15個飼用燕麥品種的遺傳變異及親緣關系分析
2019-03-21 07:18:26,,,,
種子 2019年2期
,,,,
(1.西南民族大學生命科學與技術學院,四川 成都 610041;2.西南民族大學青藏高原研究院,四川 成都 610041;3.四川農業大學草業科學系,四川 成都 610066)
燕麥是禾本科燕麥屬(AvenaL.)的草本植物,是優良的一年生糧飼兼用作物。燕麥廣泛分布于全世界五大洲76個國家,主要種植于亞洲、歐洲、北美洲北緯40°以北地區[1]。目前在生產上廣泛應用的燕麥包括普通栽培燕麥(皮燕麥)(A.sativa)和大粒裸燕麥(A.nuda)[2],其中皮燕麥的種植面積最大,主要為飼用。我國主要種植裸燕麥,主要食用,少數飼用。燕麥的飼用價值較高,其籽實、稃殼、莖葉等均是各種家畜的優良飼料。燕麥喜冷涼,耐貧瘠,可在高海拔和高緯度地區種植,已經成為我國青藏高原高寒牧區高產人工草地建設的當家草種,對緩解高寒地區家畜冬季缺草的問題具有重要的作用[3]。
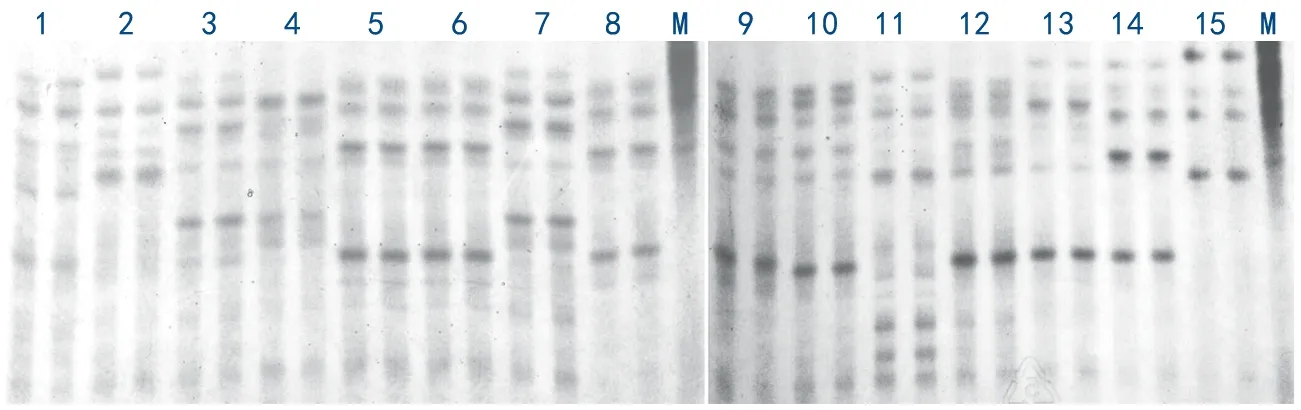
圖1 供試燕麥品種醇溶蛋白電泳圖譜
目前在青藏高原地區主要種植的燕麥品種包括青引1號、青引2號等國產燕麥品種及部分國外進口燕麥品種,而目前對這些燕麥品種間的遺傳變異等方面的研究還較少,有必要對不同燕麥品種間的遺傳差異及親緣關系進行分析,這對燕麥品種的鑒定及雜交種質的創制等方面具有重要的意義[3]。此外,在高寒地區牧草種子生產及推廣應用過程中,種子的混雜情況較為嚴重,所以對不同的牧草品種建立相關的純度及真實性的鑒定技術,對于牧草品種的推廣應用具有重要的作用。目前在牧草中包括多花黑麥草、鴨茅等都建立有相關品種的分子標記指紋圖譜[4-5],常用的方法技術包括醇溶蛋白電泳、SSR、SNP等DNA分子標記電泳等[4-7]。種子醇溶蛋白電泳、SSR、AFLP等分子標記均在燕麥種質評價、遺傳多樣性研究等方面成功運用[8-10]。本研究采用種子醇溶蛋白電泳對目前高寒地區常用的燕麥品種開展其遺傳變異及親緣關系的分析,為燕麥品種的鑒定以及資源的保護、利用等提供參考。
1 材料與方法
1.1 供試材料
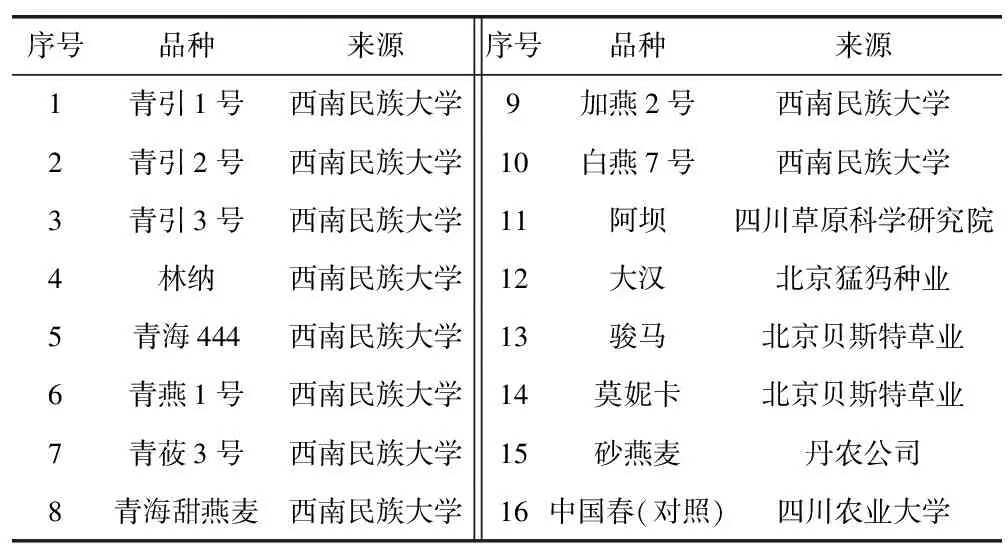
表1 供試材料
供試材料為15個飼用燕麥品種,其中青引3號、青莜3號為裸燕麥(A.nuda),砂燕麥(A.strigosa)為二倍體栽培燕麥,其余12個燕麥品種均為普通栽培燕麥(A.sativa),大多數品種為目前青藏高原高寒地區主要種植的品種(詳見表1),以小麥品種中國春作為醇溶蛋白圖譜的對照。
1.2 醇溶蛋白電泳分析
1.2.1 樣品提取
每個燕麥品種隨機選取5粒種子,設置2個重復。將皮燕麥種子去皮研磨成粉末并稱重,將其轉入1.5 mL 離心管,按照1 mg加5μL 的比例加入醇溶蛋白提取液(2-氯乙醇25%+甲基綠0.05%),室溫浸提過夜,使用前10 000 r/min離心15 min。為了鑒定品種的真實性,選擇青燕1號為檢測對象,隨機選取50粒種子,醇溶蛋白提取步驟同上。
1.2.2 A-PAGE分析
采用國際種子檢驗協會ISTA(1986)頒布的酸性聚丙烯酰胺凝膠電泳(A-PAGE,pH 3.2)標準程序[11]。采用DYCZ-24 F型電泳槽(北京六一儀器廠)進行電泳,恒壓500 V,恒溫10~15 ℃,電泳1.5 h。電泳結束后用10%的三氯乙酸固定30 min,后加入1%的考馬斯亮藍R-250染液進行染色過夜,7%的醋酸溶液褪色,數碼相機采集照片保存。
1.3 數據分析
對獲得的電泳條帶按有帶記為1,無帶記為0統計,將醇溶蛋白條帶轉換成0,1矩陣。利用軟件NTSYS-pc 2.10計算Dice遺傳相似系數(GS),并基于該遺傳相似系數進行非加權成對算術平均法(UPGMA)聚類分析和主成分分析(PCA)[12]。
2 結果與分析
2.1 醇溶蛋白多態性
15份材料共擴增出18條醇溶蛋白條帶,均為多態性條帶,醇溶蛋白位點多態率為100%(如圖1)。每個品種擴增的醇溶蛋白條帶數為4~10條,燕麥的醇溶蛋白條帶主要以遷移率較大的為主,分子量較小的位點,主要分布于α和β區域。每份品種隨機挑選5粒種子,并設置2個重復,2個重復的電泳條帶均保持一致,這也表明醇溶蛋白電泳在燕麥中具有較好的重復性(如圖1)。本研究中醇溶蛋白電泳能夠將供試的13個品種分開,僅有青海444和青燕1號2個品種的帶型完全一致,說明醇溶蛋白對于燕麥品種具有較好的鑒別能力。此外,為了鑒定燕麥種子的真實性,隨機選擇青燕1號燕麥的50粒種子進行電泳,從電泳圖譜來看完全一致,表明該品種具有較高的純度(如圖2),同時說明醇溶蛋白電泳是燕麥種子真實性鑒定的一種有效方法。
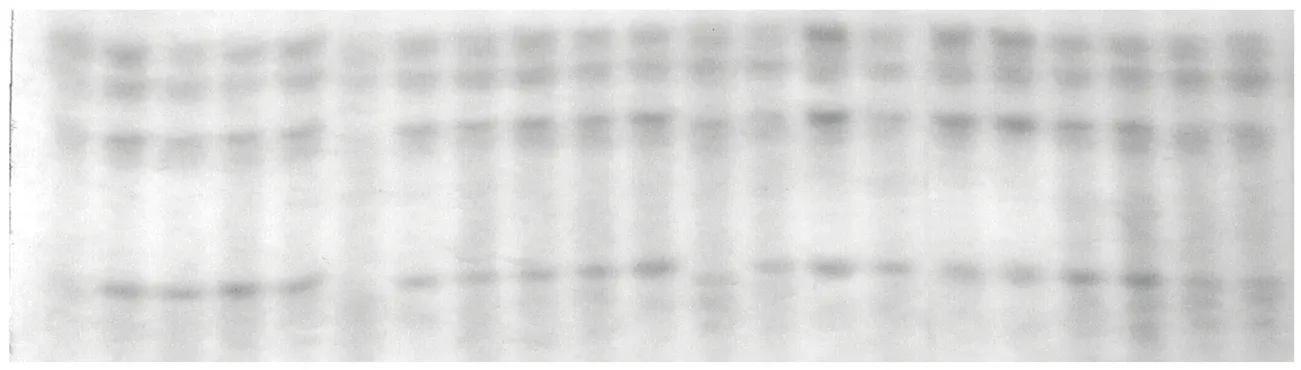
圖2 青燕1號燕麥品種種子純度檢測電泳圖譜
2.2 遺傳相似系數
基于醇溶蛋白數據,計算供試15個品種間的DICE遺傳相似系數,其變異范圍為0.143~1.000,平均值為0.527,說明供試品種間遺傳變異較大。其中青莜3號與青海甜燕麥的遺傳關系最遠,相似系數最低,為0.143;青海444和青燕1號的醇溶蛋白圖譜完全一致,遺傳相似系數為1.000,親緣關系最近。
2.3 聚類和主成分分析
基于醇溶蛋白的聚類分析結果(圖3)表明,供試的15個燕麥品種被分成6類。砂燕麥與其他燕麥品種的遺傳距離最遠,單獨聚為一類(Ⅰ)。第Ⅱ、Ⅵ類均只包含一份材料,分別為阿壩燕麥和青引1號,其表現出了與其他品種的特異性。第Ⅲ類包括了青海444、青燕1號和青海甜燕麥,其中青海444與青燕1號的親緣關系非常近。第Ⅳ類包括了青引2號、莫妮卡和駿馬3個品種。第Ⅴ類包括剩下的6個品種,其中林納、青莜3號又聚為一個亞類,青引3號與加燕2號聚為一個亞類,大漢與白燕7號聚為一個亞類。主成分分析結果與聚類分析結果基本一致,其中前3個主成分分別可以解釋總遺傳變異的56.74%、14.39%、7.35%(圖4)。
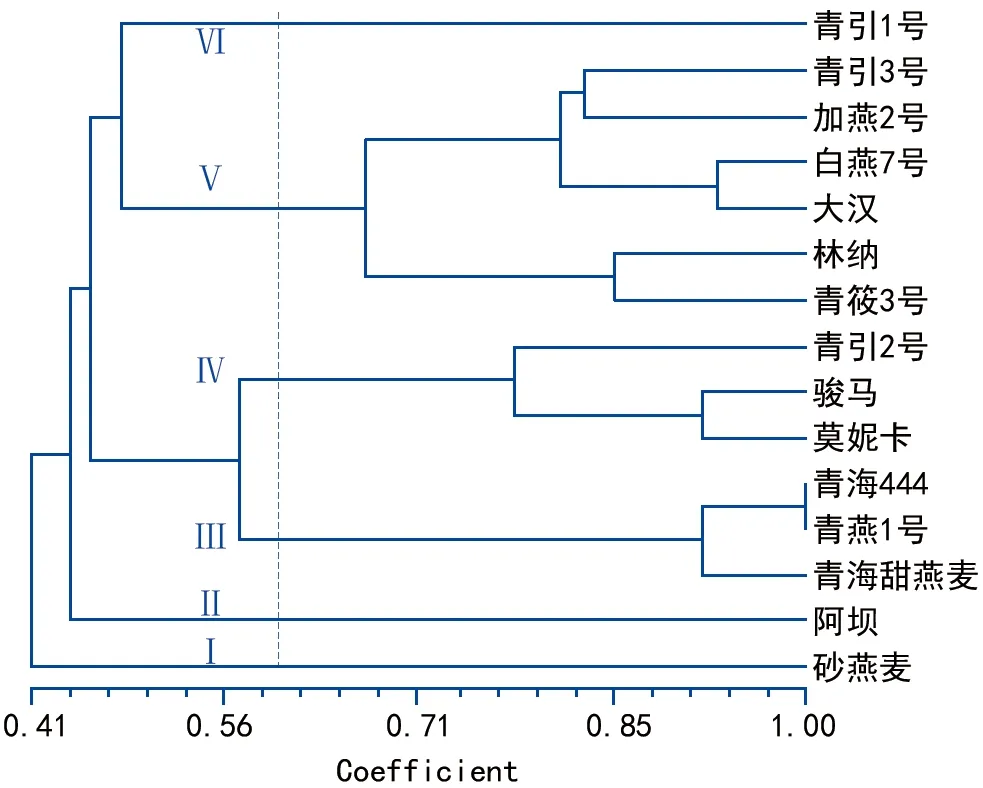
圖3 聚類分析圖
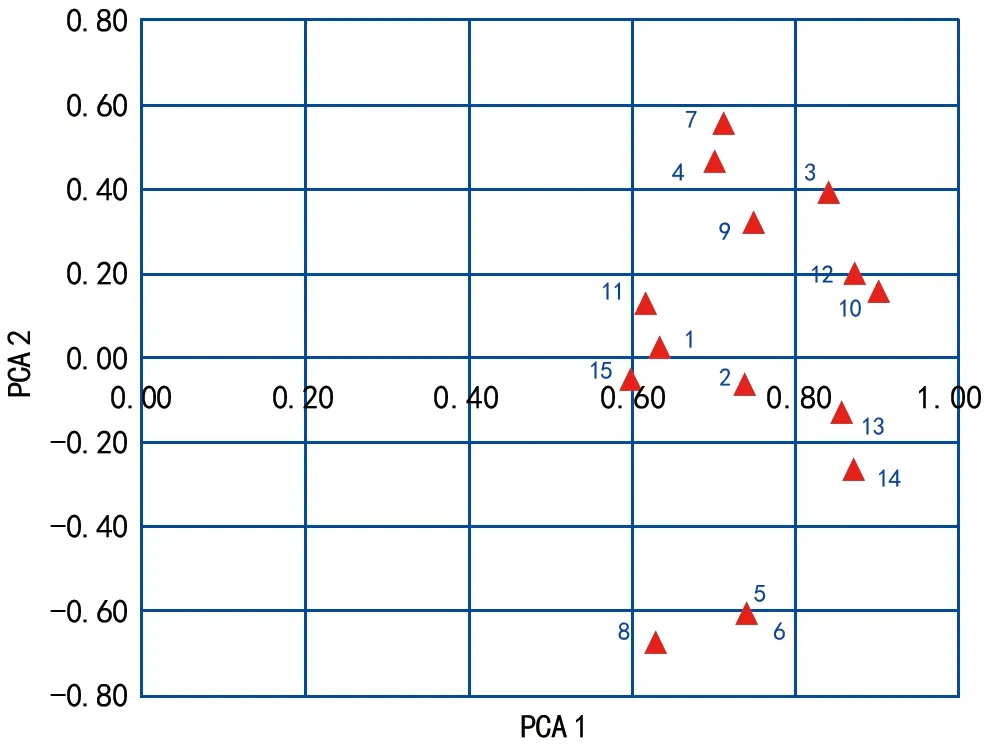
圖4 主成分分析
3 討 論
麥類作物及其近緣物種種子中的醇溶蛋白帶譜嚴格受遺傳基因控制,不受環境等外界環境因素的影響,且不同材料或者品種間差異顯著,變異類型也較豐富,目前已經廣泛應用于小麥、燕麥等麥類作物及近緣物種品種真實性及純度鑒定等研究[13]。齊冰潔等[8]采用A-PAGE電泳對74份燕麥進行分析,共分離出19種不同的醇溶蛋白帶紋,73種醇溶蛋白圖譜,揭示其存在豐富的等位變異。羅桂花等[14]采用醇溶蛋白電泳分析了42份青海燕麥農家品種,其中包括29份皮燕麥,13份裸燕麥,共檢測出6條遷移率不同的譜帶,5種電泳圖譜類型,其變異類型較少,可能與其均來自青海地區有關。劉敏軒等[15]采用醇溶蛋白超薄層等電聚焦電泳技術分析了7個飼用燕麥品種,共分離出34條不同遷移率的譜帶,其中85%的具有多態性,并成功分離鑒定了7個燕麥品種及其純度。本研究采用A-PAGE電泳對15個品種進行分析,共檢測到18條不同的醇溶蛋白位點,其多態性達到100%,存在14種圖譜類型,成功分離鑒定出13個品種。這些結果表明,醇溶蛋白電泳能夠作為一種簡單、快速、有效的手段進行燕麥品種真實性及純度的鑒定。
聚類分析結果表明,二倍體栽培燕麥類型砂燕麥與其他六倍體栽培燕麥的遺傳關系較遠,分化非常明顯。而供試材料中的皮燕麥和裸燕麥的分化反而不是很明顯,如青莜3號裸燕麥與林納皮燕麥親緣關系更近,青引3號與加燕2號親緣關系相對更近。這也表明在燕麥種質等研究中,倍性對燕麥種質特性的影響更為明顯,這在不同倍性燕麥的抗旱性評價中得到了印證[16]。在六倍體栽培燕麥中,阿壩和青引1號燕麥也與其他品種間表現出了一定的特異性,在生產上也表現出早熟、高產等特性,可以為后續的雜交種質組配等提供參考。另外,青燕1號、青海444和青海甜燕麥3個品種的親緣關系較近,特別是青海444與青燕1號的醇溶蛋白帶譜完全一樣,其在種子的外稃上均呈現出黑色。下一步研究將借助SSR等高分辨率的分子標記進行進一步的區分和分子指紋圖譜的構建。
4 結 論
本研究采用A-PAGE電泳對15個飼用燕麥品種的醇溶蛋白進行分析,共擴增出18條帶,多態性達到100%,結果揭示了供試品種間具有較大的遺傳變異。聚類分析等結果表明,砂燕麥、阿壩、青引1號與其他品種親緣關系較遠,其中青燕1號和青海444的親緣關系最近。
