不同抗性大麥品種對葉斑病的生理響應及主成分分析
2019-06-14 07:10:32黃志磊李葆春汪軍成姚立蓉司二靜孟亞雄馬小樂王化俊
麥類作物學報 2019年5期
黃志磊,李葆春,汪軍成,姚立蓉,司二靜,楊 軻,孟亞雄,馬小樂,王化俊
(1.甘肅省干旱生境作物學重點實驗室/甘肅省作物遺傳改良與種質創新重點實驗室,甘肅蘭州 730070;2.甘肅農業大學農學院,甘肅蘭州 730070;3.甘肅農業大學生命科學技術學院,甘肅蘭州 730070)
根據Statista數據顯示,截止2016年大麥在全球的種植面積和產量僅次于水稻、玉米和小麥,是第四大禾谷類作物[1],也是我國干旱和半干旱地區、氣候偏冷涼地區重要的種植作物。在大麥生長發育過程中,常受各種病菌侵害造成產量損失或者降低品質,大麥葉斑病(Spot blotch)是大麥生長發育過程中常發生病害之一,是由禾旋孢腔菌[Cochliobolussativus,無性世代(Bipolarissorokiniana)]引起的一種真菌性病害。其病原菌可以侵染大麥根部、葉片和幼胚,引起根腐病、葉斑病和黑胚病。該病害在美國、加拿大、南亞、南非和我國內蒙古東部以及東北地區發病嚴重,發病田塊減產幅度一般為20%。內蒙古呼倫貝爾市是我國三大啤酒大麥主產區之一,其大麥種植面積占內蒙古全區大麥面積的70%以上。近年來該地區大麥苗期根腐病及葉斑病危害嚴重,2012間大麥根腐病及葉斑病大爆發,導致大面積減產,根腐病和葉斑病已成為制約該地區大麥生產的重要病害之一[2]。目前還沒有防治該病害的有效方法。國外在大麥抗葉斑病鑒定、抗病遺傳資源利用等方面有較深入研究并取得重要進展。Lü等[3]用 14 479 菌株在苗期接種鑒定 205 份大麥種質資源,獲得高抗葉斑病材料11份;Gyawali等[4]探究了摩洛哥和印度春大麥苗期和成株期斑點病抗性與全基因組的關聯性;Burlakoti等[5]在美國中西部的六棱和二棱大麥育種中利用單核苷酸多態性(SNP)標記與全基因組關聯研究的方法確定了相關聯的10個抗性數量性狀基因座(QTL)。目前,國內外尚未發現對葉斑病完全免疫的大麥品種。我國對玉米、胡麻、花生、香蕉等作物葉斑病的研究已相當深入,但是對于大麥葉斑病菌研究甚少,尤其是就葉斑病菌對大麥生理特性的影響研究。郭煥強等[2]利用21個Bipolarissorokiniana菌株接種鑒定78個具有代表性的大麥主栽生產品種和骨干親本,并發現蒙啤麥3號的抗葉斑病能力強于蒙啤麥1號。植物的保護反應是復雜的新陳代謝過程,在受到病原物侵染后,抗、感品種之間PAL、PPO、POD、SOD以及其他重要酶類的活性變化有明顯差異[6-9]。研究表明,寄主體內的防御酶POD、PAL和SOD活性變化是水稻抗鏈霉菌JD211的重要原因[10]。Pro、SS和MDA是植物應對逆境脅迫時在細胞中積累的通用滲透調節物質,在遭受脅迫時可以通過在細胞內積累這些物質,以達到降低細胞水勢、維持細胞膜兩側滲透勢平衡來抵抗逆境脅迫的目的[11]。植物在受病原物侵染時,會產生一些蛋白(pathogensis-related proteins,PR蛋白)以抵御病原菌的進一步侵染,其中CHT和 β-1,3-葡聚糖酶是兩類重要的PR蛋白[12]。本研究在前人研究的基礎上以大麥品種蒙啤麥1號(感病)與蒙啤麥3號(抗病)為材料,了解在被葉斑病菌侵染后不同抗性品種組織器官中各項生理指標變化,以期為以后大麥抗大斑病的基因挖掘和育種工作提供 參考。
1 材料與方法
1.1 供試材料
供試大麥品種為蒙啤麥1號(感病)與蒙啤麥3號(抗病)。大麥葉斑菌樣選用甘肅農業大學麥類試驗室甘南Z14484強侵染力菌株。
1.2 試驗設計
試驗于2018年4-7月在甘肅農業大學人工氣候室進行。將土、沙、蛭石以3∶1∶1混合均勻,經121 ℃高壓滅菌30 min。大麥種子用5%次氯酸鈉溶液消毒30 min,然后用蒸餾水沖洗3次,每次5 min,繼續溫水浸種半小時。然后在培養皿中發芽,發芽后移栽到直徑 7 cm 的小花盆中,在 22 ℃的溫室中培養。待幼苗長到三葉期時,接菌植株用蒸餾水清洗葉片表面的蠟質層,最后將配置好的孢子懸浮液噴灑到葉片上,濃度為1×104個·mL-1(含0.25% Tween 20),一天噴施兩次,連續接種三天,接種期間溫室處于黑暗狀態,接種溫度為22 ℃,利用加濕器加濕,保持濕度100%;對照則是在相同條件下生長不接菌。于苗期、拔節期和孕穗期8:00采樣(基生葉),每個品種的接菌處理和對照均設3次重復,每個重復種4株幼苗。同時,調查大麥對病害的反應型。反應型采用常規九級劃分標準:1~3 級為高抗(R),4~5 級為中抗(MR),6~7 級為中感(MS),8~9 級為高感(S),用“+”和“-”表示實際反應型比相應各級標準反應型高或低。
1.3 生理指標測定
SOD活性采用NBT光還原法測定[13],POD活性采用愈創木酚法測定[13],CAT活性采用紫外吸收法測定[13],MDA含量采用硫代巴比妥酸法測定[13],游離脯氨酸(Pro)含量采用茚三酮法測定[13],PPO活性采用鄰苯二酚法測定[14],PAL活性采用苯丙氨酸脫氨法測定[14],LOX活性采用亞油酸鈉分光光度法測定[14],CHT活性采用二甲氨基苯甲醛法測定[14], β-1,3-葡聚糖酶活性采用3,5-二硝基水楊酸(DNS)法測定[14],SS含量采用蒽酮比色法測定[14]。
1.4 數據統計分析
數據的處理和統計分析分別采用WPS、EXCEL和SPSS軟件,采用Duncan's法進行多重比較(α=0.05)。
2 結果與分析
2.1 大麥品種的抗病性差異
接菌后,苗期、拔節期和孕穗期蒙啤麥1號反應型分別為2+、6、8,而蒙啤麥3號分別為1-、1、3,蒙啤麥3號上的菌斑生長速度與大小均弱于蒙啤麥1號(圖1),說明蒙啤麥3號對葉斑病抗性較蒙啤麥1號強,這與郭煥強等[15]的結論相符,也表明本研究的選材符合試驗要求。
2.2 接菌后大麥葉片抗氧化酶活性的變化
2.2.1 SOD活性變化
接菌后,蒙啤麥1號和蒙啤麥3號苗期和拔節期的葉片SOD活性較對照均顯著升高(圖2A),其中苗期增幅分別為5.4%和3.8%,拔節期分別為1.2%和11.1%;孕穗期,蒙啤麥1號SOD活性變化不顯著,蒙啤麥3號顯著升高,增幅為11.6%。
2.2.2 POD活性變化
接菌后,蒙啤麥1號與蒙啤麥3號的POD活性在三個時期較對照均有所提高,其中在拔節和孕穗期達到顯著水平(圖2B),蒙啤麥1號這兩個時期的增幅分別為24.6%和72.4%,而蒙啤麥3號分別為29.9%和152.8%。

圖1 接菌后蒙啤麥1號和蒙啤麥3號的表型觀察
2.2.3 CAT活性變化
接菌后,蒙啤麥1號與蒙啤麥3號的CAT活性在苗期較對照有所下降,在拔節期和孕穗期較對照均顯著升高(拔節期蒙啤麥1號除外)(圖2C),拔節期兩個品種的CAT增幅分別為10.4%和 52.6%,孕穗期分別為70.1%和65.7%。
2.2.4 PPO活性變化
接菌后,蒙啤麥1號與蒙啤麥3號三個時期的PPO活性較對照均增加(圖2D),其中蒙啤麥1號苗期變化不顯著,蒙啤麥3號變化顯著(增幅為5.3%),在其余兩個時期兩個品種均變化顯著。在拔節期,兩個品種的增幅分別為47.0%和86.8%,孕穗期分別為88.7%和183.8%。
2.2.5 LOX活性變化
接菌后,蒙啤麥1號與蒙啤麥3號的LOX活性在三個時期均較對照升高,其中拔節期和孕穗期的接種處理與對照差異均顯著,兩個品種的增幅在拔節期分別為31.1%和58.4%,孕穗期分別為11.9%和22.6%。

MP1:蒙啤麥1號;MP3:蒙啤麥3號。圖柱上的不同字母表示每個時期處理間在0.05水平上差異顯著。下圖同。
MP1:Mengpimai 1;MP3:Mengpimai 3.Different letters above the columns are significantly different between the treatments at the 0.05 probability level at same stage. The same in other figures.
圖2 接菌后蒙啤麥1號和蒙啤麥3號抗氧化酶活性的變化
Fig.2 Activity changes of the antioxidant enzymes of Mengpimai 1 and Mengpimai 3 inoculated leaf spot pathogen
2.3 接菌后大麥葉片Pro、SS和MDA含量的變化
接菌后,蒙啤麥1號和蒙啤麥3號的Pro、SS和MDA含量在三個時期均不同程度增加,而且在大部分情況下,接菌處理與對照的差異均達到顯著水平。其中,兩個品種的Pro含量增幅在拔節期分別為96.2%和159.2%,在孕穗期分別為 250.2%和280.7%;SS含量的拔節期增幅分別為48.0%和43.4%,孕穗期分別為43.1%和164.7%;MDA含量的苗期增幅分別為10.9%和21.0%,拔節期增幅分別為8.5%和34.2%。
2.4 接菌后大麥葉片抗病相關酶活性的變化
接菌后,兩個品種的β-1,3-葡聚糖酶和PAL活性在苗期、拔節期和孕穗期均較對照顯著增加(圖4A和圖4C)。其中,蒙啤麥1號的 β-1,3-葡聚糖酶活性在三個時期的增幅分別為165.4%、161.1%和155.9%,蒙啤麥3號分別為268.9%、243.2%和95.5%;蒙啤麥1號的PAL活性增幅在三個時期分別為141.4%、48.5%和208.1%,蒙啤麥3號分別為14.1%、76.7%和151.9%。兩個品種的CHT活性在接菌后變化較小(圖4B),只有蒙啤麥1號在拔節期變化顯著,增加21.1%。
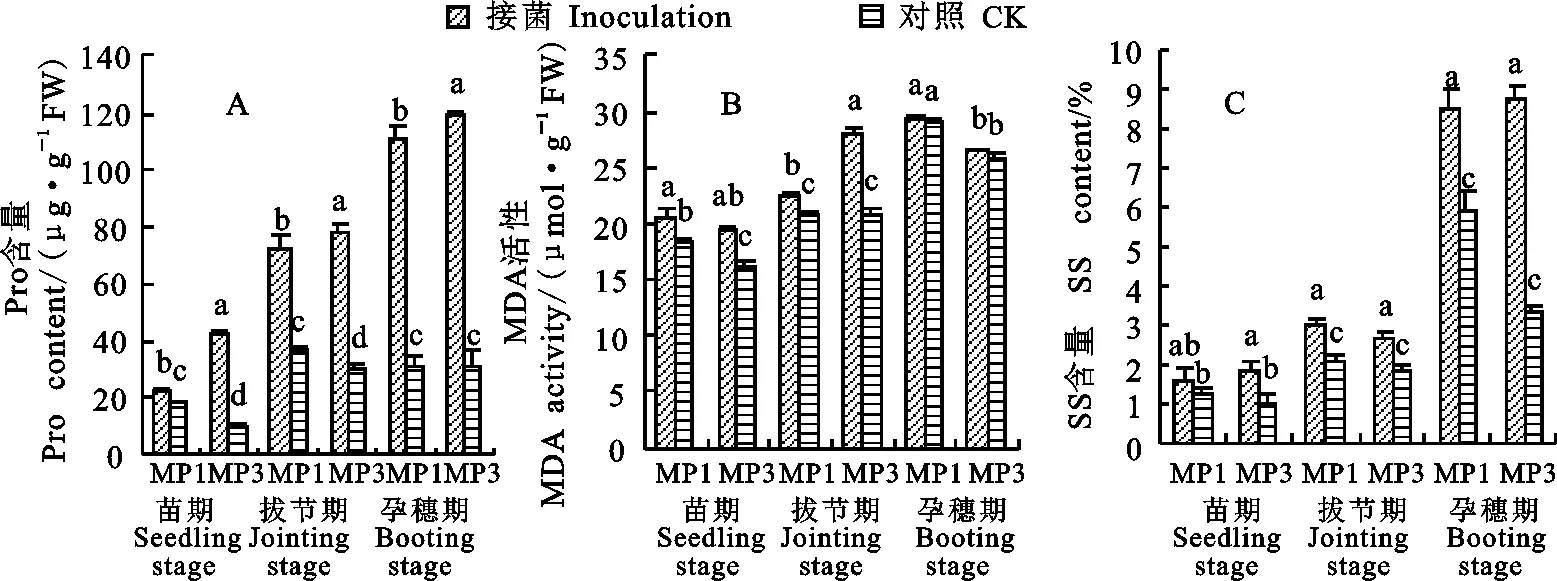
圖3 接菌后蒙啤麥1號和蒙啤麥3號Pro、SS和MDA含量的變化

圖4 接菌后蒙啤麥1號和蒙啤麥3號其他相關酶的變化
2.5 主成分分析
從以上分析結果可以看出,接菌后兩個品種的11個指標在不同時期的變化不同。為了得到更準確結果,本研究對數據進一步進行了主成分分析。結果表明,特征值大于1的主成分有四個,且這四個成分的方差貢獻率和達到了 96.313%(表1和圖5),說明這四個主成分囊括了11個生理指標所含的絕大部分信息。其中,第一主成分主要包括了SS含量、POD活性和PAL活性的信息,第二主成分主要包括了 β-1,3-葡聚糖酶活性的信息,第三主成分主要包括了LOX活性的信息;第四主成分主要包括了PPO活性的信息。以上信息表明,可選用SS含量、POD活性、PAL活性、 β-1,3-葡聚糖酶活性、LOX活性和PPO活性6個指標來評價大麥的抗葉斑病能力。表2列出了兩個品種三個時期11 個生理指標的主成分載荷。
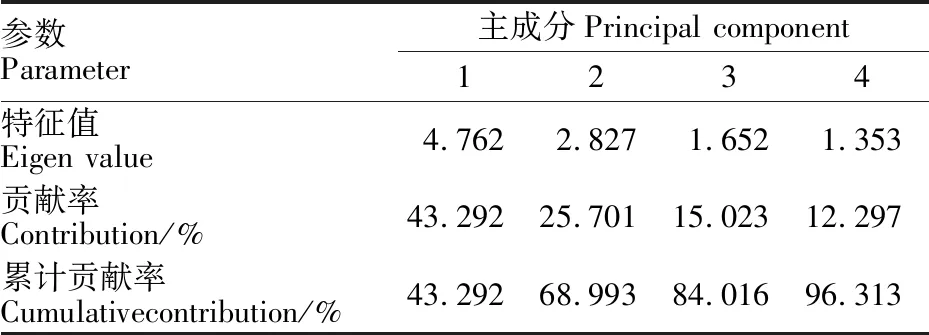
表1 大麥各生理指標對抗病性影響的主成分方差貢獻率Table 1 Variance of principal component variance of physiological indices of barley against disease
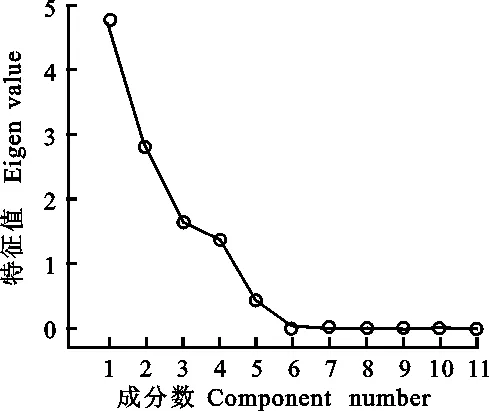
圖5 主成分的碎石圖
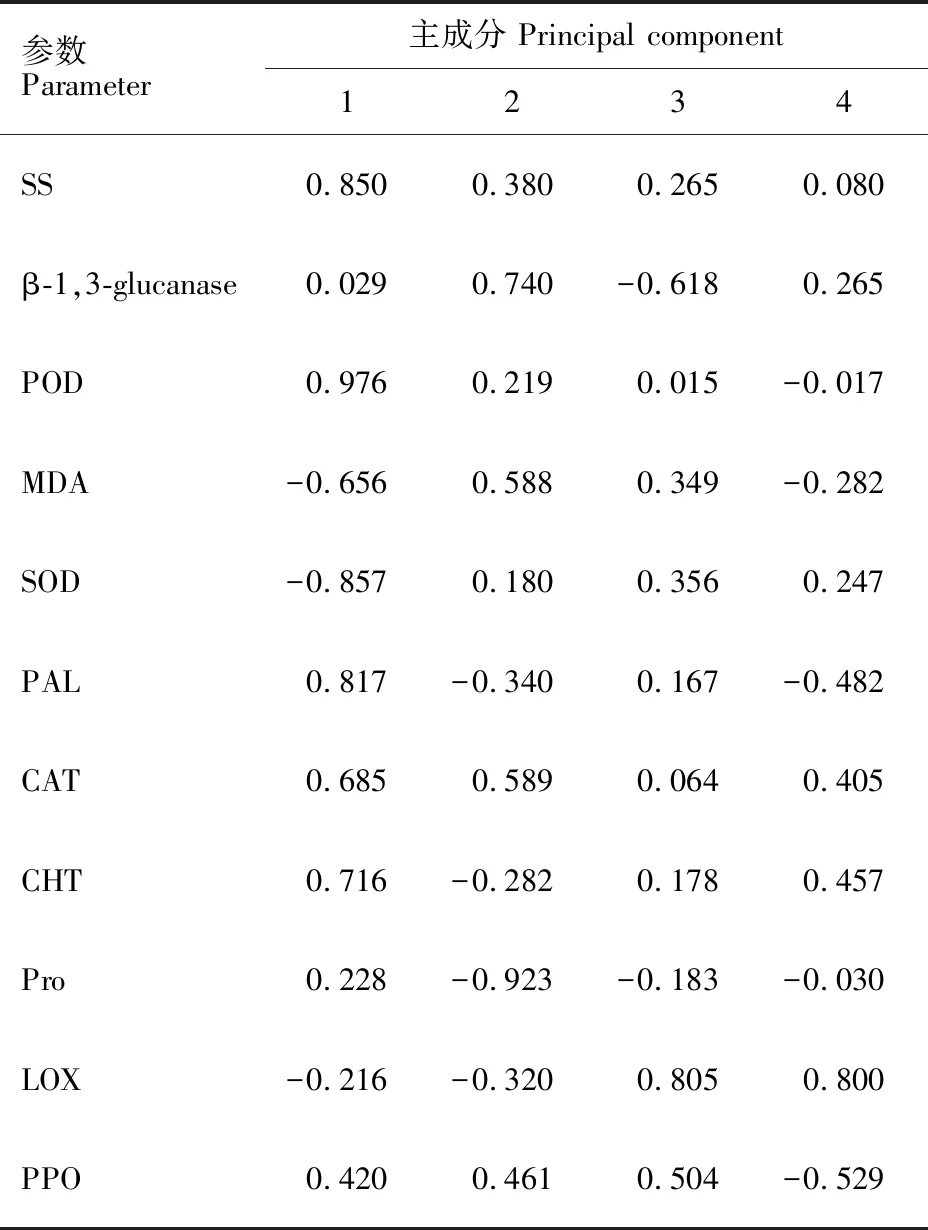
參數Parameter主成分 Principal component1234SS0.8500.3800.2650.080 β-1,3-glucanase0.0290.740-0.6180.265POD0.9760.2190.015-0.017MDA-0.6560.5880.349-0.282SOD-0.8570.1800.3560.247PAL0.817-0.3400.167-0.482CAT0.6850.5890.0640.405CHT0.716-0.2820.1780.457Pro0.228-0.923-0.183-0.030LOX-0.216-0.3200.8050.800PPO0.4200.4610.504-0.529
3 討 論
植物的抗逆性與其體內的酶類活性和化合物含量關系密切。ROS是植物體內一種重要的信號分子,但它同時也會對植物造成毒害,尤其在逆境中ROS迅速富集。因此,植物在長期適應過程中,為了響應和防御外界不良環境造成的氧化脅迫,植物形成了自己的一套調節及解毒機制,可分為回避機制和清除機制。其中清除機制包括酶促清除,如SOD、CAT、POD等,以及非酶類清除系統,包括細胞色素C、谷胱甘肽、甘露醇、抗壞血酸、不飽和脂肪酸等[19]。本試驗中,蒙啤麥1號和蒙啤麥3號在受到葉斑病菌侵害時,所測的氧化還原類酶活性都發生改變。隨著脅迫時間的增長,病害加劇,SOD、POD、LOX和PPO活性升高幅度逐漸增加,且抗病品種蒙啤麥3號升高幅度均大于感病品種蒙啤麥1號,這與汪寶卿等[20]的研究結果一致。接菌處理CAT活性較對照先降后升,可能是由于品種自身特性決定的。通過主成分分析,POD、LOX與PPO活性在抗葉斑病方面貢獻率最大,可為以后抗、感品種的篩選與鑒定提供參考。
植物在長期的進化過程中發展出了一系列細胞內源通用非生物脅迫應答機制,通過應激代謝反應在細胞內大量積累一些有機小分子化合物和多肽類物質(Pro等)來降低原生質水勢,調節逆境脅迫條件下細胞膜兩側滲透勢平衡,以有效應對不利環境條件對植物生理代謝系統造成的損害[21]。MDA是膜脂過氧化的分解產物,其含量的高低可反映植物受到損傷的程度。因此植物的Pro、SS和MDA含量也通常被視為衡量不同植物對非生物脅迫耐受能力的關鍵指標。本研究中,供試材料的Pro、MDA、SS 含量在染菌后均不同程度增加,這與張雯莉等[22]的研究結果相同。通過主成分分析發現,蒙啤麥1號與蒙啤麥3號對葉斑病在滲透調節方面的抵御主要通過SS的積累來實現。
PAL參與木質素與植保素的合成,可以增加細胞壁的厚度,提高組織木質化程度,形成病原菌入侵的物理屏障[23]。高等植物體內廣泛存在著 β-1,3-葡聚糖酶和CHT。葡聚糖和幾丁質是真菌細胞壁的主要成分,而大多數真菌的菌絲頂端葡聚糖酶和幾丁質暴露在表面,能夠直接受到 β-1,3-葡聚糖酶和CHT的作用,從而刺激寄主植物的抗病反應,而且CHT和 β-1,3-葡聚糖酶協同作用更能抑制病原菌的侵害[24]。本試驗也再次證實了該結論。在所測病害相關酶類中,接菌后PAL和 β-1,3-葡聚糖酶活性均呈上升趨勢,其中 β-1,3-葡聚糖酶與PAL的貢獻率最高,起主導作用,此結果與龍艷玲等[25]研究結論一致。
麥根腐平臍蠕胞菌(B.sorokiniana)是兼性寄生真菌[26]。大麥抗葉斑病品種蒙啤麥3號、感葉斑病品種蒙啤麥1號在受病菌侵染后各項生理指標均發生變化。從所測11個生理指標綜合表現看,抗病品種蒙啤麥3號本身的自我調節能力和抵御機制明顯強于感病品種蒙啤麥1號。但是,CAT的苗期活性變化與前人研究不一致,可能是由于大麥在各生育時期的抗性會發生改變造成[27]。如小麥、大麥和燕麥對白粉、銹病以及水稻對白葉枯病等病害的抗性表現,其抗性從苗期開始表達,會持續至整個生育期,但苗期抗性與全生育期抗性也會存在不一致的情況,在苗期表現為感病,但在成株期卻具有高水平抗性,或苗期抗病,在成株期卻表現為感病[28]。可見,不同抗性品種在不同時期的生理特性存在差異,染菌條件下,POD活性、LOX活性、PPO活性、SS含量、PAL活性與 β-1,3-葡聚糖酶活性可作為篩選抗葉斑病大麥的參考指標。
