馬鈴薯抗寒種質(zhì)資源分類(lèi)及其利用
2019-03-05 09:12:26彭曉君梅文祥宋波濤
中國(guó)馬鈴薯 2019年1期
關(guān)鍵詞:能力
康 黎,彭曉君,陳 琳,涂 衛(wèi),梅文祥,宋波濤
(華中農(nóng)業(yè)大學(xué)園藝林學(xué)學(xué)院/華中農(nóng)業(yè)大學(xué)園藝植物生物學(xué)教育部重點(diǎn)實(shí)驗(yàn)室/農(nóng)業(yè)農(nóng)村部馬鈴薯生物學(xué)與生物技術(shù)重點(diǎn)實(shí)驗(yàn)室,湖北 武漢 430070)
現(xiàn)在廣泛種植的馬鈴薯品種均為馬鈴薯普通栽培種(Solanum tuberosum),幾乎都不耐低溫霜凍,沒(méi)有低溫馴化能力,僅有‘鄭薯5號(hào)’等少數(shù)幾個(gè)品種具有極其微弱的抗寒能力[1],說(shuō)明栽培種內(nèi)不存在抗寒遺傳變異,這使得從普通栽培種內(nèi)部挖掘抗性材料可能性極低。目前,雖然也有一些具有抗寒能力的栽培品種被報(bào)道,如‘Alaska Frostless’[2]等,但離廣泛推廣應(yīng)用還有很長(zhǎng)一段距離,因此,加強(qiáng)對(duì)馬鈴薯抗寒種質(zhì)資源的研究,引入新的抗性種質(zhì)將有助于中國(guó)馬鈴薯產(chǎn)業(yè)的持續(xù)健康發(fā)展。
1 馬鈴薯抗寒種質(zhì)資源
馬鈴薯的抗寒性不僅取決于其種的屬性,也與低溫馴化相關(guān)。低溫馴化(也稱冷馴化)是將植物置于低溫環(huán)境中(0~12℃),植物體內(nèi)發(fā)生一系列生理生化變化,使植物產(chǎn)生適應(yīng)性反應(yīng),以提高對(duì)低溫抵抗能力的過(guò)程(Cold acclimation,CA)[3]。馬鈴薯栽培品種的遺傳背景十分的狹隘且?guī)缀鯖](méi)有抗寒能力和低溫馴化能力,但馬鈴薯野生種資源豐富,按Hawkes[4]的分類(lèi)標(biāo)準(zhǔn),可分為21個(gè)系235個(gè)種,栽培種數(shù)量為7個(gè),野生種數(shù)量為228個(gè),其中很多野生種存在耐低溫霜凍的資源,而且還有一些資源具有較強(qiáng)的低溫馴化能力。
Li[5]對(duì)60個(gè)野生種進(jìn)行梯度降溫處理,處理時(shí)間為24 h,結(jié)果表明S.acaule大部分的致死溫度可達(dá)-6.5℃,抗寒能力最強(qiáng),其次是S.boliviens、S.megistacrolobum和S.commersonii,致死溫度可低至-5.0℃,此外,S.demissum、S.torflapanum、S.sanctae-rose存在致死溫度為-4.0℃的無(wú)性系。Palta和Li[6]通過(guò)同樣的方法,也發(fā)現(xiàn)S.acaule、S.multidissectum、S.demissum、S.megistacrolobum、S.sanctae-rose、S.commersonii、S.vernei存在有無(wú)性系能夠忍耐-5℃以下低溫的植株。Chen和Li[7]將結(jié)薯馬鈴薯種的抗寒能力和冷馴化能力進(jìn)行了分類(lèi)(表1),基于有無(wú)低溫馴化能力和植株的抗寒能力將馬鈴薯的抗寒性分成5類(lèi):抗寒型馬鈴薯種并能夠低溫馴化;抗寒型馬鈴薯種不能夠低溫馴化;霜凍敏感型馬鈴薯種能夠低溫馴化;霜凍敏感型馬鈴薯種不能夠低溫馴化;霜凍敏感型馬鈴薯種。Vega和Bamberg[8]通過(guò)田間自然霜凍法,對(duì)101個(gè)種2 635份材料的抗寒性鑒定,并將抗性分為7個(gè)等級(jí)(0~6,0代表沒(méi)有受到損傷,6代表植株死亡),結(jié)果顯示S.acaule、S.albicans抗寒能力最強(qiáng),等級(jí)為0;其次是S.commersonii,等級(jí)為0.05;此外,S.demissum、S.megistacrolobum、S.paucissectum、S.multidissectum、S.sanctae-rose 5個(gè)野生種的等級(jí)介于0.5~1。Luthra等[9]于早春時(shí)候?qū)?1個(gè)種共86個(gè)材料的植株置于室外生長(zhǎng)7 d后,統(tǒng)計(jì)葉片的損傷率鑒定其抗寒能力,并劃分10個(gè)等級(jí)(1~10,1表示葉片損傷率低于10%,10表示葉片損傷率高于90%),其中有18個(gè)材料的等級(jí)為1,分布在S.goniocalyx、S.arnezii、S.demissum、S.spegazzinii中。趙喜娟[10]通過(guò)人工冷凍法對(duì)18個(gè)野生種的97份材料進(jìn)行抗寒性評(píng)價(jià),處理溫度-3.5℃,處理時(shí)間24 h,評(píng)級(jí)方法與田間自然霜凍法相同,結(jié)果表明S.acaule、S.bukasovii、S.commersonii等野生種均具有較強(qiáng)抗寒能力,評(píng)級(jí)介于0~1;而S.chomatophilum、S.demissum、S.megistacrolobum和S.acaule等野生種中也有部分植株抗寒能力較強(qiáng),其中S.acaulel中77%的植株評(píng)級(jí)小于2。在所有具有強(qiáng)抗寒能力野生種當(dāng)中,來(lái)自無(wú)莖系的野生種和S.commersonii被認(rèn)為是抗寒能力最強(qiáng)的2類(lèi)野生種,已有研究表明來(lái)自無(wú)莖系的野生種S.acaule在馴化前后分別能忍受-6和-9℃的低溫,而S.commersonii馴化前后分別能忍受-4.5和-11.5℃的低溫[7],是被研究和利用最多的耐凍性資源,以下將就這2類(lèi)野生種的利用情況進(jìn)行綜述。
2 馬鈴薯無(wú)莖系野生種分類(lèi)及其利用
2.1 無(wú)莖系野生種分類(lèi)
S.acaule、S.×aemulans、S.albicans等無(wú)莖系野生種早在20世紀(jì)50年代就已經(jīng)被發(fā)現(xiàn),但直到1990年才由Hawkes最早提出了無(wú)莖系的概念,在其分類(lèi)體系中,無(wú)莖系作為馬鈴薯組的一個(gè)系,包括有5個(gè)野生種。但早在無(wú)莖系概念還沒(méi)有提出來(lái),分類(lèi)學(xué)家對(duì)上述野生的分類(lèi)和命名產(chǎn)生了分歧,直到現(xiàn)在分歧仍然存在。總體來(lái)說(shuō),大部分分類(lèi)學(xué)家對(duì)馬鈴薯無(wú)莖系內(nèi)野生種劃分的分歧主要是區(qū)別系內(nèi)種與亞種之間的劃分,特別是S.albicans和S.×aemulans的劃分上。早期的分類(lèi)中S.albicans則是作為S.acaule的亞種S.acaule var.albicans或 S.acaule ssp.albicans存在;而S.×aemulans一直以來(lái)都是作為S.acaule的亞種而被人所熟知,但Spooner等[11]卻認(rèn)為S.acaule ssp.aemulans和S.×indounii只是野生種S.×aemulans的2種不同倍性,相關(guān)的分類(lèi)差異見(jiàn)表2[7,11-14]。
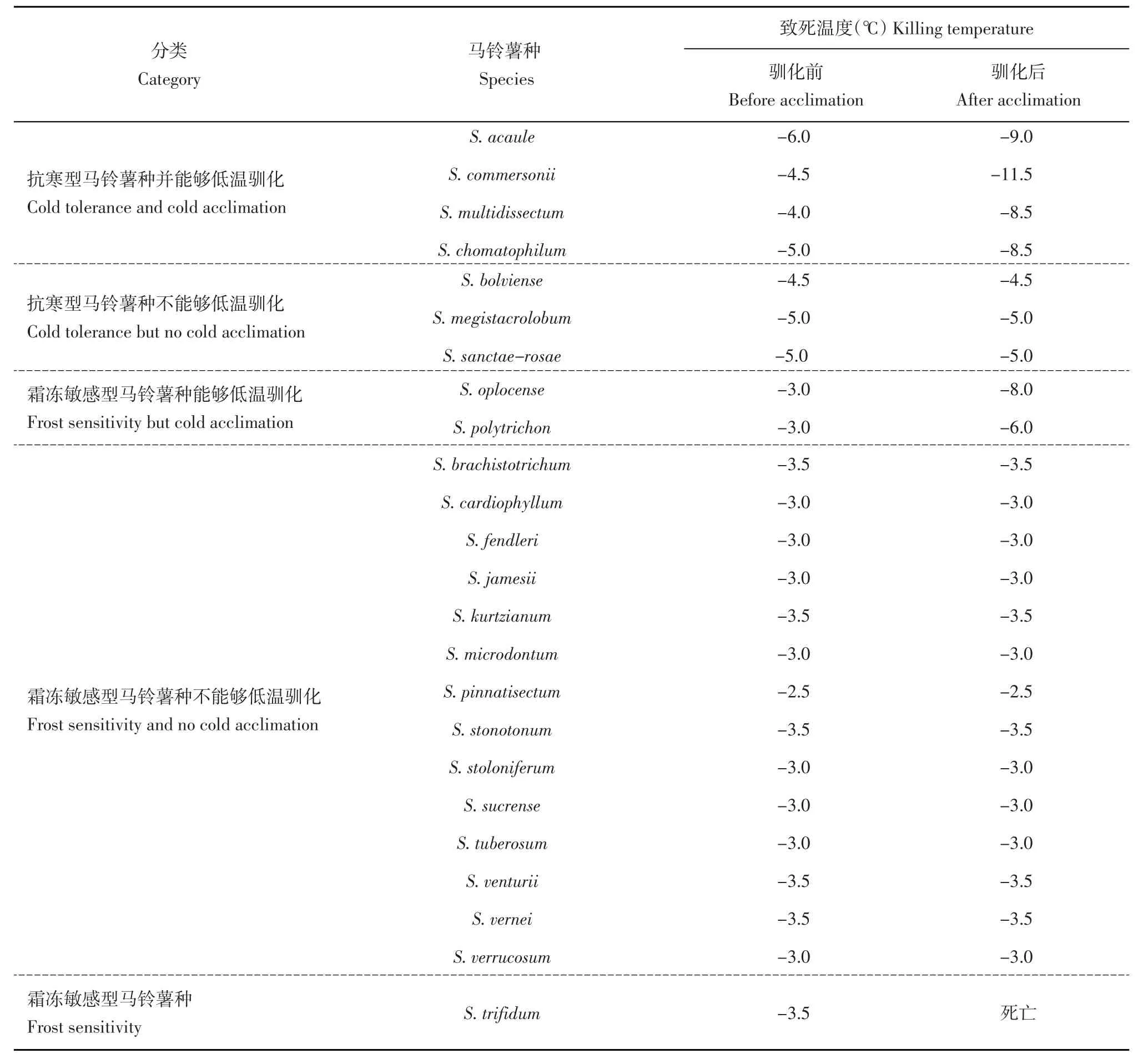
表1 馬鈴薯種的抗寒性和冷馴化能力分類(lèi)Table 1 Classification of tuber-bearing Solanum species in terms of cold tolerance and cold acclimation
2.2 無(wú)莖系野生種起源與演化
馬鈴薯無(wú)莖系野生種主要分布在南美洲安第斯山脈,分布在海拔3 500~4 500 m[14]。無(wú)莖系的起源和演化是一個(gè)非常復(fù)雜的關(guān)系,是多個(gè)野生種相互滲透的過(guò)程。
早期的研究人員通過(guò)一些形態(tài)學(xué)的觀察和雜交試驗(yàn)等研究發(fā)現(xiàn)S.acaule的植株形態(tài)與S.megistacrolobum非常相似,而葉片形態(tài)則與S.canasense非常類(lèi)似,而且S.acaule的繁殖習(xí)性也與上述2個(gè)野生種有共同之處,這表明S.acaule的形成過(guò)程中可能有S.megistacrolobum與S.canasense雜交這個(gè)環(huán)節(jié)的參與[15]。Nakagawa和Hosaka[16]通過(guò)對(duì)上百份野生種的核DNA和葉綠體DNA進(jìn)行RFLPs分析,發(fā)現(xiàn)S.acaule的葉綠體DNA在序列結(jié)構(gòu)與S.megistacrolobum、S.sanctae-rose的非常相似,都屬于C類(lèi)型;S.acaule的核DNA與S.megistacrolobum、S.sanctae-rose同樣具有極高的相似性,在進(jìn)化樹(shù)上距離非常近,都屬于Cluster 1類(lèi)型,由此推測(cè)S.acaule的起源可能和這2個(gè)野生種相關(guān)。此外,還有研究表明二倍體野生種S.boliviens可能在S.acaule形成過(guò)程中發(fā)揮了重要的作用。Goncarik[17]通過(guò)檢測(cè)數(shù)百份野生種和栽培種中葉綠體簡(jiǎn)單重復(fù)序列(cpSSR)類(lèi)型并對(duì)各個(gè)種血緣進(jìn)行分析,發(fā)現(xiàn)S.acaule和S.boliviens的cpSSR具有高度的相似性,由此推斷S.boliviens可能通過(guò)雜交的方式直接參與了S.acaule形成過(guò)程。上述研究表明,在S.acaule的形成過(guò)程中,二倍體野生種S.boliviens、S.megistacrolobum、S.canasense都可能參與其中,其中S.megistacrolobum和S.boliviens可能性最大;實(shí)際上,也有分類(lèi)學(xué)家認(rèn)為S.megistacrolobum,S.sanctae-rose和S.boliviens其實(shí)是同一個(gè)物種,即S.boliviens[11],這也進(jìn)一步證明了馬鈴薯種的形成其實(shí)是野生種在進(jìn)化過(guò)程長(zhǎng)期相互滲透的過(guò)程。

表2 馬鈴薯無(wú)莖系野生種分類(lèi)Table 2 Classification of Solanum acaule Juz
野生種S.acaule也與無(wú)莖系其他種的形成有著密切的聯(lián)系。S.×aemulans可能是S.acaule ssp.acaule和二倍體野生種S.megistacrolobum的天然雜交后代[18],且因?yàn)镾.megistacrolobum能產(chǎn)生2n配子,而導(dǎo)致了S.×aemulans存在三倍體和四倍體2種不同的倍性;S.albicans則有可能是S.acaule ssp.punae和一個(gè)與無(wú)莖系親緣關(guān)系較遠(yuǎn)的二倍體雜交后經(jīng)過(guò)天然加倍而形成的[4]。
除了參與無(wú)莖系其他野生種的形成,S.acaule還可能參與了其他野生種和原始栽培種的形成。S.×brucheri可能是S.acaule ssp.acaule和二倍體野生種S.infundibuliforme的天然雜交后代[19]。原始栽培種S.×juzepczukii和S.×curtilobum可能也會(huì)有S.acaule的血緣[20-22],具體演化的過(guò)程為:S.×juzepczukii為S.acaule與S.stenomum的天然雜交后代,S.×curtilobum則為S.×juzepczukii的2n配子與S.tuberosum ssp.andigenum的天然雜交后代(圖1)。
2.3 無(wú)莖系野生種在抗寒育種中的應(yīng)用
馬鈴薯無(wú)莖系野生種中擁有豐富的耐低溫霜凍材料,是馬鈴薯抗寒育種的重要種質(zhì)資源,S.acaule則是無(wú)莖系中利用最為普遍的野生種,是目前已知抗寒能力最強(qiáng)的野生種之一。英國(guó)早在20世紀(jì)40年代就試圖利用S.acaule來(lái)培育抗寒馬鈴薯品種,但收效甚微。從目前已有的報(bào)道來(lái)看,利用S.acaule選育出來(lái)的并且具有抗寒能力品種只有‘Alaska Frostless’[2],該品種的選育過(guò)程中有一個(gè)加倍后的S.acaule無(wú)性系參與了其中,其經(jīng)過(guò)馴化后LT50可以達(dá)到-4.6℃。除了‘Alaska Frostless’,其余的均為一些可用于抗寒育種的中間材料。Estrada[23]用四倍體的S.acaule和二倍體的S.phureja以及四倍體的普通栽培種依次雜交后成功獲得了雜種后代,有27%的后代具有較強(qiáng)的抗寒能力;稍晚一些,Estrada[24]又直接從S.acaule和S.phureja的雜種后代中鑒定了一個(gè)四倍體acaphu,acaphu可直接和四倍體栽培種雜交產(chǎn)生四倍體的后代,且后代中某些植株能夠忍耐-5℃低溫2 h不死;Bamberg等[25]通過(guò)用加倍后S.commersonii作為橋梁種,成功獲得了ACL-CMM-TBR 3者的種間雜種,該種間雜種具有一定的抗寒能力和馴化能力,可用于后續(xù)的抗寒育種。
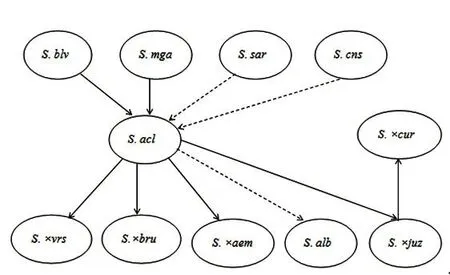
圖1 無(wú)莖系的起源與演化Figure 1 Origin and evolution of acaule Juz
2.4 馬鈴薯野生種在抗病育種中的應(yīng)用
除了抗寒能力之外,馬鈴薯無(wú)莖系野生種中還擁有豐富的抗馬鈴薯病毒材料,是馬鈴薯抗病毒育種的重要種質(zhì)資源,特別是野生種S.acaule。據(jù)不完全統(tǒng)計(jì),野生種S.acaule中存在有抗癌腫病,抗馬鈴薯X病毒(Potato virus X,PVX)、馬鈴薯卷葉病毒(Potato leafroll virus,PLRV)、馬鈴薯紡錘塊莖類(lèi)病毒(Potato spindle tuber viroid,PSTVd)的材料[26,27]。
野生種S.acaule在馬鈴薯病毒抗性育種中的利用已經(jīng)非常的廣泛,已經(jīng)獲得了非常多育種中間材料和一些品種。有研究用7XY.1、Bolona等四倍體的普通栽培種和S.acaule雜交,授粉第2 d再用S.phureja(IVP35)進(jìn)行二次授粉,待第一次授粉后14~27 d采用胚搶救(Embryo rescue),成功地獲得了3個(gè)可育的四倍體雜種,這3個(gè)雜種均檢測(cè)出了PSTVd抗性,而且在隨后的研究中表明,這3個(gè)四倍體均具有優(yōu)良結(jié)薯表型[28,29];Watanabe等[30]通過(guò)染色體加倍和多次回交的方式,獲得了含有S.acaule血緣的四倍體雜種,這些雜種中有部分植株檢測(cè)到了PLRV和PSTVd抗性;還有研究人員通過(guò)將S.acaule和普通栽培種進(jìn)行體細(xì)胞融合,獲得一批倍性不一的體細(xì)胞雜種,這些體細(xì)胞雜種中均存在有PVX極端抗性植株,且有些植株結(jié)薯性狀非常優(yōu)良,可作為育種中間材料[31-33]。此外,波蘭科學(xué)家利用染色加倍將S.acaule的一個(gè)基因型變成同源八倍體后與普通栽培種雜交后進(jìn)行多次回交,成功的獲得回交雜種MPI44.106/10,該雜種被用于抗PVX的育種親本,并用其選育出了多個(gè)抗PVX的品種,典型的如‘Barbara’[34];中國(guó)黑龍江省農(nóng)業(yè)科學(xué)院馬鈴薯研究所同樣通過(guò)加倍的方法將野生種S.acaule的PVX抗性導(dǎo)入到了普通栽培種當(dāng)中,并通過(guò)3~4代輪回選擇后成功獲得了多份抗PVX的育種親本[35]。
3 S.commersonii及其在抗寒育種中利用
3.1 野生種S.commersonii概述
S.commersonii隸屬馬鈴薯組孔目松系,為二倍體野生種,EBN(Endosperm balance number)=1,主要分布在阿根廷,烏拉圭,巴西等國(guó)家,生長(zhǎng)于安第斯山脈海拔400 m左右的區(qū)域。在Hawkes[4]的分類(lèi)體系中,S.commersonii分為2個(gè)亞種,分別是S.commersonii ssp.commersonii和S.commersonii ssp.malmeanum;但在Spooner等[11]的分類(lèi)體系中,S.commersonii ssp.malmeanum不再是S.commersonii的一個(gè)亞種,而是作為一個(gè)獨(dú)立的野生種,命名為S.malmeanum。
S.commersonii種內(nèi)具有豐富的抗性資源,據(jù)不完全統(tǒng)計(jì),S.commersonii種內(nèi)不僅存在有馬鈴薯瘡痂病、PVX、科羅拉多甲蟲(chóng)等病蟲(chóng)害的抗性,對(duì)某些不良環(huán)境也具有極強(qiáng)的耐性,其中最為典型的就是耐低溫霜凍[11]。研究表明S.commersonii具有極強(qiáng)的低溫馴化能力,馴化后其LT50達(dá)-10℃以下,低溫馴化能力高于7℃以上[7],是已知低溫馴化能力最強(qiáng)的馬鈴薯野生種。
3.2 S.commersonii在抗寒育種中的應(yīng)用
目前在利用S.commersonii進(jìn)行抗寒育種方面,已經(jīng)取得了一些初步的成果。Caidi等[36]利用二倍體的S.commersonii和四倍體的栽培種進(jìn)行體細(xì)胞融合后得到了雄性不育雜種后代,70%是四倍體,其中編號(hào)為cmm1的種間雜種的馴化能力達(dá)到了6℃;Nyman和Waara[37]利用二倍體栽培種和S.commersonii體細(xì)胞融合得到雜種,經(jīng)檢測(cè),某些種間雜種馴化后能夠忍受-9℃左右的低溫。Chen等[38]同樣利用S.commersonii和二倍體栽培種體細(xì)胞融合后獲得了一個(gè)四倍體的雜種后代,該四倍體和栽培種‘Wis AG 231’回交后代中,有21%的后代低溫馴化能力得到了顯著提升(即馴化能力大于或等于2.5℃),其中某些植株的結(jié)薯產(chǎn)量甚至超過(guò)了回交親本。Bastia等[39]同樣利用體細(xì)胞融合的方法獲得了不同倍性的雜種,在四倍體雜種中,一些雜種馴化后LT50低于-5℃,而在八倍體中,有些雜種馴化后LT50甚至可達(dá)-9℃。Sepp?nen等[40]也是通過(guò)原生質(zhì)體融合的手段獲得了S.commersonii和栽培種可育的體細(xì)胞雜種,其中一個(gè)編號(hào)為SH9A的雜種經(jīng)過(guò)低溫馴化其LT50為-5.2℃,且其自交后代中同樣能檢測(cè)抗低溫植株存在,值得一提的,SH9A還具有對(duì)PVX的極端抗性。上述這些都是通過(guò)體細(xì)胞融合的方式得到雜種后代,而Carputo等[41,42]則是通過(guò)雜交的手段,成功獲得了S.commersonii和四倍體栽培種‘UP88-P5’的F1代(三倍體),這些后代經(jīng)過(guò)馴化后能夠忍耐-6.2℃的低溫2 h,其中編號(hào)為B10的植株可以忍耐-7.5℃的低溫,在該三倍體與四倍體栽培種‘Tollocan’的BC1和BC2中,均有抗寒能力較強(qiáng)的植株存在(BC1中有植株的馴化抗寒性為-5.5℃,BC2中植株的馴化抗寒性也可達(dá)-4.8℃)。
4 馬鈴薯抗寒育種展望
馬鈴薯野生種擁有豐富的抗寒種質(zhì)資源,但目前對(duì)抗性資源的利用尚未獲得理想的效果,至今還沒(méi)有能夠被推廣種植的抗寒品種。目前馬鈴薯育種技術(shù)主要有傳統(tǒng)雜種育種、倍性育種、體細(xì)胞融合、橋梁育種、分子標(biāo)記輔助選擇育種以及轉(zhuǎn)基因技術(shù)等方法。傳統(tǒng)雜交育種是目前最主要的育種手段之一,但在抗寒育種上收效甚微,一方面,由于EBN不同的原因,大部分馬鈴薯抗寒資源難以直接和栽培種進(jìn)行雜交;另一方面,大部分野生種抗寒能力是一個(gè)由多基因控制的數(shù)量性狀,在多次回交的過(guò)程中容易導(dǎo)致抗性衰減。倍性育種、體細(xì)胞融合、橋梁育種雖然能夠克服遠(yuǎn)緣雜交不親和性,創(chuàng)制新的抗寒種質(zhì)資源,但在后續(xù)的過(guò)程仍需要進(jìn)行常規(guī)的雜交。在轉(zhuǎn)基因技術(shù)上,由于目前對(duì)于抗寒能力的遺傳規(guī)律和分子機(jī)制的解析尚不夠完全,所取得的成果也比較少,民眾對(duì)轉(zhuǎn)基因技術(shù)也還存在較大的爭(zhēng)議。此外,在分子標(biāo)記開(kāi)發(fā)上,目前極少有能應(yīng)用于抗寒育種的分子標(biāo)記被報(bào)道。因此,加強(qiáng)對(duì)馬鈴薯抗寒遺傳規(guī)律和分子機(jī)制的研究,開(kāi)發(fā)更多能應(yīng)用于抗寒育種的分子標(biāo)記,將極大的推動(dòng)馬鈴薯抗寒育種的進(jìn)程。
猜你喜歡
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動(dòng)漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學(xué)版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國(guó)生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(shè)(2018年6期)2018-08-16 07:23:10
新高考(英語(yǔ)進(jìn)階)(2018年1期)2018-04-18 14:00:11
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14
