互花米草入侵對鳥類的生態影響
2019-05-13 07:06:32朱曉靜魯長虎
生態學報 2019年7期
陳 潘,張 燕,朱曉靜,魯長虎,*
1 南京林業大學生物與環境學院,南京 210037 2 安徽師范大學環境科學與工程學院,蕪湖 241000
生物入侵嚴重威脅全球生物多樣性[1- 2],直接或間接影響本地生物類群以及生態系統的功能和服務,是目前導致生物多樣性喪失的最主要原因之一[3- 4]。入侵植物可以強烈的改變生態系統中的生物和非生物特征[5],影響本地植被、昆蟲、哺乳類、鳥類等許多生物類群[6]。入侵不僅改變本地植被群落影響鳥類棲息地的組成,也通過改變食物網結構影響鳥類的種群數量和物種多樣性[7],入侵植物對鳥類的生態影響正受到越來越多的研究關注[8]。
互花米草(Spartinaalterniflora)隸屬禾本科(Gramineae)米草屬(Spartina),為多年生草本植物,原產北美洲大西洋沿岸和墨西哥灣,由于其耐鹽、耐淹和促淤護堤作用,曾被世界多地引種,用于改善沿海生態環境[9]。但互花米草往往快速增殖取代本地植被,改變原有生態系統的結構和功能,同時影響鳥類等多種生物類群。互花米草入侵沿海濕地導致的鳥類棲息地退化和生態影響在世界范圍內引起了長期、廣泛的關注,例如:新西蘭[10]、英國南部海岸河口[11]、澳大利亞[12- 13]、美國舊金山海灣[14- 15]、西班牙[16]等地均有相關研究報告。互花米草自20世紀70年代引入中國,在過去的幾十年其分布面積快速擴散,沿海各省從遼寧到廣西均有分布[17]。其中,近94%的互花米草分布在江蘇、上海、浙江和福建的沿海濕地,對本地土壤地貌、生物類群、生態系統功能等造成嚴重影響[18],已成為我國受關注度最高的入侵物種之一[19]。我國東部沿海濕地是許多珍稀水鳥的重要棲息地,也是東亞-澳大利亞遷徙線路候鳥的重要停歇點[20],分布多個國家級自然保護區,例如:鹽城濕地珍禽國家級自然保護區、崇明東灘鳥類國家級自然保護區等。目前,互花米草已在多個保護區內擴散形成優勢植被群落,對鳥類群落造成多種生態影響[21- 23]。本文系統梳理了國內外關于互花米草入侵對鳥類生態影響的研究進展,分析了入侵對鳥類棲息地、食物資源、繁殖、群落等方面的抑制和促進作用,對未來研究方向以及互花米草的清除治理提出了建議。
1 互花米草入侵改變鳥類棲息地格局
植被群落是鳥類棲息地的重要組成部分,是構建鳥類合適棲息地的基礎[24]。由于互花米草較強的適應性及溫和的入侵地環境條件,入侵后常常能夠快速增殖,在競爭中取代本地物種,在潮灘形成單優勢植物群落,改變本地植被群落類型和分布[25]。互花米草入侵對植被群落的影響在一定程度上也改變了沿海灘涂鳥類棲息地景觀格局[26- 27]。
米草屬植物對沿海灘涂的侵占,在世界范圍內引起依賴灘涂生境鳥類棲息地的改變。英國威爾士戴菲河大米草(Spartinaanglica)僅增加10%的面積,就已顯著改變鳥類棲息地結構,造成黑腹濱鷸(Calidrisalpina)、劍鸻(Charadriushiaticula)、蠣鷸(Haematopusostralegus)等數量明顯下降[28]。英國南部海岸河口大米草入侵光灘面積的增加導致鸻鷸類的可利用覓食地面積減少[11]。美國太平洋沿岸互花米草在中低潮灘的擴張導致涉禽覓食地的喪失,遷徙鳥類被迫改變覓食地選擇[29- 30]。互花米草入侵美國威拉帕海灣形成高密度單一化的植被群落,嚴重限制了濱鳥的棲息地面積,使水鳥越冬和繁殖關鍵生境減少了16%—20%,導致鸻鷸類在光灘的覓食時間減少了50%[31]。對美國舊金山海灣濕地鳥類的環境容納量分析發現,互花米草擴張使鳥類的棲息地面積減少了33%[32]。雖然美國上個世紀利用疏浚棄土種植互花米草建造的人工沼澤為一些鳥類提供了棲息地,但因理化特征的差異導致鳥類群落結構組成明顯與自然濕地不同,不能完全替代自然濕地功能[33]。
遙感監測顯示,互花米草入侵導致我國沿海紅樹林、潮灘蘆葦(Phragmitesaustralis)、堿蓬(Suaedaglauca)等鳥類適宜生境面積銳減[17],造成沼澤濕地中一些專性和瀕危鳥類棲息地喪失[34]。互花米草入侵顯著地將泥灘轉變為草地,改變了棲息地的景觀格局,研究顯示絕大多數鳥類選擇泥灘或蘆葦生境,而被互花米草入侵后的泥灘和蘆葦生境可利用性降低,不利于鳥類的棲息[21]。在我國江蘇鹽城國家級自然保護區,互花米草入侵改變了植被群落格局,嚴重阻礙了丹頂鶴(Grusjaponensis)的活動,降低了該區域作為丹頂鶴棲息地的價值[27]。對互花米草的清除實驗表明,治理后的恢復生境可為雁鴨類等重要水鳥提供良好的棲息地,生物多樣性維持的生態服務功能有了明顯的恢復[35]。西班牙奧迪埃爾沼澤在清除入侵植物密花米草(Spartinadensiflora)并栽種本地植被的兩年后,鳥類多樣性和棲息地質量得到顯著提高[16]。
但也有一些研究認為互花米草入侵可能促進了生態系統的演化,為一些鳥類提供了合適棲息地[15,23]。對崇明東灘互花米草入侵的長期監測顯示,互花米草種群在一個5年的高速增長期后會有一個更長時間的衰退。互花米草會促使灘面淤高造成潮水量減少,導致互花米草退化和死亡,這又為本地堿蓬、蘆葦的生長提供了演替條件[36]。互花米草還可以在其他植被不宜生長的光灘生長,作為先鋒植物為紅樹林等其他植被的演替創造條件[37]。美國威拉帕海灣的研究顯示,在低潮灘互花米草往年枯死的植株會被潮水沖刷掉,且春季新生植株相對比較稀疏,為鳥類提供了可利用的棲息地,春季遷徙期一些鳥類會在互花米草群落中覓食,并利用互花米草群落作為隱蔽所躲避猛禽等天敵的捕食[38]。有學者認為,我國沿海光灘新形成的互花米草群落可以為一些鳥類提供覓食、筑巢等棲息地[39- 40]。近年有研究認為入侵植物與本地生態系統各類動植物間存在長期的相互作用,對入侵植物的快速清除有時可能對生態系統帶來副作用,反而危及部分鳥類的棲息[41]。一些鳥類已經適應入侵互花米草對自然生境的改造,例如入侵互花米草群落是加州長嘴秧雞(Ralluslongirostris)的重要棲息地[42],在崇明東灘對斑背大尾鶯(Megaluruspryeri)的研究也證實了這一觀點[23]。
2 互花米草入侵對鳥類食物資源的影響
外來植物的入侵不僅改變本地植被群落,也通過改變食物鏈影響著鳥類[22]。作為較高營養級,許多鹽沼鳥類依靠本地植物種子、塊莖以及昆蟲和大型底棲動物作為食物來源[21]。互花米草入侵導致植被、昆蟲、大型底棲動物的分布和多樣性發生變化[43- 45],這可能間接影響到鳥類的可利用食物資源和覓食行為。
2.1 對植物性食物的影響
本地優勢植被的種子、塊莖是一些鳥類的重要食物資源[43]。入侵的互花米草競爭替代了本地優勢植被,減少了鳥類的食物資源。研究顯示,雁鴨類、小天鵝和白頭鶴等大型鳥類均以海三棱藨草(Scirpusmariqueter)等本地植被的球莖為食,并不采食互花米草,因此互花米草入侵降低了棲息地對鳥類群落的承載力[21]。對崇明東灘進行互花米草移除后,本地植被的恢復為雁鴨類提供了豐富的植物性食物資源,鳥類物種多樣性得到提高[35]。雖然大多數本地鳥類不會取食互花米草種子和根莖,但一些小型鳥類也會在互花米草群落中覓食[15,22],可能是受限于種間競爭壓力選擇次優食物資源。
2.2 對昆蟲群落的影響
昆蟲與植物、鳥類關系密切,是生態系統的重要組成部分,大多數鳥類食譜都包含一定數量的昆蟲[6]。外來植物入侵后,不同昆蟲類群響應機制不同,導致昆蟲數量和群落組成發生變化[46]。崇明東灘互花米草入侵后冬季昆蟲食物資源與鳥類捕獲率之間的關系研究顯示,互花米草生境中昆蟲密度顯著低于蘆葦生境,并且鳥類糞便中的食物結構都與蘆葦群落中的食物資源結構更為相似[22]。對互花米草與本地植被中昆蟲群落的比較研究認為,互花米草分布區昆蟲數量、多樣性均顯著低于蘆葦分布區。在三類群落(蘆葦、堿蓬、互花米草)中,互花米草群落的昆蟲種類和數量最少[47],不同群落中的優勢昆蟲種類不同,昆蟲群落結構發生明顯變化[44]。但也有一些研究顯示,入侵植物和本地植物組成的混合群落增加了昆蟲的物種多樣性,為鳥類提供了更豐富的食物資源[48]。目前,還沒有研究顯示互花米草的入侵可以顯著增加昆蟲的種類和數量,可能是因為本地昆蟲與互花米草缺乏協同進化的過程,需要進一步的深入研究。
2.3 對大型底棲動物的影響
沿海灘涂濕地具有豐富的底棲動物類群,是許多鳥類的重要覓食場所,尤其對遷徙鳥類的中途停歇、能量補充具有重要意義[14,49]。互花米草入侵對大型底棲動物類群的復雜影響,可能通過食物網間接影響著鳥類群落[21]。許多研究揭示了互花米草入侵對大型無脊椎動物類群的影響,但至今還沒有統一的結論[45]。不少研究認為互花米草入侵改變了灘涂植被群落、土壤結構等棲息地因子,顯著降低了大型底棲動物的密度和豐富度[50- 51]。部分研究認為互花米草入侵后提高了灘涂濕地初級生產力,提高了大型底棲動物的總資源量[52]。還有一些研究認為互花米草入侵對大型底棲動物沒有顯著影響,種類數量和多樣性在不同生境中總體差異不顯著[53- 54]。這些不同的研究結果表明,互花米草入侵對底棲動物類群可能存在較為復雜的影響機制。候森林等[55]在江蘇射陽口的研究顯示,潮間帶互花米草的入侵降低了大型底棲動物的物種數、密度、多樣性,但在潮溝地帶互花米草的入侵卻提高了相應指數。還有研究證實,即使互花米草自然演替死亡,但密集的根系改變了棲息地基質的理化性質,導致底棲動物群落在短期內仍無法恢復,間接影響鳥類種群數量的恢復[56- 57]。互花米草的入侵史、入侵程度、研究地特征、底棲動物類群等因素都可能導致不同的研究結果:(1)互花米草入侵時間越長,與本地植被生境底棲動物群落的差異可能越小[45],可能是因為本地動物類群對環境變化的逐步適應;(2)光灘生境更容易受到互花米草入侵的影響,較大的環境改變導致底棲動物類群差異較大[53];(3)不同底棲動物類群對互花米草入侵的響應不同,例如:互花米草入侵后,腹足類、雙殼類數量下降顯著[53],蟹類動物量卻顯著增加[58]。
互花米草入侵導致鳥類潛在食物資源的變化,可能逐步改變了鳥類的覓食地選擇、覓食行為規律、覓食效率等,最終影響鳥類的身體條件、種群數量、繁殖成功率等。
3 互花米草入侵對鳥類繁殖的影響
鳥類的成功繁殖依賴合適的棲息地[24]。許多研究顯示入侵植物給本地鳥類繁殖帶來負面影響,降低了繁殖成功率、雛鳥生長速率、巢址忠誠度等[8]。大多數繁殖期鳥類選擇本地植被群落筑巢,并避免利用互花米草生境,尤其是互花米草取代本地植被形成的單一密集群落,很難發現本地常見鳥類巢址[34,57]。在我國江蘇鹽城國家級自然保護區,20%—40%的堿蓬覆蓋率是黑嘴鷗(Larussaundersi)、普通燕鷗(Sternahirundo)的理想繁殖生境,互花米草入侵縮減了堿蓬面積,導致該區域鳥類筑巢數量下降[26]。在崇明東灘的研究顯示,本地專性鳥類傾向于選擇本地蘆葦生境筑巢,泛性鳥類則會選擇蘆葦-互花米草混合生境,沒有鳥類對單一互花米草生境表現出正選擇[59]。這可能是因為互花米草改變了棲息地結構,造成適宜的巢址微生境多樣性下降,過度均質化的生境不能夠為鳥類提供合適巢址[60]。
但也有研究顯示,入侵植物有時可以為鳥類提供新的繁殖棲息地,沒有造成繁殖成功率的下降[48]。在上海崇明東灘,擴散的斑背大尾鶯專性在互花米草群落營巢,且繁殖成功率與其在中國北方、日本的原產地幾乎沒有差異[59]。在美國舊金山海灣,加州長嘴秧雞、沼澤鷦鷯(Cistothoruspalustris)、阿拉米達歌帶鹀(Melospizamelodia)等鳥類也會在互花米草入侵生境中筑巢繁殖[15,42,61]。這些體型較小的鳥類選擇在互花米草群落中筑巢,可能是迫于種間競爭壓力[59]。因為多數本地鳥類不利用互花米草生境,茂密的植株也為小型種類提供了較好隱蔽。還有研究證實,在一些本地植被退化的生態系統,某些鳥類甚至需要依賴入侵植物完成生活史,優先選擇入侵植物作為巢址[62]。例如:美國加州長嘴秧雞高度依賴互花米草入侵生境筑巢繁殖,對互花米草的清除治理已嚴重威脅到該物種的正常繁殖[42]。
雖然一些鳥類能夠在互花米草生境中繁殖,但可能面臨風險。研究顯示,阿拉米達歌帶鹀在互花米草群落中筑巢的繁殖成功率比在本地植被群落中低30%,可能是因為互花米草大多生長在較低海拔,高潮期潮水會淹沒部分鳥巢導致繁殖失敗[61]。入侵植物對植被群落結構的影響有時會改變巢址的被捕食率,鳥類在入侵植物中筑巢繁殖較易受到嚙齒類的破壞,雛鳥被捕食率顯著高于本地植被群落[63]。郝志等[64]在互花米草群落中設置人工巢,結果顯示高達67.7%的鳥巢被嚙齒類等捕食,可能是因為互花米草群落中嚙齒動物天敵的缺乏。互花米草入侵是否導致繁殖鳥類巢捕食率升高,還需要進一步的研究證實。
4 互花米草入侵在鳥類群落層面上的影響
本地植被類型對維持鳥類數量、物種多樣性、均勻度具有重要作用。外來植物入侵引起的植被群落結構、棲息地景觀格局、可利用食物資源的變化,最終都會影響本地生境中鳥類種群數量和物種多樣性[48]。
4.1 對鳥類群落的抑制作用
許多研究表明,互花米草入侵導致本地鳥類種群數量和多樣性明顯下降,尤其對棲息地專性鳥類和鸻鷸類、雁鴨類、鶴類等中大型水鳥群落抑制作用顯著。對崇明東灘春季鳥類群落結構的研究顯示,超過90%的鳥類都選擇本地植物組成的棲息地,互花米草群落中鳥類的物種數和密度都顯著低于其他棲息地類型[34]。震旦鴉雀(Paradoxornisheudei)、東方大葦鶯(Acrocephalusorientalis)等專性鳥類避免使用互花米草形成的單一群落,只有一些泛性鳥類選擇互花米草棲息地,但數量也顯著低于周圍蘆葦生境[34,65]。在鹽城自然保護區,互花米草群落中冬季鳥類多樣性指數在各類生境中最低[66]。對崇明東灘繁殖期鳥類群落結構的研究顯示,泛性鳥類的物種數和多度在各類生境中差別不大,專性鳥類在蘆葦生境和互花米草-蘆葦混合生境中差別不大,但在單一互花米草生境中則顯著下降[59]。
互花米草入侵對于依賴灘涂濕地覓食的遷徙和越冬期水鳥群落可能更為不利,造成世界范圍內許多河口及海岸濱鳥種群數量的下降。相比較互花米草生境,大多數鸻鷸類更傾向于選擇光灘或本地植被生境,入侵引起的光灘向草地的轉變已嚴重影響了水鳥的棲息[43,49,57]。在美國威拉帕灣,互花米草面積的逐年增加,造成遷徙過境的水鳥數量逐年下降[14],互花米草的擴張導致1991年至2001年間鸻鷸類的數量減少了67%[31]。在英國南部海岸,大米草的入侵導致濱鳥的死亡率和遷出率上升[11],黑腹濱鷸種群數量的下降甚至在大米草自然演替枯萎后也很難恢復[56]。在我國鹽城保護區,丹頂鶴依賴草灘、葦灘等生境覓食,互花米草在生長穩定期蓋度可達100%,單一互花米草群落內很難見到丹頂鶴分布[67]。在崇明東灘的互花米草移除示范區內,水鳥的種類與數量均高于對照區,說明互花米草治理后生境能夠吸引鸻鷸類、雁鴨類等等水鳥越冬棲息和覓食,鳥類群落結構多樣性得到改善[68]。
4.2 對鳥類群落的促進作用
外來入侵植物對本地植被群落結構的改造,對不同的鳥類可能影響不同。一些入侵植物可以提供本地鳥類無法利用的生態位,吸引其他非本地物種棲息利用,在一定程度上促進了本地物種的多樣性[69-70]。雖然許多本地鳥類避免使用互花米草生境,但這也為一些鳥類留下了空白生態位,一些小型雀形目鳥類可以很快適應互花米草入侵生境。在崇明東灘的研究顯示,一些體型較小的鳥類在種間競爭的壓力下會選擇鑲嵌群落或互花米草群落作為次優棲息地[59]。在某些林地生態系統,入侵植物增加了鳥類棲息地復雜程度,提供了更多的適宜生態位和棲息地類型[8]。在互花米草入侵形成穩定植被群落后,非本地繁殖鳥類斑背大尾鶯首次出現在崇明東灘。與本地蘆葦生境相比,互花米草群落較少的競爭和較密的莖桿或許能夠提供更好的棲息與繁殖生境,并為斑背大尾鶯提供了主要食物來源[23]。這項研究表明,互花米草入侵可能提供了本地植被不具備的適宜生境,新的棲息地選擇會擴大一些鳥類的分布范圍。在美國舊金山海灣的研究也證實,互花米草為體型較小的沼澤鷦鷯提供了合適棲息地,使其擴大了分布范圍[15]。
隨著外來植物入侵歷史的增長,本地生物類群有時可以逐漸適應入侵生境,當移除入侵植物后反而引起物種多樣性下降[71]。在較早的研究中認為加州長嘴秧雞只有在海水高潮期才會利用互花米草生境[72],但隨著互花米草入侵歷史的增加以及與本地米草的雜交,加州長嘴秧雞開始適應新的米草群落,并逐漸依賴該生境棲息覓食,對互花米草的快速清除反而造成該種鳥類種群數量的下降[42]。
5 小結與展望
互花米草作為促淤護堤的有效物種,在引種初期起到了良好效果。但隨著互花米草的快速擴散增殖,改變了本地植被群落結構,顯著降低了棲息地質量,對鳥類等生物類群的負面影響逐漸顯現[21]。主要表現在以下方面:(1)植被群落結構不利于鳥類棲息、筑巢、覓食[15,26]。尤其對于鸻鷸類、雁鴨類、鶴類等需要利用開闊棲息地的中大型鳥類,過密且均質化的植被類型可能阻礙了這類水鳥對互花米草生境的利用,在我國鹽城保護區丹頂鶴對互花米草沼澤景觀具有負向選擇[73]。(2)本地鳥類食物資源豐度和多樣性下降[22,50]。互花米草入侵引起本地植被、底棲生物、昆蟲等鳥類潛在食物資源量下降,即使某些類群數量沒有下降,整體食物資源多樣性也顯著降低,無法滿足鳥類的覓食需要。(3)本地鳥類種群數量和物種多樣性顯著下降[14,59]。互花米草入侵后,許多本地鳥類避免使用互花米草生境,種群數量、物種多樣性、繁殖成功率等均受到影響,導致本地鳥類群落結構發生變化。在我國鹽城沿海,互花米草入侵已經顯著改變了本地植被群落分布,進而影響到鳥類棲息地選擇、種群數量、群落結構等,入侵后的植被與鳥類分布格局形成了一定的梯度模式(圖1)。
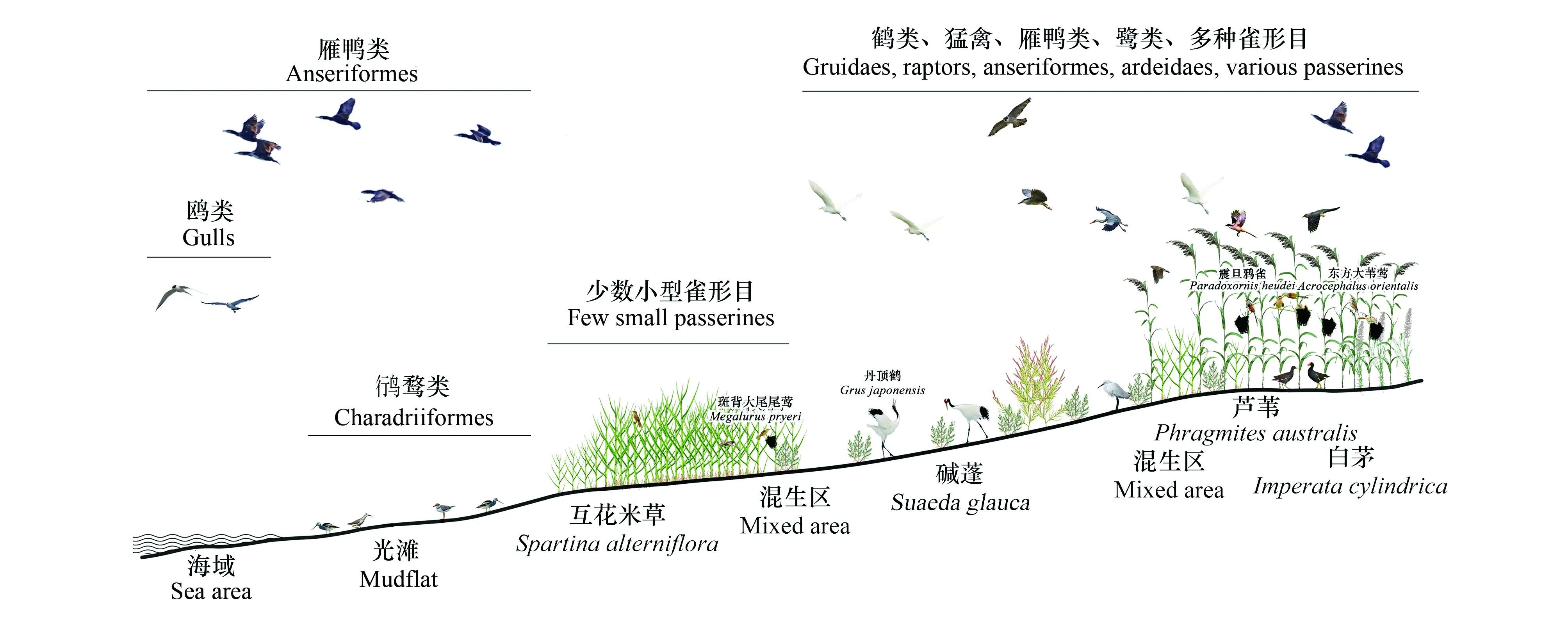
圖1 中國鹽城沿海互花米草入侵生境植物群落及主要鳥類類群分布模式圖Fig.1 Distribution pattern of plant community and main bird groups inSpartina alterniflorainvasive habitat on the Yancheng coast of China
但隨著入侵歷史的增長,互花米草對本地鳥類群落可能產生不同影響[41]。或許由于長期的協同進化,某些鳥類開始逐步適應互花米草群落,選擇入侵生境作為棲息、覓食地,一些種類甚至能夠在其中筑巢繁殖[15]。但只限于極少數小型雀形目鳥類,大多數本地鳥類未表現出對互花米草生境的適應,鳥類群落是否隨入侵時長發生變化需要進一步的研究。互花米草入侵可能還為非本地鳥類提供了本地鳥類無法利用的空白生態位,增加了一些鳥類的分布范圍[23],在一定程度上豐富了本地鳥類物種多樣性。因此,互花米草入侵對鳥類群落甚至整個生態系統的影響可能需要更多研究進行綜合評價。
未來的研究應關注以下兩個方面:(1)開展長時間、大尺度、多因子的監測研究。入侵植物對鳥類的生態影響往往隨入侵生境類型、物種特征、斑塊成熟程度、入侵歷史等因素發生變化,不同地點、不同時期對同一入侵植物的評價可能不同[8]。了解互花米草入侵生境中鳥類的種群、群落乃至生態系統的動態變化, 需要標準化的研究方法進行長期的觀察和跟蹤。應根據長時間、大尺度、多因子的研究數據,建立互花米草入侵對鳥類群落以及生態系統影響的綜合評價模型,從而能夠預測互花米草入侵對鳥類等生物類群的潛在影響以及生態系統結構和功能的變化趨勢。(2)開展多物種的比較研究,制定合理的互花米草管理對策。目前,對互花米草入侵背景下特定鳥類物種的覓食、繁殖僅有少數研究[23,42,62]。互花米草入侵可能對不同鳥類物種產生不同影響,全面系統的了解互花米草入侵對鳥類群落的影響機制,需要開展多物種的種間比較研究,還需要考慮互花米草入侵對鳥類的食物、天敵等其他生物類群的影響等。在我國崇明東灘實施的互花米草治理項目優化了鳥類棲息地,取得了顯著的生態效果[74],互花米草的清除和本地植被的恢復可以顯著改善本地鳥類群落狀況。但對美國加州互花米草的清除卻對已適應并依賴互花米草生境的鳥類造成影響[42],由于動植物間的協同進化關系,對入侵植物的清除有時甚至產生強烈的負面生態效應[75]。在不同地區開展互花米草治理過程中可能需要考慮更多因素,而不是簡單的快速清除,應根據入侵生境類型、鳥類物種、植被狀況等,制定因地制宜、科學有效的管理對策,逐步恢復本地植被生境。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國塑料(2016年3期)2016-06-15 20:30:00
