高氮和低氮條件下玉米穗位葉持綠性狀的QTL定位
2019-03-08 03:24:58李東亞湯繼華譚金芳韓燕來
植物營養與肥料學報 2019年1期
關鍵詞:檢測
李東亞,王 祎,2,*,湯繼華,許 恒,譚金芳,2,韓燕來,2,*
(1 河南農業大學資源與環境學院,河南鄭州 450002;2 河南省糧食作物協同創新中心,河南鄭州 450002;3 省部共建小麥玉米國家重點實驗室,河南鄭州 450002)
玉米作為三大重要糧食作物之一,被廣泛應用于工業原料、食品加工等領域。2012年我國玉米總產超過稻谷,成為總產和種植面積均第一的糧食作物[1]。前人研究認為,我國人均玉米占有率越高,其生活水平及畜牧業發展水平越高[2]。我國玉米高產主要依賴于氮肥的大量投入,研究表明,1985至1994十年間,化肥在玉米增產中起到51.5%的作用[3],并且數據顯示2008—2009年玉米的氮肥使用比例占我國主要農作物氮肥消費總量的19%,遠高于水稻、小麥等作物的氮肥用量[4]。但是我國玉米的氮肥利用率僅有26.1%,遠低于國際水平,且低于我國水稻和小麥的氮肥利用率[5]。氮肥長期大量施用已引發土壤酸化、水體富營養化、溫室效應等一系列環境問題[6-8]。因此,提高玉米氮肥利用率、培育氮高效品種,對實現我國玉米高產高效和減輕過量施氮對環境污染的壓力具有重要意義[9]。
葉片黃化是植株氮素缺乏的主要癥狀,研究表明葉片葉綠素含量與葉片氮含量顯著相關,可用來反映葉片氮營養水平[10]。葉片壽命與氮素水平具有密切關系,葉片光合作用的提高或降低亦與氮素的輸出有關。玉米吐絲后的光合作用對產量的形成具有決定作用,其中穗位葉對籽粒碳水化合物的供應尤為重要[9]。氮素供應不足加速穗位葉黃化,降低光合速率,最終導致產量下降[11]。Thomas等[12]研究表明,持綠性是指植物在進入成熟期后葉片衰老延緩而保持綠色的特性,被廣泛認定為植物的理想農藝性狀。持綠型植株不僅持綠時間長,而且葉片顏色也相對較深。有研究發現,在植物成熟后期,葉片持綠每延長1天,籽粒產量可提高2%[13],并且持綠性與植物保持較高光合能力、增加產量、提高品質、增強抗逆性等密切相關[14-17]。籽粒灌漿期氮素供求關系的平衡決定著植株葉片持綠性[18],從而影響植株氮效率和產量[19-21]。當前可用于評價玉米持綠性的指標包括綠葉面積持續期、花后絕對綠葉面積、相對綠葉面積、葉綠素含量等[22]。葉綠素含量作為與植株氮含量緊密關聯的次級指標能夠很好地反映葉片的氮含量,并與葉片光合作用密切相關,且具有相對較高的遺傳穩定性,可用于持綠QTL定位分析[23-25]。
前人在不同氮條件下,利用單片段代換系對玉米穗部性狀[26]、開花期相關性狀[27]、苗期根系形態及氮吸收量[28]等方面的QTL定位進行了初步研究,但有關玉米穗位葉持綠相關性狀的QTL定位研究相對報道較少。鄭洪建[29]定位了80個與持綠相關的玉米QTL,其中包含31個主效QTL,但其研究是在正常供氮條件下進行的,有關低氮條件下玉米持綠相關QTL的定位特征尚不清楚。本研究利用大田全生育期條件下低氮脅迫不敏感自交系許178和低氮脅迫敏感材料綜3構建的單片段代換系作為研究對象[30-31],對兩年大田高、低氮條件下玉米穗位葉的持綠性狀進行了QTL定位分析,旨在為玉米持綠相關基因的精細定位奠定基礎。
1 材料與方法
1.1 試驗材料
本研究所用的玉米單片段代換系由河南農業大學湯繼華教授提供。此群體綜3 (氮敏感材料,SPAD值較許178低,表1) 為供體親本,許178 (氮不敏感材料) 為受體親本,經過4個世代回交和3個世代自交,結合SSR分子標記輔助選擇的方法構建而成[30-31]。
1.2 田間試驗設計
2013年和2014年的6至10月,在河南新鄭“華北小麥玉米輪作營養與施肥觀測試驗站”對單片段代換系群體進行田間試驗。土壤基本理化性狀如下:全氮0.940 g/kg、有機質10.4 g/ kg、堿解氮28.4 mg/kg、有效磷8.17 mg/kg、速效鉀63.0 mg/kg、pH值7.82。試驗采取完全隨機區組設計,設置高氮 (N 240 kg/hm2) 和低氮 (N 75 kg/hm2) 兩個處理,每個處理重復三次。供試肥料為尿素和磷酸二氫銨,兩氮素處理的磷、鉀肥用量相同 (P2O5192 kg/hm2,K2O 60 kg/hm2)。灌溉、除草和防蟲等田間管理均與大田管理一致。前茬作物小麥收獲后,于6月10號進行玉米播種,播種時不施肥。拔節期(7月2日) 和大喇叭口期 (7月30日) 采用穴施方式追肥。
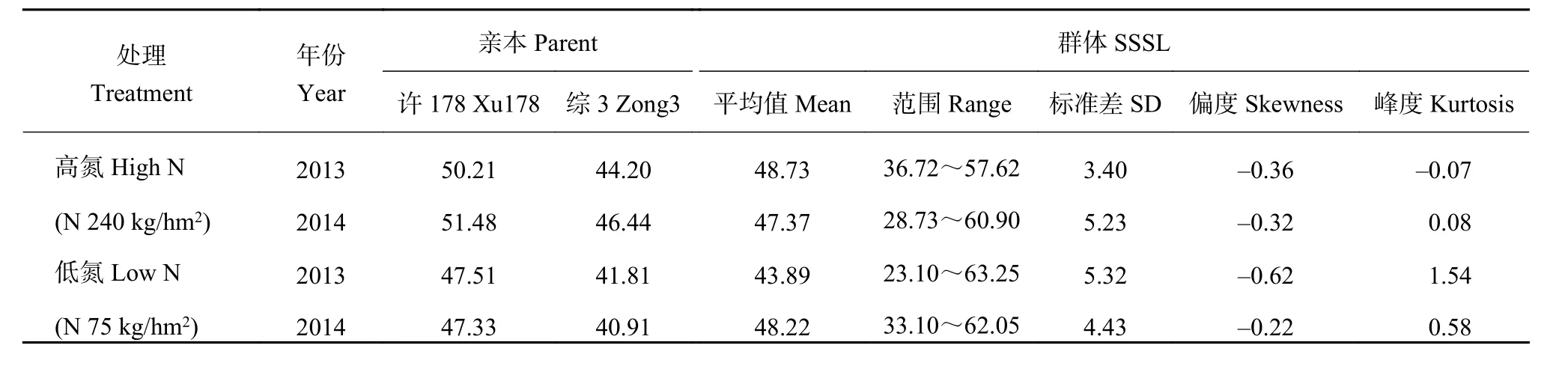
表1 群體SPAD值Table 1 SSSL SPAD values
1.3 測定指標
玉米吐絲后第10天,使用SPAD 502葉綠素含量測定儀測定穗位葉葉綠素含量。測定時選取穗位葉中間部位約10 cm的區域連續測定5個點 (避開主脈),取平均值作為該葉片的SPAD值。每個小區選取6株長勢一致的植株進行測定。
1.4 數據分析與QTL定位
采用SPSS 23.0和Excel 2016軟件進行數據分析,若群體單株表型值與許178在P < 0.01水平上差異顯著,則認為該單片段代換系的代換片段上存在一個持綠QTL,并同時估算各個QTL的加性效應值及貢獻率[32]。
加性效應值 = (單片段代換系的表型值 - 許178表型值)/2;
加性效應貢獻率 = 加性效應貢獻值/許178表型值 × 100%。
1.5 QTL命名
根據Mccouch等[33]制定的原則對QTL命名,q為QTL的縮寫,hn表示高氮 (high nitrogen),ln表示低氮 (low nitrogen),SG代表持綠 (stay-green),第一個數字為QTL所在染色體的編號,最后一個字母為此性狀在該染色體上的QTL序號。
2 結果與分析
2.1 不同氮水平下親本和SSSL群體穗位葉持綠表型分析
由表1可知,高氮條件下,2013年親本許178和綜3的表型值分別是50.21和44.20,許178較綜3高13.60%;2014年兩親本的表型值分別是51.48和46.44,許178比綜3高10.85%。低氮條件下,2013年親本許178和綜3的表型值分別是47.51和41.81,許178比綜3高13.63%;2014年兩親本的表型值分別是47.33和40.91,許178較綜3高15.69%。綜上分析,兩親本在不同氮條件下穗位葉的持綠性具有顯著性差異。高氮條件下,2013年不同SSSLs的SPAD值變異范圍是36.72~57.62,平均值為48.73 ± 3.40;在2014年其變異范圍為28.73~60.90,平均值是47.37 ± 5.23。低氮條件下,不同SSSLs在2013和2014年的SPAD的變化范圍分別是23.10~63.25、33.10~62.05,平均值分別是43.89 ±5.32、48.22 ± 4.43。不同SSSLs的SPAD值在不同處理條件下均具有較大變異,為QTL的定位提供豐富的表型數據。
2.2 不同氮水平下玉米穗位葉持綠QTL分析
通過QTL代換作圖,對SSSL群體不同氮水平下檢測到的玉米穗位葉持綠QTL進行分析。兩個氮水平下共檢測到53個穗位葉持綠QTL,且在10條染色體上均有分布。其中2013年,兩個氮水平下共檢測到22個QTL;2014年,兩個氮水平下共檢測到31個QTL。單個QTL的加性效應貢獻率為-2.45%~-22.65% (表 2)。
高氮 (HN) 條件下,在P < 0.01時,兩年共檢測到29個玉米穗位葉持綠QTL,10條染色體均有分布,其中僅2013年檢測到的QTL qhnSG1e表現為正
向加性效應 (貢獻率為4.02%),其余QTL均表現為負向加性效應 (貢獻率為-2.45%~-13.77%) (表2)。HN條件下,有6個QTL兩年被重復檢測到,分別是qhnS1d,加性效應貢獻率分別是-4.68%和-13.77%;qhnSG2a,加性效應貢獻率分別是-4.80%和-11.61%;qhnSG3a,加性效應貢獻率分別是-7.01%和-6.92%;qhnSG4a,加性效應貢獻率分別是-7.40%和-2.45%;qhnSG8b,加性效應貢獻率分別是-4.47%和-9.17%;qhnSG10c,加性效應貢獻率分別是-9.46%和-5.05%。其余23個QTL僅在2013或2014年被檢測到。
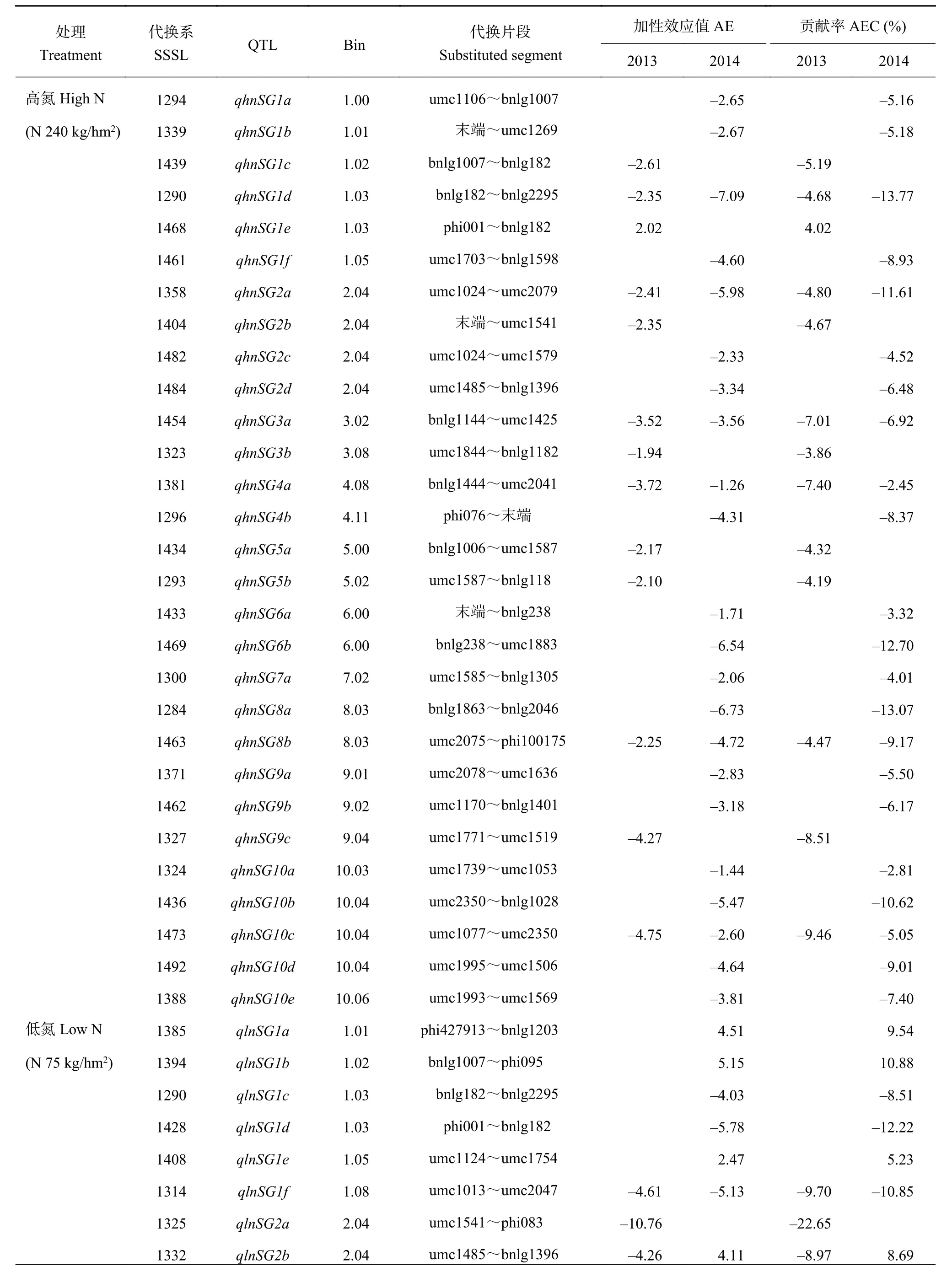
表2 不同氮水平下SSSL群體穗位葉持綠QTL及其效應值Table 2 QTL location of ear leaf stay-green in maize in the SSSL population under different nitrogen conditions
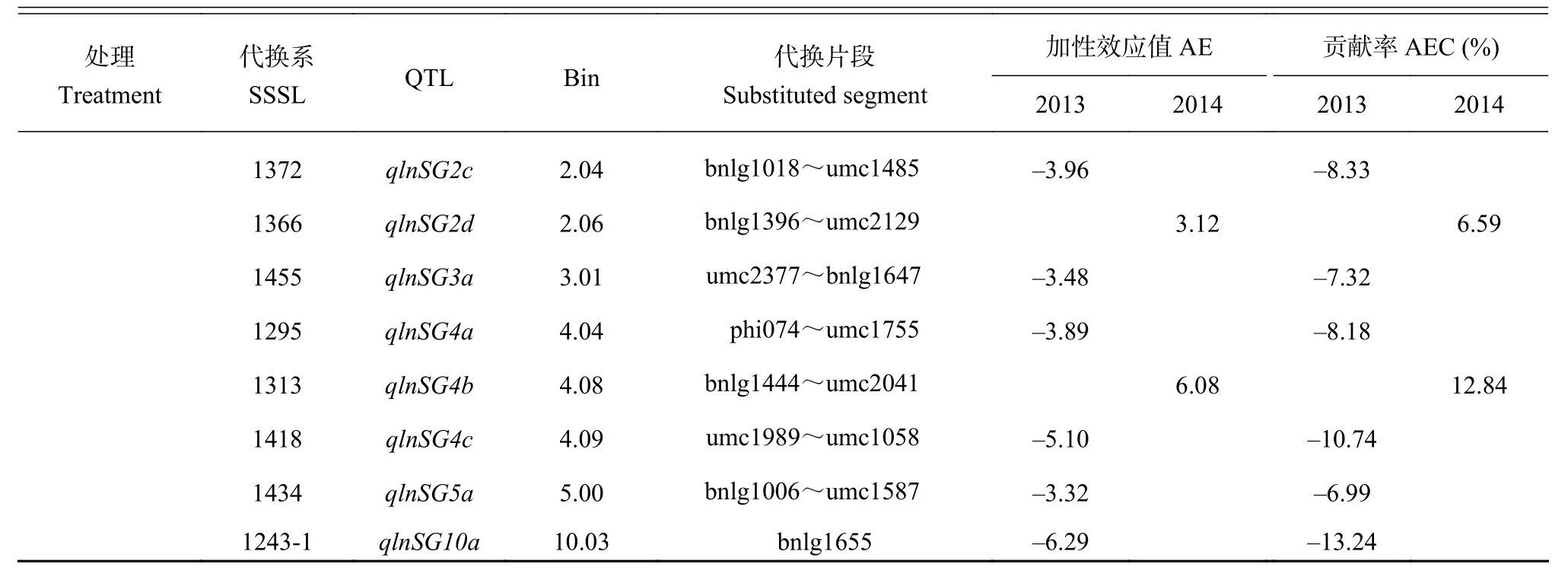
續表 2 Table 2 continued
低氮 (LN) 條件下,兩年共檢測到16個QTL (P <0.01),分別分布在第1、2、3、4、5和10號染色體上,其中2013年檢測到的9個QTL均表現為負加性效應 (貢獻率為-6.99%~-22.65%),2014年檢測到的QTL有6個正加性效應 (貢獻率為5.23%~12.84%),3個負加性效應 (貢獻率分別為-8.51%、-12.22%和-10.85%)。有2個QTL在兩年被重復檢測到,分別是qlnSG1f和qlnSG2b。qlnSG1f位于Bin1.08區域,代換片段為umc1013~umc2047,兩年加性效應貢獻率分別是-9.70%和-10.85%;qlnSG2b位于Bin2.04區域,代換片段為umc1485~bnlg1396,2013年為負加性效應 (貢獻率為-8.97%),2014年為正加性效應 (貢獻率為8.69%)。其余14個QTL僅在2013或2014年被檢測到。
2.3 QTL在染色體上的分布
圖1所示為兩年試驗中兩個氮水平條件下檢測到的QTL在染色體上的分布狀況。結果表明,定位到的穗位葉持綠QTL在玉米10條染色體上均有分布。其中第1染色體上檢測到的QTL數目最多(14個),而第7染色體上僅檢測到1個QTL,數目最少。通過分析QTL在染色體上的分布發現,多數定位到的持綠QTL在高、低氮條件下同時檢測到,表明該持綠QTL為非特異性QTL。另外,有些染色體片段僅在高氮條件下檢測到QTL,稱為高氮特異QTL,例如第1染色體的phi097~umc1269區間、第2染色體的umc1024~umc1579區間、第3染色體的umc1844~pnlg1182區間,以及第6、第8、第9和第10染色體上均有高氮特異的持綠QTL檢測到。這其中位于第8染色體的umc2075~phi100175(代換系1463的代換片段) 和第10染色體的umc1077~umc2350 (代換系1473的代換片段) 區域,在兩年試驗中可以重復檢測到。相較于高氮特異的持綠QTL,低氮特異的持綠QTL數目較少,僅分布在第1染色體的umc1013~umc2047區間和第4染色體的phi074~umc1755區間內。其中第1染色體的umc1013~umc2047 (代換系1314的代換片段) 區域檢測到的低氮特異持綠QTL在兩年的大田試驗中重復出現,表明該區段可能僅控制低氮條件下玉米穗位葉持綠,有利于我們挖掘低氮條件下特異的QTL。
3 討論
研究表明,植株氮素營養與持綠之間存在既密切又復雜的關系[19-21,34]。因此,克隆直接與氮效率緊密相關的葉片持綠基因,對于深入分析葉片持綠與氮效率之間的關系具有重要意義。方永豐等[35]通過對劉宗華等[36]、王愛玉等[37]、Zheng等[38]研究中定位到的持綠QTL進行Meta分析,發現5個玉米持綠“一致性”QTL區間,并在這些QTL區間內初步確定了8個與玉米持綠相關的候選基因。本研究在高、低兩種供氮條件下,通過兩年大田試驗,利用172個單片段代換系材料,分析了玉米吐絲后第10天穗位葉SPAD值,共定位到53個持綠QTL,其中低氮條件下定位到的特異染色體區段umc1013~umc2047和高氮條件下定位到的特異染色體區段umc1077~umc2350,與方永豐等[35]通過Meta分析檢測到的持綠QTL區間有重疊。馮萬軍等[39]對玉米谷氨酰胺合成酶 (GS) 基因家族進行定位分析發現,ZmGS1-2 (GRMZM2G024104) 基因位于第1染色體,該位置位于本研究中代換系材料1314 (Bin1.08,umc1013~umc2047) 的代換區間內。GS根據亞細胞定位可分為胞質型GS1和質體型GS2,高等植物GS1主要參與種子萌發時氮源的轉運和葉片衰老時氮源的轉移再利用,說明該染色體片段內的基因與葉片衰老和氮代謝關系密切。Yang等[40]利用鄭58和B73構建的165個F3:4自交系群體材料,通過三個表型 (葉綠素含量、光合效率和持綠葉面積) 對玉米持綠性進行定位,共檢測到23個與持綠相關的QTL,其中第8染色體上的區段phi100175~umc1858在三個表型中均被檢測到,該區段所在位置與本研究中檢測到的高氮特異持綠QTL區間 (Bin8.03,umc2075~phi100175) 部分重疊。這些結果表明本研究中重復定位到的QTL區間內可能存在與氮效率關系密切的玉米穗位葉持綠主效QTL。另外,筆者所在課題組前期對本研究所用的代換系群體苗期氮效率相關QTL進行了定位[28],研究發現在1314株系的代換片段 (umc1013~umc2047) 上定位到低氮條件下植株生物量QTL和氮含量QTL,該片段在本研究中定位到低氮特異的持綠QTL;代換系1473的代換片段 (umc1077~umc2350) 在前期的研究中定位到高氮條件下根系形態及氮積累量QTL,而該片段在本研究中定位到高氮特異的持綠性QTL。這一結果表明,玉米穗位葉的持綠性與氮效率之間存在密切關系。后期對高、低氮特異QTL進行精細定位與分析,有助于解析玉米氮效率與持綠之間的復雜關系,對于培育高、低氮條件下持綠玉米品種具有重要的參考意義。
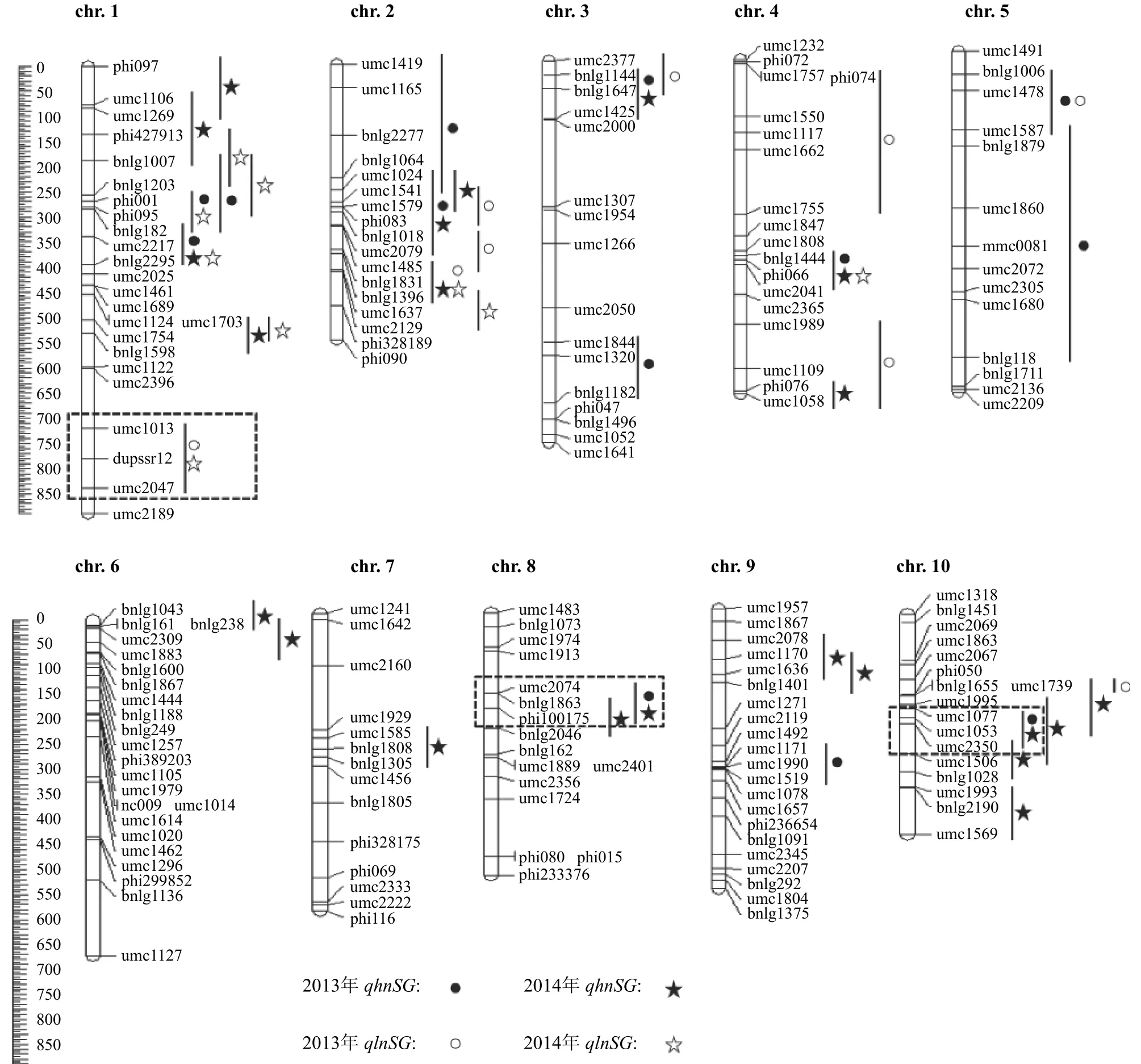
圖1 不同氮水平下玉米穗位葉持綠QTL在染色體上的分布Fig. 1 Position of the QTLs for ear leaf stay-green under low and high N conditions in maize
4 結論
本研究分別在umc1077~umc2350 (Bin10.04,代換系1473) 和umc1013~umc2047 (Bin1.08,代換系1314) 區段內檢測到高、低氮特異的持綠QTL,且該區段與前人定位到的氮效率相關QTL區間重疊,說明這兩個持綠QTL與玉米氮效率關系密切,后期對該區間進行精細定位,獲得候選基因,有益于探討玉米氮效率與持綠性之間的復雜關系。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
