安慶西江長江江豚遷地保護基地河岸帶植物群落結構特征
2019-05-29 06:58:42馬鳳嬌藺丹清張曉可尹登花張家路
水生生物學報 2019年3期
馬鳳嬌 藺丹清 張曉可 尹登花 張家路 陶 峰 劉 凱
(1. 中國水產科學研究院淡水漁業研究中心, 農業部長江下游漁業資源環境科學觀測實驗站, 無錫 214081;2. 南京農業大學無錫漁業學院, 無錫 214081; 3. 安慶師范大學生命科學學院, 安慶 246133)
河岸帶屬于水陸生態交錯區, 大多數學者把河岸帶定義為高低水位之間的河床及高于水位之上的水陸交界處, 直至河水影響消失為止的地帶[1]。由于河岸帶地理位置特殊, 并受河流地貌和水文環境的雙重影響, 因此兼具了河流和陸地的特征,是水域生態系統和陸地生態系統之間的緩沖區域[2,3]。河岸帶復雜的生境使其具有很高的生物多樣性[4], 保證了河流生態系統結構和功能的完整性,因此河岸帶植物群落的物種組成和群落特征尤為重要。此外, 作為河岸帶生態系統的重要組成部分,河岸帶植被具有調節水質、為其他生物提供避敵場所、防止水土流失以及維持堤岸的穩定性等重要功能。
長江江豚(Neophocaena asiaeorientalis)是江豚屬(Neophocaena)唯一的淡水種群, 僅分布于長江中下游干流及洞庭湖和鄱陽湖[5—7]。20世紀末至今,隨著人類活動影響的不斷加劇, 與魚類及其他水生生物資源急劇衰退的趨勢相似, 長江江豚種群數量也急劇下降, 直到2012年江豚種群數量仍以每年13.7%的速度銳減, 極度瀕危[8]。2017年長江江豚科學考察結果顯示, 長江江豚種群數量約為1012頭,長江江豚種群數量大幅下降的趨勢得到遏制, 但其極度瀕危的狀況沒有改變、依然嚴峻, 亟待保護[9]。安慶西江在歷史上是位于長江下游北岸的一條支航道, 為了減輕防洪壓力, 分別于1979和2007年人工將其上、下游出口封堵, 形成長度約為9 km的故道, 但下游通過閘門仍然與長江干流季節性連通, 這與長江天鵝洲豚類自然保護區具有相似的生境[10]。西江兩岸基本為自然岸帶, 受人類活動干擾較少, 水質良好, 漁業資源豐富。2016年10月,農業農村部實施長江江豚遷地保護行動, 先后將13頭長江江豚遷入西江, 建立安慶市西江長江江豚遷地保護基地。鑒于安慶西江的重要性, 目前關于西江水質和浮游動物[11]、魚類[12]的研究均已有報道, 但作為西江主要初級生產者的河岸帶植被一直未見報道。本研究的目的在于通過對西江河岸帶植物進行系統調查, 掌握其基本群落特征, 以期為西江長江江豚棲息地評價提供基礎數據, 并為西江長江江豚遷地保護基地的科學管理提供支撐。
1 材料與方法
1.1 研究區域概況
西江位于長江下游北岸, 地理位置為E: 116°28′—117 °03′, N: 30 °20′—30 °50′。東與安慶市毗鄰, 西連潛山、太湖, 北接桐城, 南鄰望江, 與東至隔江相望。氣候處于北亞熱帶向中亞熱帶季風濕潤氣候的過渡地帶, 四季分明, 氣候溫和。
1.2 研究方法
基于均勻布點規則及西江兩岸植被分布情況,共設置10個取樣斷面, 其中北岸5個斷面, 編號為N1—N5; 南岸5個斷面, 編號為S6—S10, 每個斷面沿水分垂直梯度降低的方向上, 設置3個樣方, 共計30個樣方。斷面分布見圖1。
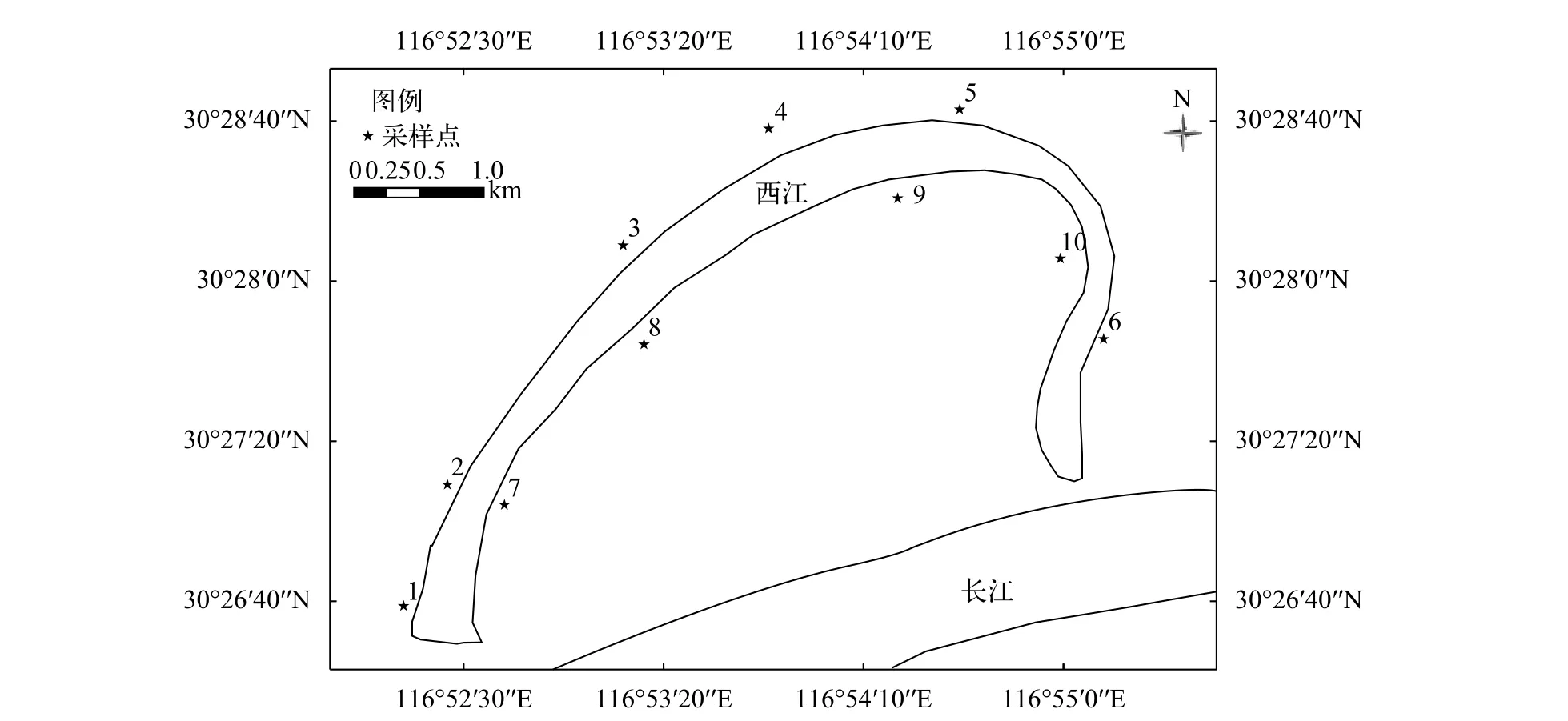
圖1 西江濕地植被采樣斷面分布圖Fig. 1 The distribution map of wetland vegetation sampling transects in Xijiang Oxbow
對斷面的植物進行定性和定量采集。定性采集主要記錄各個斷面內所有發現的植物種類。定量采集主要采用樣方法對各個斷面河岸帶植物進行調查, 每個斷面設置3個樣方, 每個斷面沿水域至陸域的方向對草本植物、灌木和喬木分別進行調查; 其中喬木層調查樣方面積為20 m×20 m, 灌木層樣方面積為2 m×2 m, 記錄灌木的種類、高度和蓋度等; 草本植物樣方面積為1 m×1 m, 記錄樣方內植物的種類、高度、蓋度及其數量。采用收割法收割植株地上部分, 在電子天平上稱量其鮮重(g)。在調查過程中盡可能采集標本, 以保證所調查物種具有憑證。在采集過程中詳細記錄與物種相關的形態特征, 物候特征、分布地點、生境等方面信息。采樣斷面具體位置信息見表1。
1.3 數據分析
物種所屬科、屬根據《中國植物志》[13]確定。
采用Shannon-Wiener指數(H′)和Simpson指數(D′)分析植物物種多樣性特征。其計算公式分別為: Shannon-Wiener多樣性指數H′=-∑PilnPi;Simpson多樣性指數D′=1-∑Pi2; 其中,Pi為種i的相對重要值,Pi=Ni/N;Ni為種i的絕對重要值,N為種i所在樣方的各個種的重要值之和。Patrick豐富度指數:R=S;S為每個樣方內的物種數。數據處理利用Excel軟件。
根據水生植物對水環境的適應程度, 按生活型可初步劃分為濕生植物、挺水植物、漂浮植物、浮葉植物和沉水植物。植物科的區系按照吳征鎰(2003)關于中國種子植物科的區系統計分析方法進行, 屬的區系按照吳征鎰(1991)中國種子植物屬的分布區類型進行分析[14,15]。
2 結果
2.1 河岸帶植物區系基本構成
西江河岸帶(濕地和灘地)維管植物共有166種(不包括栽培種), 隸屬于50科131屬, 其中包括8種外來物種, 隸屬于5科8屬(附錄中*標注)。其中藻類植物1科1屬1種; 蕨類植物1科1屬2種; 種子植物48科129屬163種, 又可分為裸子植物和被子植物兩大類群, 裸子植物1科1屬1種; 被子植物47科128屬162種, 其中雙子葉植物39科99屬118種, 單子葉植物8科29屬44種。在科水平上統計, 物種數在10種及以上的科有4科, 為蝶形花科(Papilionaceae)、菊科(Compositae)、禾本科(Gramineae)、莎草科(Cyperaceae), 多為典型的大科, 共65種, 是西江河岸帶植物群落的主要物種。
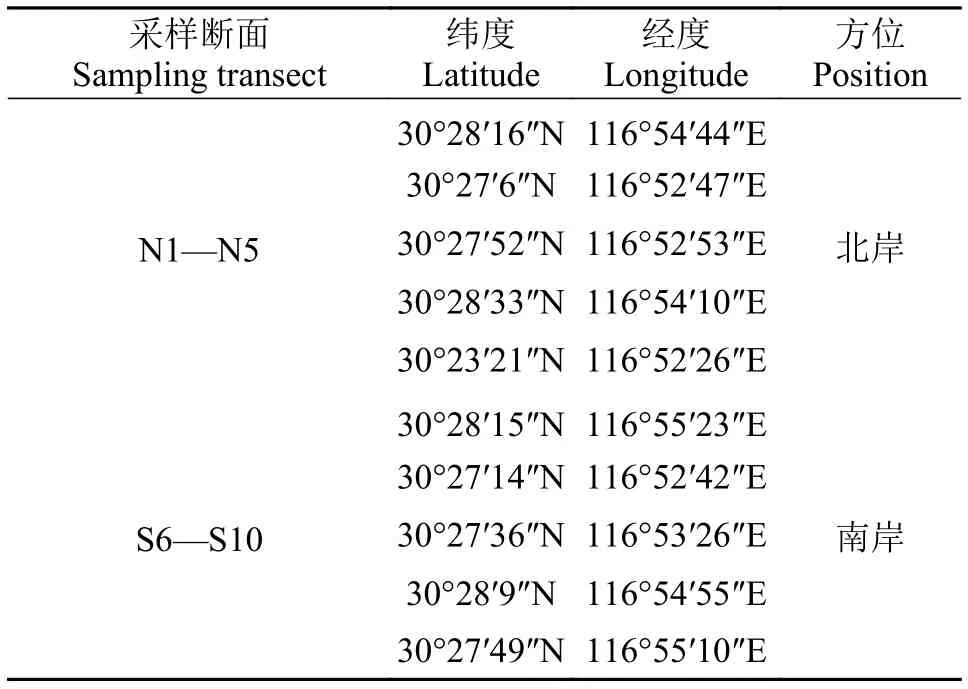
表1 西江采樣斷面坐標數據Tab. 1 Coordinate data of the sampling transects in Xijiang Oxbow
2.2 西江河岸帶植物區系的生活型分析
生活型分析用來研究植物生態功能群與環境之間的關系, 將西江河岸帶植物生活型劃分為濕生植物、挺水植物、漂浮植物、浮葉植物和沉水植物五類。濕生植物共147種, 占物種總數的88.55%,主要包括蕨類植物、裸子植物和被子植物; 水生植物共19種, 占11.45%。在水生植物中, 挺水植物共10種, 占6.02%, 包括蓮(Nelumbo nucifera)、喜旱蓮子草(Alternanthera philoxeroides)、水蓼(Polygonum hydropiper)、蘆葦(Phragmites australis)等;漂浮植物共3種, 占1.81%, 為浮萍(Lemna minor)、紫萍(Spirodela polyrrhiza)、水鱉(Hydrocharis dubia); 浮葉植物共3種, 占1.81%, 為芡實(Euryale ferox)、野菱(Trapa incisavar.quadricaudata)、菱(Trapa bispinosa); 沉水植物共3種, 占1.81%, 為輪藻(Charaspp.)、輪葉黑藻(Hydrilla verticillata)、菹草(Potamogeton crispus)。西江河岸帶植物以濕生植物占優勢, 其次是挺水植物。
2.3 植物區系地理成分分析
對安慶西江河岸帶被子植物47科按照吳征鎰[15]進行分布區類型統計分析, 劃分為5個分布區類型(表2)。世界分布類型所占比例最高, 占總科數的61.70%, 主要有禾本科、菊科、莎草科、蓼科、唇形科、十字花科、玄參科等29科。組成安慶西江被子植物科的分布區類型比較集中, 主要為世界分布、泛熱帶分布, 具有明顯的熱帶親緣性, 但群落分布的優勢科主要還是以廣布性的禾本科、菊科和莎草科為主。
將安慶西江岸帶被子植物128屬進行歸類, 除了熱帶亞洲和熱帶美洲間斷分布、地中海區、西亞至中亞分布、中亞分布和中國特有分布外, 其他11個分布區類型在本區域均有分布(表3)。除世界分布屬外, 以北溫帶和泛熱帶分布屬最多。這表明安慶西江地處亞熱帶, 為熱帶向溫帶過渡地區, 植物區系明顯具有熱帶、亞熱帶與暖溫帶的雙重性質。
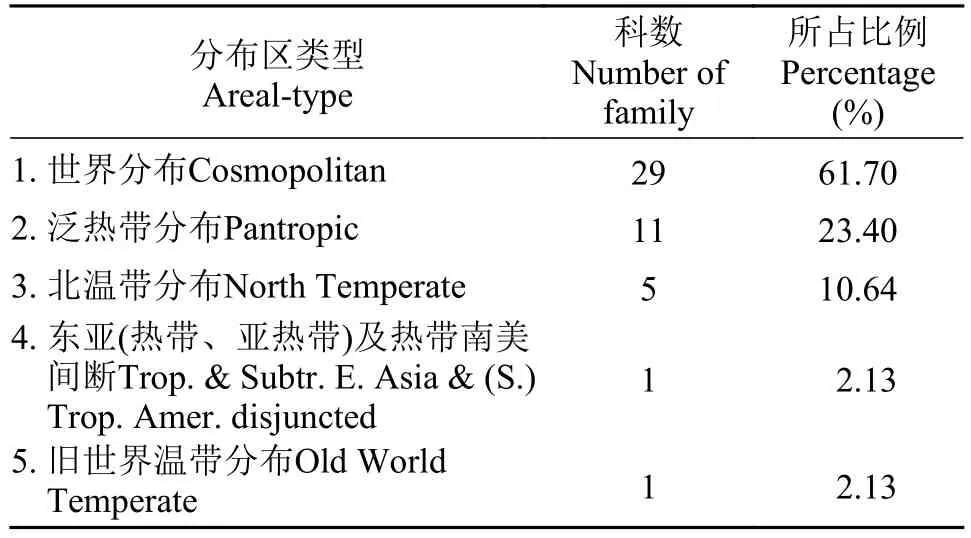
表2 西江河岸帶被子植物科的分布區類型Tab. 2 Areal types of the families of angiosperms in riparian zone of the Xijiang Oxbow
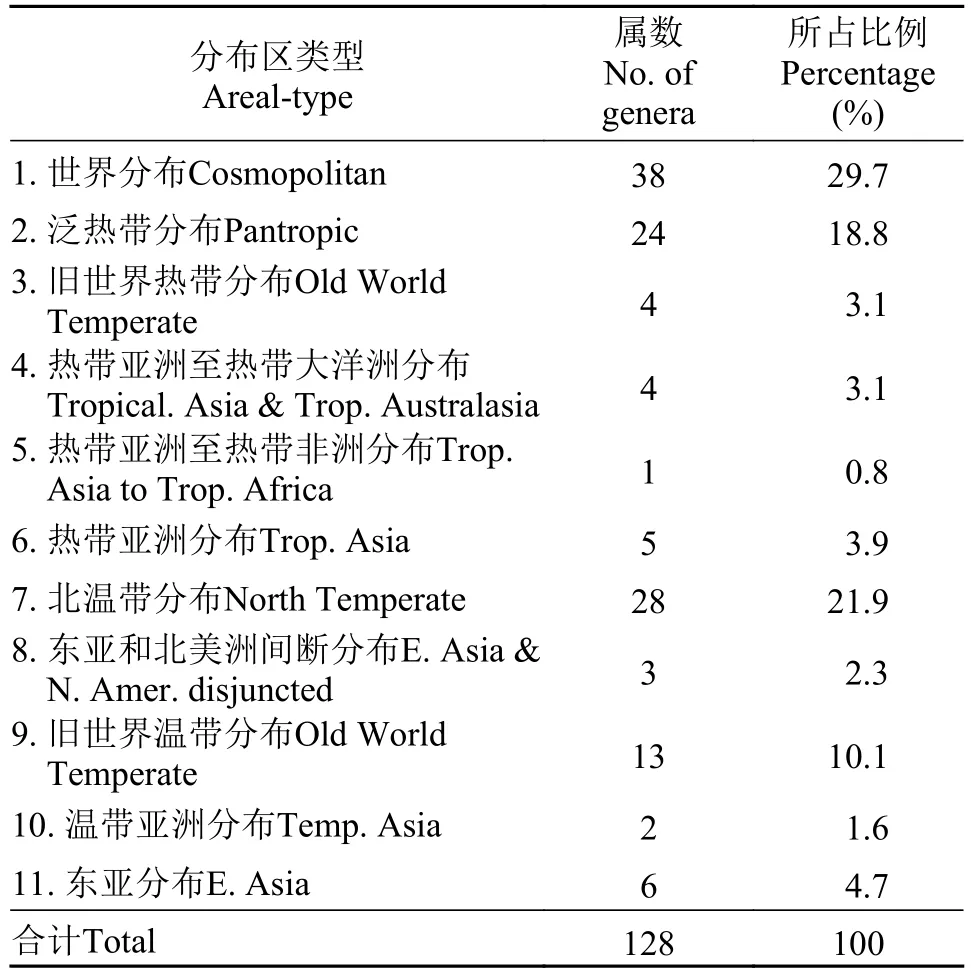
表3 西江河岸帶被子植物屬的分布區類型Tab. 3 Areal types of the genera of angiosperms in riparian zone of the Xijiang Oxbow
2.4 植物生物量
對10個斷面沿著水分梯度對植物群落生物量的分析表明, 生物量變化較為復雜。由表4可知, 西江河岸帶植物群落的平均生物量差異較大。其中,N3斷面的植物群落的平均生物量最小, 為348.00 g/m2, 而最大生物量出現在N5斷面, 可達4346.67 g/m2, 可能是由于受人為影響較小, 其生物多樣性和生態環境受干擾少, 植物覆蓋面積較大, 有利于生物量的累積。植物群落平均生物量變異程度都較大, 其中N5斷面生物量的空間變異最為顯著 (CV=110.01%), 主要是由于不同斷面的各優勢植物的生物量分布差異大導致的。此外, 為了更好地比較西江岸邊不同區域河岸帶植物群落的變化, 本研究采樣地點相對分散, 這可能會對各個斷面群落的差異造成一定影響, 在以后的研究中還需進一步驗證。
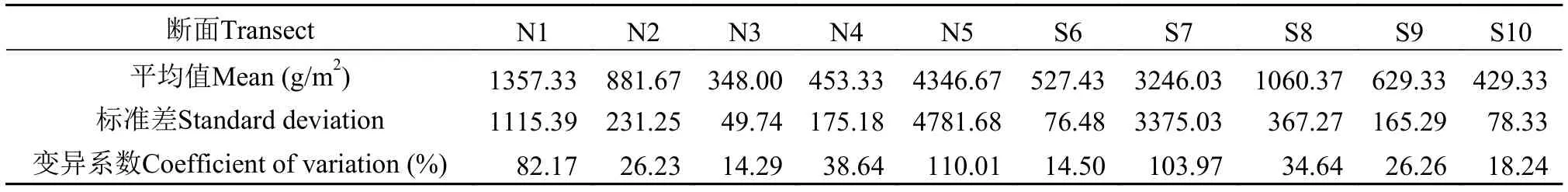
表4 西江河岸帶10個斷面植物群落平均生物量及其標準差和變異系數Tab. 4 The average biomass of plant communities and their standard deviation and coefficients of variance in 10 sampling transects in riparian zone of the Xijiang Oxbow
2.5 植物群落多樣性特征值
西江北岸河岸帶群落的Shannon-Weiner多樣性指數變化為0.213—0.857, 均值為0.563±0.350;Simpson多樣性指數變化為0.135—0.465, 均值為0.292±0.187。西江南岸河岸帶群落的Shannon-Weiner多樣性指數變化為0.553—0.899, 均值為0.696±0.447, Simpson多樣性指數變化為0.252—0.421, 均值為0.337±0.221。北岸植物群落的多樣性指數變化較大(圖2), 隨著土壤水分的減少, 物種多樣性指數總體呈增加的趨勢。在河岸帶部分地段, 由于人類活動(如開挖魚塘)導致帶狀分布特征不明顯。較高的物種多樣性指數出現在北岸, 可能與北岸的灘地面積相對較大, 部分岸帶區域人為影響較少有關。
同時, 豐富度指數的大小也在一定程度上反映了群落生境的特點。西江河岸帶植物群落30個樣方的物種豐富度, 變化范圍在3—14 (圖3)。西江南岸的植物群落的物種豐富度變化相對平穩, 物種豐富度在8種左右, 北岸物種豐富度變化幅度較大, 差異明顯, 表明西江南岸河岸帶植物種類較為豐富,這與物種多樣性指數的變化一致。
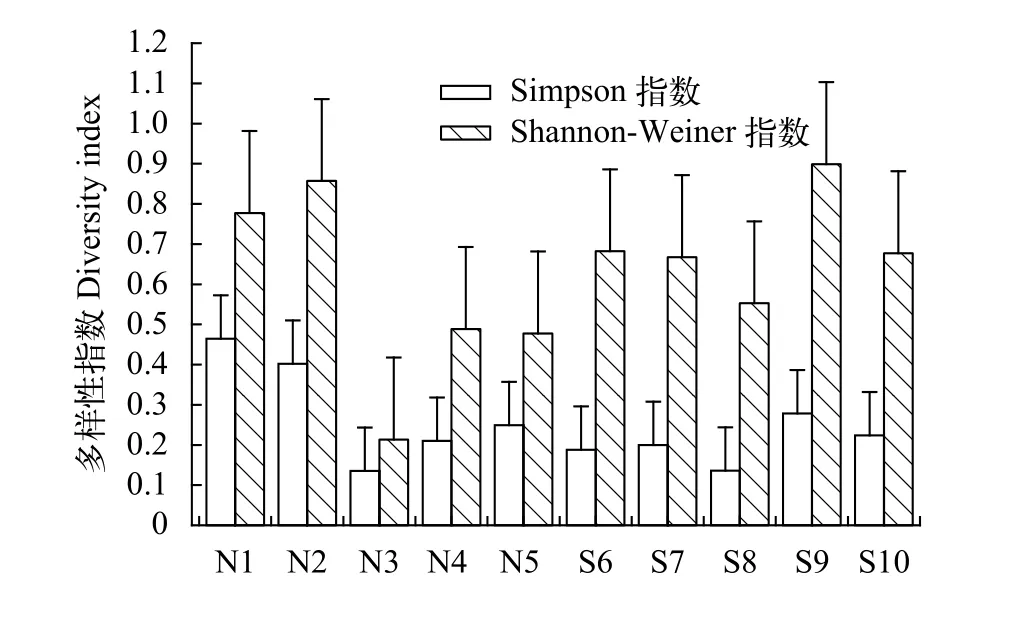
圖2 西江河岸帶各斷面多樣性指數變化(平均值±標準差)Fig. 2 Species diversity index in different transects in riparian zone of the Xijiang Oxbow (Mean±SD)
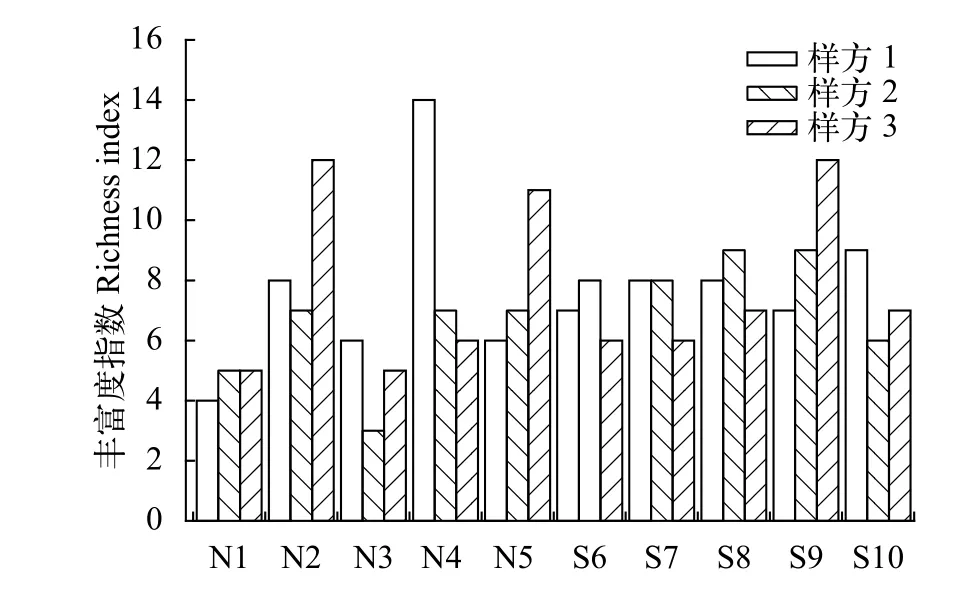
圖3 西江河岸帶各斷面物種豐富度指數變化Fig. 3 Species richness index in different transects in riparian zone of the Xijiang Oxbow
3 討論
3.1 西江河岸帶植物群落結構
西江河岸帶植物共有166種, 隸屬于50科131屬,被子植物占絕對優勢, 與長江天鵝洲故道相比[16],前者以蝶形花科、菊科、禾本科的物種較多, 后者以禾本科、菊科、蓼科的物種較多, 因此, 兩者形成的單優勢種群落都比較鮮明。在空間上, 西江河岸帶植物群落呈明顯的帶狀分布, 接近水域的主要是灰化苔草群落, 離水域較遠的主要是虉草群落和蘆葦群落, 隨著高程的增加逐漸轉變為蔞蒿群落、益母草群落、青蒿群落和狗牙根群落(圖4), 這類似于洞庭湖濕地植被空間分布特征[17]。從分布區類型上來看, 西江河岸帶植物科、屬的分布類型比較集中, 在植物科的水平上, 世界分布科的數量占優勢, 天鵝洲故道科的分布類型的數量多于西江;在植物屬的水平上, 世界分布和泛熱帶分布占較高的比例。對于植物屬的組成, 西江河岸帶植物屬的組成較為分散, 以寡種屬和單種屬為主, 與安徽省濕地植物屬的組成結構特征相似[18]。
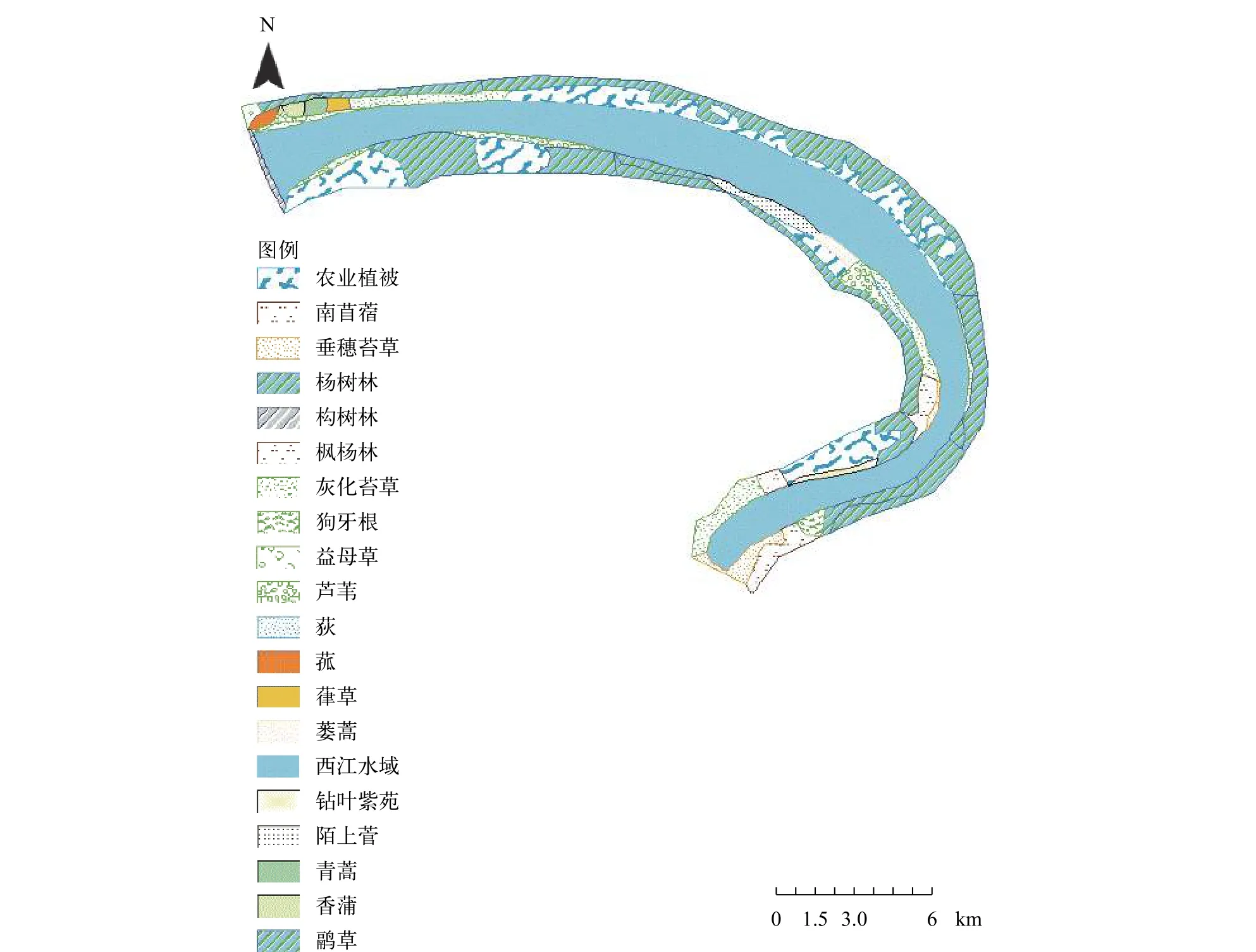
圖4 西江濕地植被分布圖Fig. 4 The distribution pattern of wetland vegetation in Xijiang Oxbow
3.2 西江河岸帶植物群落多樣性的影響因子
環境因子對植物的生長和分布具有不可忽視的作用, 主要包括氣候和地形、水文條件、土壤條件、泥沙淤積等非生物因子[19]。本研究主要從氣候和地形以及土壤因子兩方面進行分析。一方面,氣候和地形可通過調節植物可利用水分, 進而影響植物的分布。西江地勢中間高兩翼低, 氣候溫和,雨量適中, 為當地植物的定居和繁殖提供了優越的條件。曾有研究報道, 受距離水岸遠近及水面高低的影響, 河岸帶植物呈現縱向梯度的帶狀分布[20,21],與之類似, 西江河岸帶植物優勢種也表現出“水生-濕生”空間變化格局。
另一方面, 土壤也是影響河岸帶植物生長和分布的主要因子之一。已有研究報道, 河岸帶植物能充分利用土壤及周圍生態系統的營養元素, 也顯著影響了土壤中的氮、磷、有機碳等物質的含量[22,23]。還有學者對鄱陽湖濕地植被開展了相關研究, 發現植物分布主要受土壤含水量的影響, 其他土壤因子也是通過土壤含水量來影響植物分布的[24]。西江沿岸的土壤具有過渡性, 土壤中的氮元素分布不均勻, 但土壤有機物質易分解且逐年積累。本研究發現西江河岸帶植物多喜濕或水生, 且這些植物多為一年生或者兩年生, 可在短期內形成優勢種群, 可以很好地適應西江的土壤過渡性變化。

附表1 安慶西江河岸帶植物種類名錄Appendix Tab. 1 Species list of riparian vegetation on the bank of the Xijiang River in Anqing

續附表1
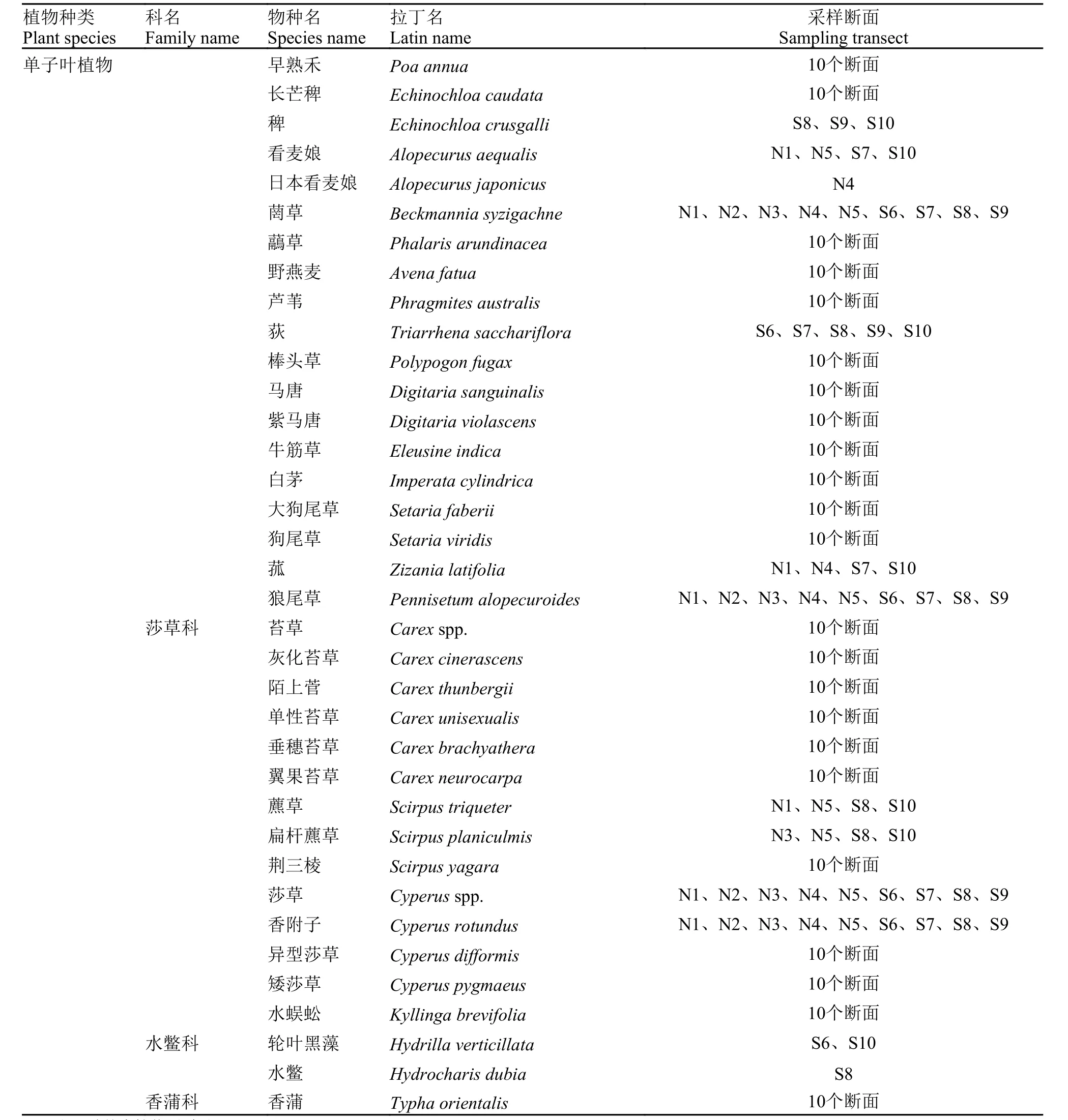
續附表1
除了自然環境, 人類活動同樣可能對河岸帶植物多樣性產生影響。西江河岸帶植物物種組成表明(附表1), 人類活動如農田開墾、人類沿河岸居住等活動影響了河岸帶群落的物種組成, 一些農田雜草(車前、獨行菜、薺菜等)、人類伴生植物(葎草、鬼針草、蒼耳等)伴隨著人類活動被帶入到河岸帶植物群落中。
由以上分析可知, 為了提升棲息地的質量, 必須盡可能維持西江河岸帶濕地的原始狀態, 以保持西江河岸帶植物群落和水域生態系統的穩定性。
3.3 西江河岸帶植物群落與長江江豚遷地種群棲息生境的關系
長江江豚作為河流生態系統的頂級生物, 其棲息和活動受魚類資源、水域環境、地形地貌和河岸固化等因素影響, 對棲息生境的要求較高, 且對其變化敏感[25,26]。現有報道顯示, 長江江豚喜歡棲居在長江干流近岸300 m的淺水水域[8,27], 并且經常在江河、江湖交匯處和沙洲附近等魚類資源較豐富的水域活動, 可能是因為這些水域河床坡度平緩,底質為淤泥, 水流緩慢, 水生生物資源豐富, 人類活動較少[8,28—31]。因此, 本研究重點分析河岸帶植物與水域環境和魚類資源互相影響的關系。
西江河岸帶植物種類豐富多樣, 植被覆蓋率高,這有利于改善西江水環境質量[32,33]。同時, 西江常水位下為封閉水體, 魚類群落以小型定居型物種占優, 多為產黏性卵魚類, 因此廣泛分布的河岸帶植被為西江魚類提供了優良的產卵場和索餌場[34], 對于西江魚類種群增長, 特別是適合于長江江豚捕食的小型魚類繁殖具有積極作用。既滿足了長江江豚生存對水體的要求, 也保證的長江江豚餌料魚的持續供應量。總體而言, 安慶西江為靜水環境, 水中泥沙含量低, 沿岸帶均為自然岸線, 且緩坡廣泛分布, 適宜多種河岸帶植物定居和繁殖, 也是長江江豚和其食物魚資源棲息所需要的水文和水生態環境。因此, 在保護區后續管理過程中, 應盡可能保持河岸帶自然化, 慎重考慮河岸帶植物演變可能帶來的影響, 及時監測和評估江豚保護區的環境變化。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
