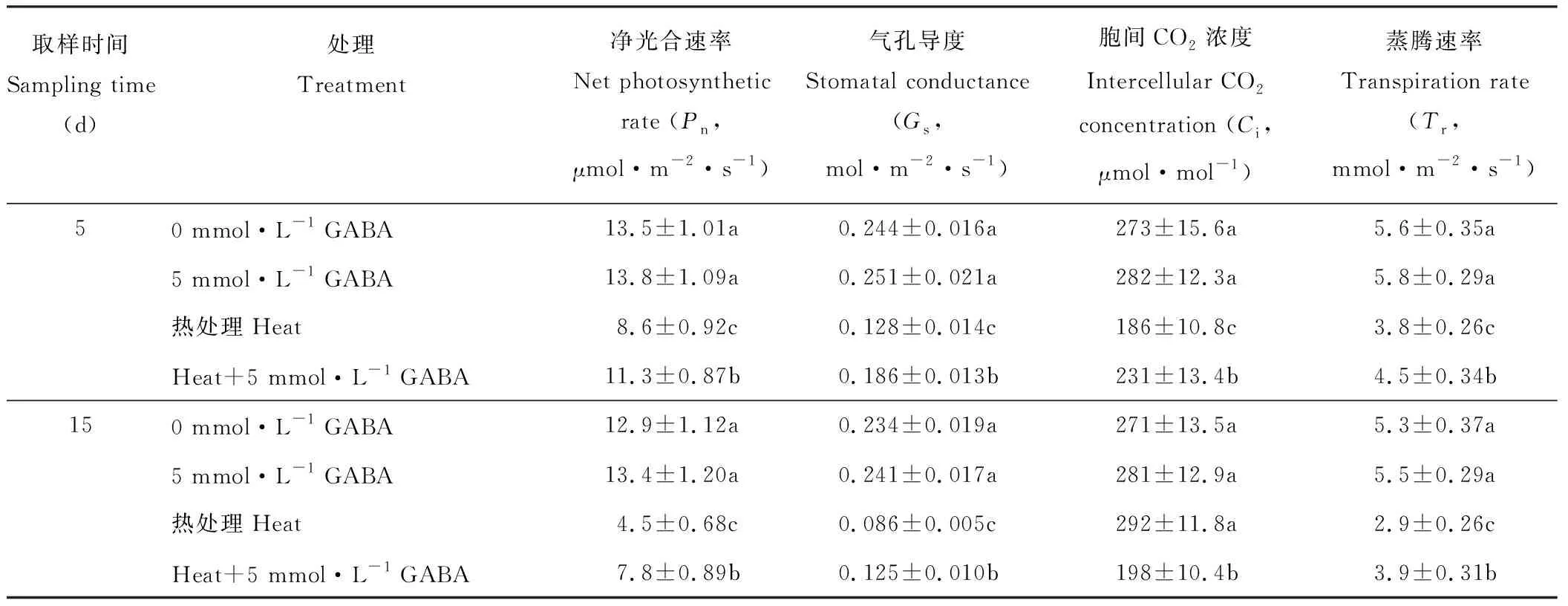
表3 外源GABA對高溫脅迫下黑麥草葉片光合作用的影響Table 3 Effect of exogenous GABA on leaf photosynthesis in perennial ryegrass under heat stress
由表4可知,在短期(5 d)處理條件下,高溫脅迫和施用外源GABA對黑麥草葉片的CE、Vcmax和Jmax均無顯著影響;處理較長時間(15 d)時,高溫脅迫顯著降低了黑麥草葉片的CE、Vcmax和Jmax,但是施用外源GABA則顯著緩解了高溫條件下CE、Vcmax和Jmax的下降,比未施用GABA的處理分別提高了70%,31%和23%。高溫脅迫顯著提高了黑麥草葉片的Ls和CP,降低了Amax,但是施用外源GABA則顯著緩解了因高溫脅迫而造成Amax的下降以及Ls和CP的升高。
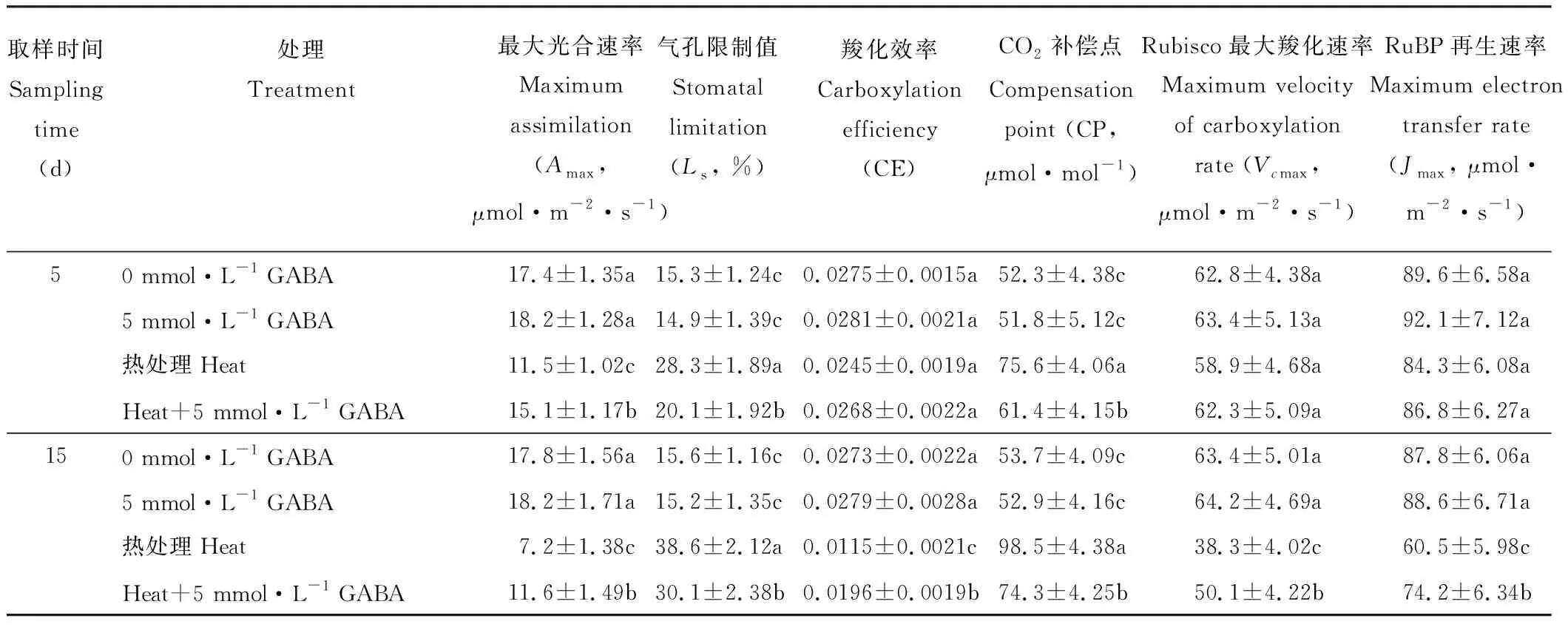
表4 外源GABA對高溫脅迫下黑麥草葉片光合特性的影響Table 4 Effect of exogenous GABA on leaf photosynthetic traits in perennial ryegrass under heat stress
2.6 GABA對高溫脅迫下黑麥草葉片碳水化合物含量的影響
從表5可以看出,在短期(5 d)處理條件下,高溫脅迫和施用外源GABA對黑麥草葉片的果糖含量無顯著影響;處理較長時間(15 d)時,高溫脅迫顯著降低了黑麥草葉片的果糖含量,但是施用外源GABA則顯著緩解了高溫條件下果糖含量的下降。高溫脅迫處理顯著降低了黑麥草葉片的葡萄糖、蔗糖和淀粉含量,在高溫處理5 d時,分別降低至對照的76%,79%和73%,在處理15 d時則分別降低至對照的66%,64%和53%;高溫處理下施用外源GABA顯著提高了黑麥草葉片葡萄糖、蔗糖和淀粉的含量,與未施GABA處理相比,在處理5 d時,分別提高了17%,18%和19%,在處理15 d時,則分別提高了24%,22%和40%。
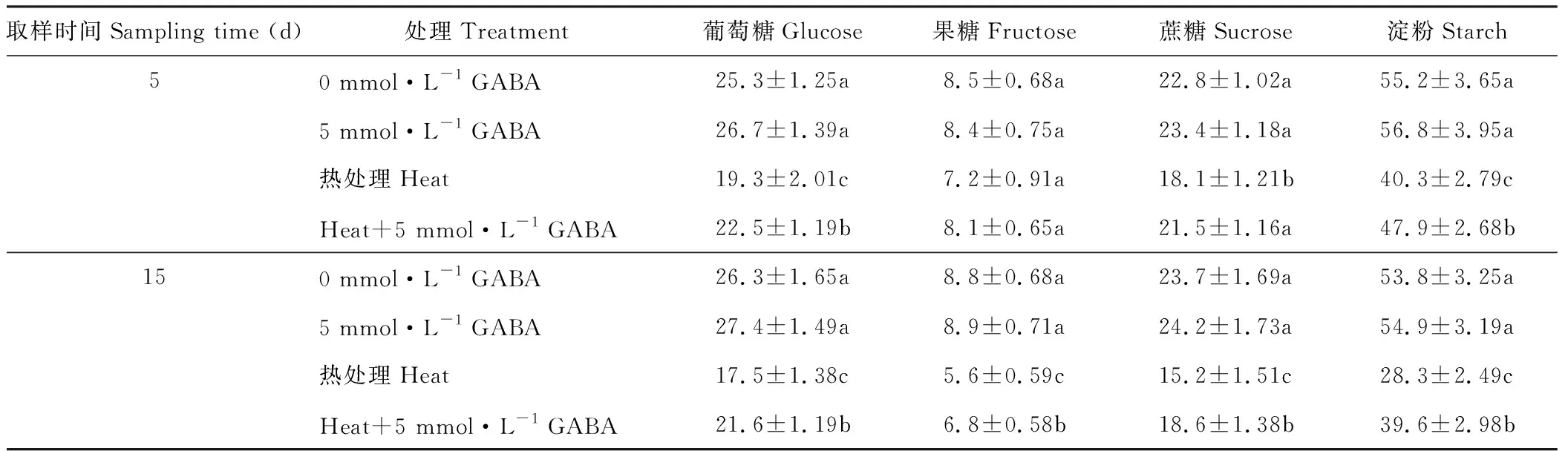
表5 外源GABA對高溫脅迫下黑麥草葉片碳水化合物含量的影響Table 5 Effect of exogenous GABA on leaf carbohydrates content in perennial ryegrass under heat stress (mg·g-1 DW)
2.7 GABA對高溫脅迫下黑麥草葉片糖代謝關鍵酶基因表達的影響
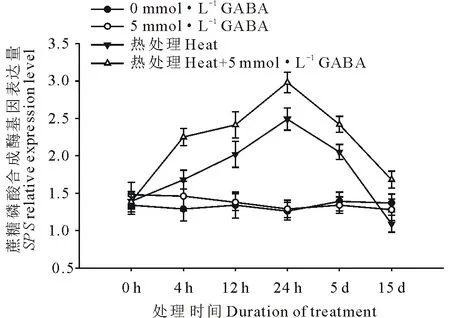
圖4 外源GABA對高溫脅迫下黑麥草葉片蔗糖磷酸合成酶基因表達的影響Fig.4 Effect of exogenous GABA on the expression level of SPS genes in leaves of perennial ryegrass under heat stress
由圖4和圖5可知,高溫誘導了黑麥草葉片蔗糖磷酸合成酶基因(SPS)和蔗糖合成酶基因(SS)的上調表達,并且隨著處理時間的延長表達水平呈顯著上升趨勢,在高溫處理24 h時達到頂峰,隨后呈緩慢下降趨勢,在處理15 d時表達量降至對照水平;在正常生長條件下,外源施用GABA對SPS和SS基因的表達水平沒有顯著影響;但在高溫脅迫條件下,與未施用GABA的處理相比,外源施用GABA則顯著誘導和提高了黑麥草葉片SPS和SS基因的表達水平,在15 d的處理時間內均顯著高于正常對照和高溫下未施用GABA的處理。
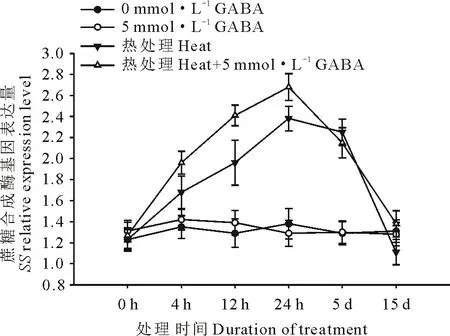
圖5 外源GABA對高溫脅迫下黑麥草葉片蔗糖合成酶基因表達的影響Fig.5 Effect of exogenous GABA on the expression level of SS genes in leaves of perennial ryegrass under heat stress
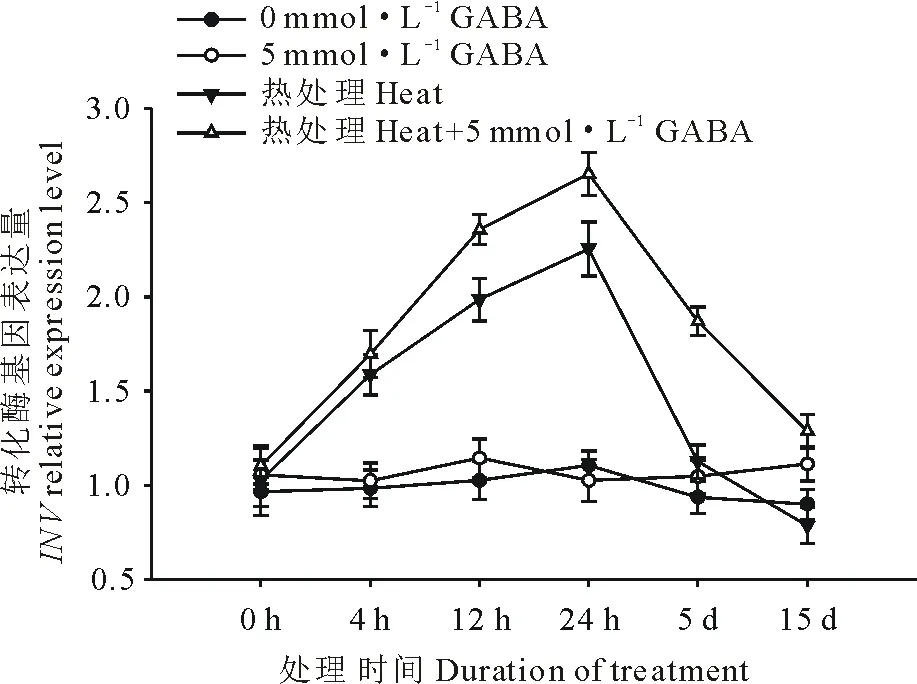
圖6 外源GABA對高溫脅迫下黑麥草葉片轉化酶基因表達的影響Fig.6 Effect of exogenous GABA on the expression level of INV gene in leaves of perennial ryegrass under heat stress
由圖6可以看出,高溫脅迫誘導了黑麥草葉片轉化酶基因(INV)的上調表達,且隨著處理時間的延長表達量上升,在處理24 h到達頂峰,隨后下降,在處理5 d以后則降至對照水平。在正常生長條件下,外源施用GABA對INV基因的表達水平沒有顯著影響;在高溫脅迫條件下,與未施用GABA的處理相比,外源施用GABA則更顯著地提高了黑麥草葉片INV基因的表達水平,除了在處理4 h以外,在15 d的處理時間內均顯著高于正常對照和高溫下未施用GABA的處理。
3 討論
3.1 外源GABA緩解了高溫脅迫對黑麥草的傷害
高溫常造成冷季型草坪草生長發育受阻、草坪質量下降、細胞膜受傷害、葉片枯黃衰老等變化,而植物的生長速率和電導率常作為衡量植物受高溫脅迫損害程度和抗逆性大小的重要指標[24]。GABA可以作為外源物質參與到植物的生長發育以及逆境調控等一系列的生理過程和抗逆能力[25]。本試驗結果顯示,高溫脅迫下黑麥草的生長顯著受到抑制,葉片電導率升高,但是施用外源GABA后黑麥草的冠層高度增加、細胞膜透性下降,這表明外源GABA對黑麥草在高溫脅迫下的生長具有促進作用,施用外源GABA可以顯著提高黑麥草的耐高溫能力,這與在冷季型草坪草匍匐剪股穎中的研究結果相一致[17]。
3.2 外源GABA減輕了高溫對黑麥草光合器官的損傷和光抑制,提高了光合碳同化能力
光合作用是植物對外界環境因子最敏感的生理過程之一,在一定程度上可反映植株對逆境脅迫的抗性[3]。高溫脅迫下植物葉片光合作用的降低既受氣孔因素的限制,同時也受非氣孔因素的影響[4]。根據Farquhar等[26]觀點,氣孔因素引起的Pn降低表現為Gs和Ci都降低,而非氣孔因素引起Pn降低則伴隨著Gs的下降而Ci升高。本試驗結果顯示,在高溫脅迫5 d時,黑麥草葉片的Pn隨著Gs和Ci下降而降低,且CE、Vcmax和Jmax無顯著性變化,但在高溫脅迫15 d時Pn和Gs顯著下降,而Ci則沒有下降甚至升高,并且CE、Vcmax和Jmax均顯著降低,表明在短時間高溫或輕度高溫脅迫時,Pn的下降主要受氣孔導度的下降等氣孔因素的影響,而在較長時間的高溫脅迫或重度高溫脅迫下光合作用的下降主要是受CE、Vcmax和Jmax等非氣孔因素的限制,這與前人的研究結果基本一致[27-28]。已有研究證明外源噴施GABA能調節鹽堿等逆境條件下植物葉片的光合性能[29]。本研究結果顯示,高溫脅迫下施用外源GABA后黑麥草葉片的Pn顯著提高,緩解了高溫對Gs和Tr的抑制,減少了Ci的積累,此外還顯著提高了Amax、CE、Vcmax和Jmax,表明外源GABA可通過緩解高溫對葉片氣孔和非氣孔因素的限制,從而改善了黑麥草葉片的光合特性,提高了植株的耐熱性,這與黃娟等[30]在黃瓜上的研究結果一致。
葉綠素熒光參數對環境逆境脅迫反應敏感,可反映環境因子對植物光合生理狀況的影響[31]。因此,對葉片葉綠素熒光參數的測量成為評價逆境脅迫對植物光合器官損傷的重要指標[32]。本試驗結果顯示,高溫脅迫下黑麥草葉片的Fv/Fm、qP、ETR和ΦPSⅡ顯著下降,而NPQ顯著升高,表明在高溫脅迫下黑麥草葉片的光系統Ⅱ活性中心受到了傷害,光能轉換效率和電子傳遞能力下降,光能利用率降低,光能過剩加劇了對葉片光合的抑制和光合器官的破壞,從而造成了葉綠素熒光參數的下降,這與在冷季型草坪草高羊茅(Testucaarundinacea)中的研究相一致[27]。維持植物光合器官結構的穩定,提高葉片光合能力對抵抗逆境脅迫至關重要。前人研究表明[11],施用GABA可提高低氧脅迫下甜瓜葉片和鹽脅迫下番茄葉片的光合能力。在本研究中,外源施用GABA顯著提高了高溫脅迫下黑麥草葉片的Fv/Fm、qP、ETR和ΦPSⅡ,降低了NPQ,表明外源GABA可提高光系統Ⅱ反應中心的光能轉換效率、潛在活性和開放比例,同時還增強了過剩光能的耗散,有利于減輕高溫脅迫對光合的抑制,增強光合系統反應中心的穩定性,從而減輕高溫脅迫對黑麥草葉片光合器官造成的損傷,提高了高溫脅迫下黑麥草的光合能力和耐熱性。
3.3 外源GABA提高了高溫脅迫下黑麥草葉片碳水化合物代謝水平
不同的植物種類對溫度的敏感性和耐受性不同,一定范圍內的高溫可以促進植物體內的生理生化反應,從而促進光合和碳水化合物代謝,而過高的溫度便可降低光合作用,增強了呼吸作用而消耗了更多的碳水化合物等能量物質使其含量顯著降低[33]。逆境條件下可溶性糖(包括蔗糖、果糖、葡萄糖等)的增加能夠增加細胞汁液濃度,有利于細胞結構的穩定,從而能夠維持逆境下細胞內正常的生理代謝而提高抗逆性[34]。前期的研究結果表明[18],高溫脅迫下可溶性糖和淀粉含量顯著下降,并且其含量的高低與黑麥草的耐熱能力密切相關。在馬鈴薯(Solanumtuberosum)中發現,高溫脅迫下其葉片蔗糖含量升高,淀粉含量減少,而葡萄糖含量無顯著變化,這可能與蔗糖合成酶(SS)和轉化酶(INV)酶活性的提高有關[7, 35],而在甘蔗(Saccharumofficinarum)中卻發現,高溫脅迫下其葉片的糖代謝關鍵酶(SS、SPS、INV)活性都降低,蔗糖和淀粉含量減少[35]。本研究結果顯示,短期的高溫脅迫下(5 d)黑麥草葉片的SPS、SS和INV酶基因的表達量顯著升高,而在較長時間(15 d)的高溫脅迫下表達水平顯著下降,同時葉片的葡萄糖、果糖、蔗糖和淀粉等碳水化合物含量也顯著下降,表明高溫對黑麥草的碳水化合物代謝造成了影響,致使光合產物減少,代謝關鍵酶基因表達量降低,促進了呼吸作用使淀粉分解而含量下降。但是,高溫脅迫下施用外源GABA,顯著提高了黑麥草葉片SPS、SS和INV酶基因的表達水平和碳水化合物(蔗糖、果糖、葡萄糖、淀粉)含量,表明外源GABA可通過緩解高溫脅迫下光合作用的下降和提高碳水化合物代謝關鍵酶基因的表達水平和光合產物,從而提高了碳水化合物代謝水平和增強了黑麥草的耐高溫能力。
4 結論
通過對高溫條件下施用外源GABA對多年生黑麥草的生長、細胞膜透性、光合色素含量、光合及葉綠素熒光參數、碳水化合物含量及其代謝關鍵酶基因表達的分析表明,高溫脅迫下施用外源GABA,Fv/Fm、ETR、qP和ΦPSⅡ提高了13%~50%,而NPQ則降低了25%~32%,CE、Vcmax和Jmax分別提高了70%,31%和23%,降低了葉片氣孔和非氣孔限制值,增強了糖代謝關鍵酶基因表達,碳水合化合物含量提高了17%~40%,緩解了高溫對葉片PSⅡ反應中心的抑制,增強了光合能力,提高了碳水化合物含量,因而有效地提高了黑麥草的耐高溫能力。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
