RNA-Seq定量分析鹽膚木對鉛脅迫的響應
2019-07-19 07:22:48夏麗丹胡華英周垂帆
農業環境科學學報 2019年7期
關鍵詞:植物
夏麗丹 ,張 虹 ,胡華英 ,曹 升 ,周垂帆 *
(1.福建農林大學林學院,福州 350002;2.海峽兩岸水土保持協同創新中心,福州 350002;3.福建長汀紅壤丘陵生態系統國家定位觀測研究站,福州 350002)
鉛是一種有毒的重金屬,采礦、印染、電鍍等行業排放的廢棄物中含有大量的鉛,這使得土壤環境污染嚴重,且鉛具有強累積性,可通過食物鏈進入人體,與人體中的多種酶發生反應,損傷人的免疫、消化、神經與生殖系統,從而威脅人體健康[1]。
鉛不是植物生長的必需元素,過量的鉛會對植物產生毒害效應:葉片小、發育不良、生物量減少、葉綠素含量降低、光合作用下降等[2-4]。但植物在長期的進化過程中會形成特定的各種適應性的生長習性和生理、形態特征[5],一些植物則會表現出較強的重金屬耐性。揭示植物適應鉛脅迫的分子機制,有利于抗性植物的培育,從而發揮植物修復物種的最大優勢。目前的研究已發現并推測出眾多參與重金屬響應的基因,其中以與重金屬運輸相關的蛋白基因的研究最為廣泛,主要包括HMAs(重金屬ATP酶)家族,ABC(ATP結合盒)轉運家族、CDF(陽離子擴散促進者)家族和ZIP(鋅/鐵調節轉運蛋白)家族等,這些蛋白基因在提高植物對重金屬的耐性上起到了一定的作用。此外,還有一些基因家族對重金屬離子在細胞中的運輸及提高植物的抗性等有重要作用,主要有YSL(黃色條紋樣)蛋白家族、Nramps(天然抗性相關巨噬細胞蛋白)家族與CTR(銅離子轉運蛋白)家族等[6-7]。現已有40多種木本植物完成全基因組測序工作,但大多集中于葡萄、石榴等可食用果樹[8],而對重金屬耐性植物的研究較少。
鹽膚木(Rhus chinensis)是我國主要經濟樹種之一,具有良好的經濟與藥用價值。調查研究發現,鹽膚木是重金屬污染嚴重礦區為數不多的能夠自然定居的木本植物,生物量較高、生長快、適應性強、根系發達、根萌蘗性強,對鉛、鉻等重金屬表現出較強的耐性,因此可作為南方重金屬污染區生態修復的先鋒植物[4,9-10]。但目前關于鹽膚木重金屬脅迫的研究主要集中于重金屬污染脅迫下的生理學響應及光譜特征[11]、遷移機制及富集特征[4]等,而有關鹽膚木重金屬脅迫下基因響應機制的研究鮮有報道。這極大制約了我們對鹽膚木蛋白基因在重金屬耐性方面的認知。目前,RNA-Seq技術是以新一代高通量測序為平臺的RNA測序技術,該技術能夠在單核苷酸水平上對任一物種的整體轉錄情況進行檢測,能夠更精確、更廣泛地提供物種在特定狀態下的基因表達情況,是深入研究復雜轉錄組的強大工具[12-14]。
鑒于此,本研究利用Illumina HiSeqTM2000高通量轉錄組測序技術對鹽膚木鉛脅迫下的根系組織進行轉錄組測序,并對差異基因進行Gene Ontology(GO)與Pathway分析,試圖揭示鹽膚木響應鉛脅迫的相關分子機理,以期為鹽膚木應用于南方土壤重金屬污染修復提供理論依據。
1 材料與方法
1.1材料的種植與處理
本研究以鹽膚木為供試植物。取大小均一、飽滿的鹽膚木種子,在98%濃硫酸中浸泡消毒90~105 min,每隔10 min用玻璃棒攪拌一次。消毒完成后,用流水清洗種子,搓去種子表面黑色蠟質,并用純水浸泡24 h,而后洗凈置于濕濾紙上培養,每2 d更換一次濾紙,并移除發霉和空殼的種子。該發芽過程在20±2℃人工氣候培養箱中避光進行。待芽長至約2 cm,將其移至營養土中栽培,待植株長至20 cm左右,取長勢較為一致的幼苗進行土培脅迫試驗。
脅迫試驗土壤取自福建農林大學后山紅黃壤,參考魯如坤[15]的土壤元素測定方法,測得土壤背景值,結果見表1。試驗用土1.5 kg·盆-1,每盆種植1株,重復3次。設置土壤鉛濃度為 0(CK)、250(Pb250)、1000(Pb1000)mg·kg-1(實測值分別為0、278.843 、1 103.498 mg·kg-1),鉛以 Pb(NO3)2溶液的形式加入土壤中,用純水澆灌保持土壤濕度為40%~50%。土壤平衡2~3d后,將鹽膚木植株移栽到供試土壤中(福建農林大學下安溫室大棚),溫度為25℃左右,脅迫一個月后取根進行測序試驗。

表1 土壤背景值Table 1 Soil background value
1.2 RNA提取與測序
總RNA的提取采用Trizol法[16],由廣州基迪奧生物公司完成RNA的提取、質控、文庫構建與測序。
1.3 基因功能表達、注釋及富集分析
1.3.1 Unigene注釋
Unigene基本功能注釋信息給出Unigene的蛋白功能注釋、COG/KOG功能注釋、GO功能注釋、Path?way注釋等。通過blastx將Unigene序列比對到蛋白數據庫Nr、Swissprot、KEGG(系統分析基因產物在細胞中的代謝途徑以及這些基因產物功能的數據庫)和COG/KOG(e-value<0.000 01)(基因產物進行直系同源分類的數據庫),得到與給定Unigene具有最高序列相似性的蛋白,從而得到該Unigene的蛋白功能注釋信息。
1.3.2 GO功能顯著性富集分析
GO是一個國際標準化的基因功能分類體系,提供了一套動態更新的標準詞匯表來全面描述生物體中基因和基因產物的屬性。GO功能分析一方面給出差異表達基因的GO功能分類注釋;另一方面給出差異表達基因的GO功能顯著性富集分析,通過GO功能顯著性富集分析能確定差異表達基因行使的主要生物學功能。計算得到的p-value通過FDR[17]校正之后,以q-value≤0.05為閾值,滿足此條件的GO term定義為在鉛脅迫下樣品間差異表達基因中顯著富集的GO term。
1.3.3 Pathway功能顯著性富集分析
在生物體內,不同基因相互協調行使其生物學特性,基于Pathway的分析有助于更進一步了解基因的生物學功能。KEGG是有關Pathway的主要公共數據庫[18]。Pathway顯著性富集分析以KEGG Pathway為單位,應用超幾何檢驗,找出與整個基因組背景相比,在差異表達基因中顯著性富集的Pathway。通過Pathway顯著性富集分析能確定差異表達基因參與的最主要生化代謝途徑和信號轉導途徑。通過計算以q-value≤0.05為閾值篩選出樣品間差異基因的主要代謝路徑。
2 結果與分析
2.1 樣品測序質量與組裝結果
使用短reads比對軟件Bowtie 2[19]將高質量clean reads比對參考基因序列(表2),CK、Pb250、Pb1000與參考基因的匹配率分別為85.69%、84.58%、84.56%,表明測序數據可用性較高。使用短reads組裝軟件Trinity[20]做轉錄組從頭組裝(表3)發現,轉錄本拼接長度大多集中在200~299 nt,轉錄本長度介于201~15 581 nt,N50值為1566,拼裝組裝效果符合要求。

表2 各樣品測序質量統計Table 2 Sampling quality statistics of each sample

表3 組裝結果統計Table 3 Assembly result statistics
2.2 Unigene基本注釋統計
2.2.1 數據庫注釋統計
如圖1所示,鹽膚木70 459個Unigene與KEGG、KOG、Nr與Swissprot四大數據庫分別比對得到20 363、23 655、41 047、34 476個 Unigene,共有 43 006個Unigene得到注釋,27 453個Unigene未得到注釋,說明鹽膚木轉錄組中還存在大量未知基因,而有14 087個Unigene在四大數據庫中均得到注釋,占比32.76%。
2.2.2 物種分布統計
利用blastx將組裝出來的Unigene序列與Nr數據庫進行比對,有9927個Unigene與柑橘、3509個Unigene與可可、1563個Unigene與麻風樹等植物的序列同源(圖2)。這些基因為本研究轉錄組的注釋提供了豐富的參考序列,并且從這些數據中可以推斷出鹽膚木與柑橘的進化關系較近。
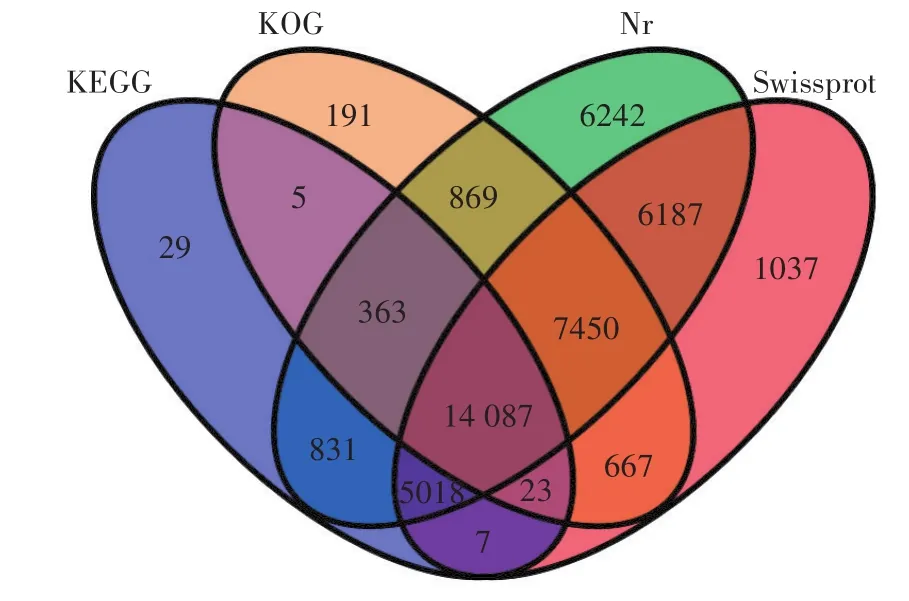
圖1 四大數據庫注釋維恩圖Figure 1 Four big database annotations Venn diagram
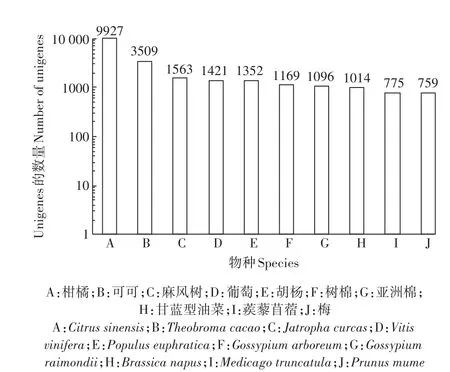
圖2 物種分布統計圖Figure 2 Species distribution chart
2.3差異基因數量統計
對3個樣品間的差異基因進行兩兩比較,用FDR與log2FC來篩選差異基因,篩選條件為FDR<0.05且|log2FC|>1,結果見圖3。
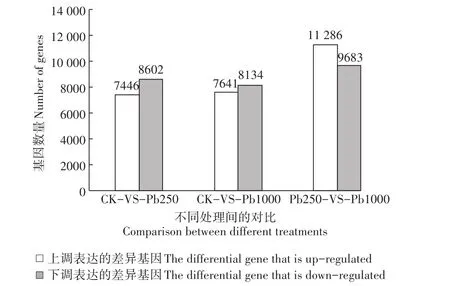
圖3 樣品間差異基因數量統計Figure 3 Quantitative statistics of differential genes between samples
在本研究中,CK-VS-Pb250、CK-VS-Pb1000的上調基因表達量分別為7446、7641個,下調基因分別為8602、8134個。相對于CK而言,Pb1000的上調基因比Pb250增加了195個,表明鹽膚木在較高鉛濃度下一些平常不表達的基因開始表達,這與其抵御鉛脅迫的耐性相關[21];Pb1000的下調基因比Pb250降低了468個,這可能是因為鉛脅迫對鹽膚木的生理系統造成了傷害,使其活性降低,基因表達量下降[22]。Pb250-VS-Pb1000的上調、下調基因分別為11 286、9683個,皆大于前兩者,表明鹽膚木在遭受高濃度鉛脅迫時,雖然大量基因表達量降低,但其自身會調用更多不常用的基因來應對逆境。
2.4 差異基因的GO分析
GO共有3個本體,分別描述基因的分子功能(Molecular function)、細胞組分(Cellular component)及參與的生物過程(Biological process)。鹽膚木鉛脅迫轉錄組基因注釋結果如圖4和圖5所示。
CK-VS-Pb250 GO分類統計見圖4:在分子功能分類中,共有12個差異表達基因顯著富集,分別是結構分子活性、2類RNA結合相關基因、4類水解酶相關基因、2類磷酸酶相關基因、2類聚合酶相關基因和西格瑪因子活動;在生物過程分類中,共有34個差異表達基因顯著富集,分別是基因表達、電子傳遞鏈、氧化還原過程、光合電子傳遞鏈、RNA修飾、細胞離子穩態、核糖體、17類代謝過程相關基因、5類合成過程相關基因、3類復合物相關基因和2類光合作用相關基因;在細胞組分分類中,共有24個差異表達基因顯著富集,分別是膜蛋白復合物、3類核糖體亞基相關基因、4類細胞器相關基因、3類細胞質相關基因、4類細胞及細胞器相關基因和9類光合作用相關基因。
CK-VS-Pb1000 GO分類統計見圖5:在分子功能分類中,共有11個差異表達基因顯著富集,分別是結構分子活性、四吡咯結合、6類氧化還原酶相關基因、單加氧酶活性、脂肪酶激活劑活性和rRNA結合;在生物過程分類中,共有2個差異表達基因顯著富集,分別是基因表達和碳水化合物衍生物分解代謝過程;在細胞組分分類中,共有16個差異表達基因顯著富集,分別是3類復合物相關基因、4類核糖體及核糖體亞基相關基因、2類細胞器相關基因、4類細胞相關基因和3類光合作用相關基因。
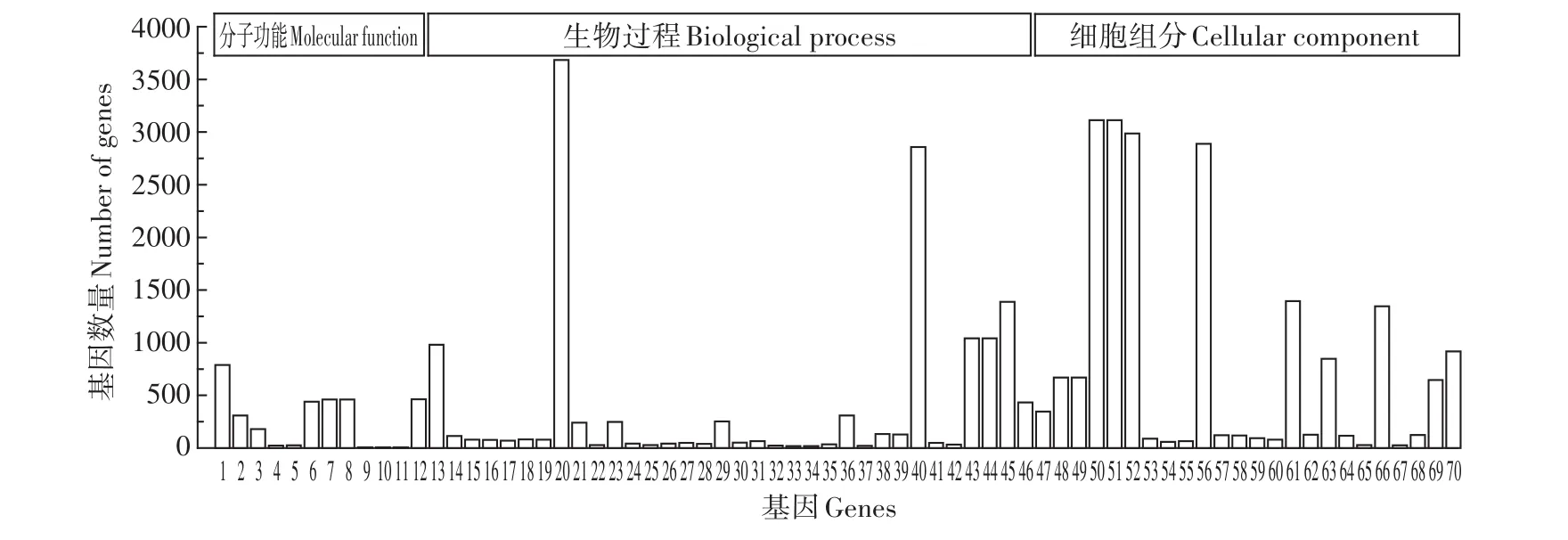
圖4 CK-VS-Pb250差異基因GO功能注釋Figure 4 CK-VS-Pb250 differential gene GO function annotation
由圖4和圖5可得,在分子功能上,Pb250脅迫下磷酸酶與水解酶相關基因表達量較高,而Pb1000脅迫下則是氧化還原酶,此外,四吡咯結合相關基因也有較高的基因表達量。在生物過程上,Pb250脅迫下富集了34個差異基因,而Pb1000僅有2個,表明植物受損嚴重,基因表達量驟降;Pb250富集量最多的差異基因是代謝過程,主要是有機物質的代謝,其中核糖體蛋白相關基因居多,如40S核糖體S3-3樣蛋白、60S核糖體蛋白L7、核糖體蛋白L11等。在細胞組分上,Pb250脅迫下主要是細胞結構相關基因富集較多,也存在較少數量的核糖體亞基,而Pb1000下核糖體相關基因出現大量富集,主要為核糖體相關亞基與核糖核蛋白復合物。
2.5 差異基因的KEGG分析
由表4可得,核糖體是CK-VS-Pb250、CK-VSPb1000與Pb250-VS-Pb1000差異樣品中共有的代謝通路,表明核糖體是鹽膚木適應鉛脅迫的主要代謝通路。在CK-VS-Pb250與CK-VS-Pb1000中分別有1211、1026個核糖體DEGs,表明高濃度鉛脅迫使得核糖體相關基因表達量減少。光合作用代謝通路僅存在于CK-VS-Pb250中,表明輕度鉛脅迫下,鹽膚木仍能有效進行光合作用,而重度鉛脅迫使得植物光合作用受到抑制,植物生長受到影響。此外,光合天線蛋白代謝通路的差異表達基因主要有葉綠素a/b結合蛋白Ⅱ型前體、葉綠素a/b結合13、葉綠素a/b結合蛋白3、預測葉綠素a/b結合蛋白CP24 10B等與葉綠素相關的基因。與CK相比,Pb1000較Pb250的光合天線蛋白通路減少了2個DEGs,分別是葉綠素a/b結合蛋白和預測葉綠素a/b結合蛋白1D樣。
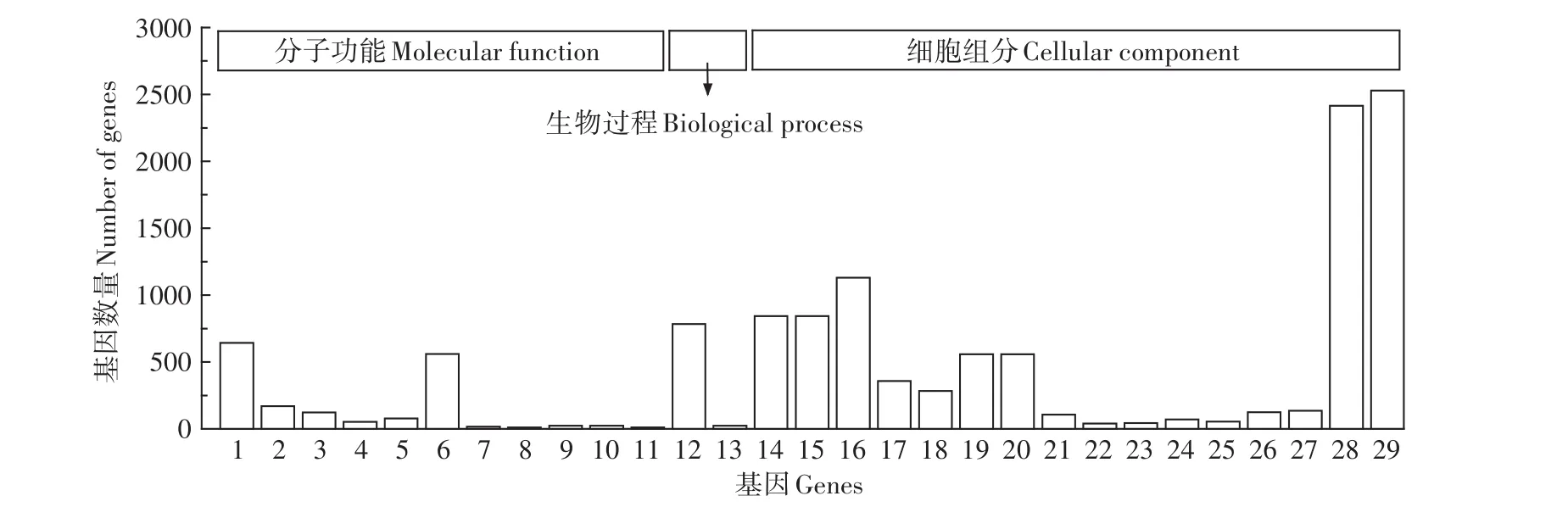
圖5 CK-VS-Pb1000差異基因GO功能注釋Figure 5 CK-VS-Pb1000 differential gene GO function annotation
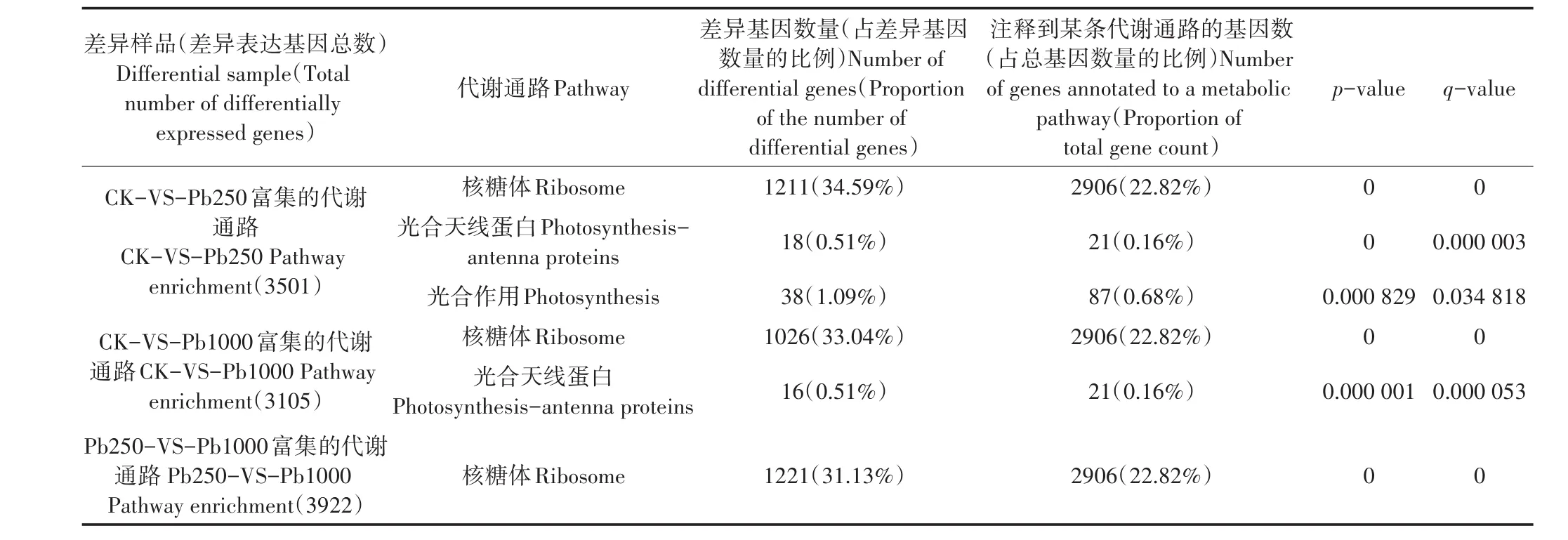
表4 差異表達基因的KEGG代謝途徑分析Table 4 Analysis of KEGG metabolic pathways of differentially expressed genes
3 討論
水解酶(XTH)是植物細胞壁重構過程中的關鍵酶之一,不僅具有松弛細胞壁、促進細胞生長的作用,而且也具有強化細胞壁和維持細胞壁完整性的作用[23-24]。在輕度鉛脅迫下有4類水解酶相關基因出現顯著富集,表明鹽膚木在輕度鉛脅迫下通過提高細胞中水解酶活性修復受損細胞壁。同時,在擬南芥和番茄中發現,超量表達CaXTH3能夠提高轉基因植株抗旱性和耐鹽性[25-26]。核苷三磷酸酶通過提供能量來促進核內成熟mRNA穿過核孔復合體進入胞漿,是細胞核mRNA出核轉運的主要限速酶[27];焦磷酸鹽是控制細胞內焦磷酸濃度的關鍵酶,而焦磷酸是RNA、DNA、蛋白質及糖原等生物大分子生物學合成過程中的副產物,其濃度會影響細胞內部分生理反應的平衡[28]。在輕度鉛脅迫下這兩類磷酸酶相關基因出現顯著富集,表明鹽膚木在輕度鉛脅迫下通過提高相關磷酸酶活性來調節細胞內被擾亂的平衡,以維持細胞內的正常運作。同時,Park等[29]將擬南芥H+-PPase基因轉入番茄中發現,轉基因植株與野生型植株相比生長速度快、根系發達,且有較強的抗旱能力。在高濃度鉛脅迫下,鹽膚木的各類氧化還原酶活性顯著增強,氧化還原酶能夠進行大量的催化反應,生物合成結構復雜的化合物[30],以抵御鉛侵害。此外,四吡咯化合物是植物光合作用、呼吸作用等生物學過程不可或缺的重要組分,維系著植物的生長和發育,當四吡咯合成途徑受阻時,一些光敏型中間代謝物積累,則會產生氧化脅迫,誘導細胞死亡,嚴重阻礙植物生長[31]。在重度鉛脅迫下四吡咯結合相關基因出現顯著富集,表明高濃度鉛脅迫已對四吡咯化合物的結合產生阻礙,影響了植物正常生長。施翔等[4]和Souza等[32]發現,高濃度重金屬脅迫下植物生物量降低。綜上,通過不同濃度鉛脅迫發現,細胞內水解酶、磷酸酶等相關基因在輕度鉛脅迫下主要通過提高自身活性來增強細胞抗性,而在重度鉛脅迫下,則是氧化還原酶相關基因通過合成各類化合物以抵御逆境。
中心體是非膜細胞器之一,是細胞中的微管組織中心,在提供細胞器定向運輸支架和細胞運動調節中發揮重要作用[33]。此外,核糖體也屬于非膜細胞器,本研究發現,無論是在輕度還是重度鉛脅迫下,非膜細胞器相關基因都出現大量富集,表明鹽膚木在鉛脅迫下細胞受損嚴重,而其自身首先通過細胞器調節相關的細胞運動以應對逆境。細胞質是生命活動的主要場所,絕大多數的化學反應都在細胞質中進行,主要成分為核糖體、多種酶類和中間代謝物、各種營養物等。在輕度鉛脅迫下有3類細胞質相關基因出現顯著富集,表明鹽膚木在遭受輕度鉛脅迫時,通過調節自身基因抵御侵害,如提高酶類基因表達、增強代謝等。Yao等[34]發現在番茄與蘋果愈傷組織中過量表達細胞質蘋果脫氫酸酶基因,不僅能夠促進細胞發育和植物生長,促進質子泵相關基因的表達和三磷酸腺苷酶的生成,而且能夠調節有機酸轉運和溶質勢,減少ROS的生成,從而提高轉基因作物對寒冷和鹽脅迫的抗性。綜上,鉛的侵入使得鹽膚木根系細胞受損嚴重,而鹽膚木會通過調節細胞內細胞器、細胞質等相關的細胞運動以應對逆境。
從差異表達基因的GO與KEGG的分析中均發現,核糖體相關基因是鹽膚木應對鉛脅迫的主要調節基因。核糖體蛋白不僅參與了rRNA的加工、折疊、核糖體亞基組裝和轉運過程,還在亞基結構的穩定性、核糖體與各種翻譯因子的相互作用和新生肽的折疊與定位等過程中發揮作用,甚至還可能承擔著核糖體外的生物學功能[35-37]。核糖體由rRNA和蛋白質構成,是細胞內蛋白質合成的分子機器,如果核糖體不起作用,細胞就無法分裂,植物生長將停止。與CK相比,Pb1000較Pb250核糖體通路減少了185個DEGs,因此推測高濃度鉛脅迫影響鹽膚木根系的正常代謝,抑制植株正常生長,通過對根系生理特性如SOD、CAT等抗氧化酶和MDA的測定分析也印證了這一點[38]。此外,光合作用代謝通路中的捕光葉綠素a/b結合蛋白等能夠促進葉綠體合成輔酶并提高光能利用率[39],表明在高濃度鉛脅迫下,光合作用受到一定的抑制,植物生長受到影響,筆者在對葉綠素含量的研究分析中也發現鉛顯著抑制了鹽膚木葉綠素含量[38]。姚廣等[40]也發現,鉛脅迫顯著抑制了玉米地上部分和地下部分的生長、降低了葉片光合色素含量,杜連彩[41]也有相似的研究結果。綜上,核糖體相關基因是鹽膚木應對鉛脅迫的主要應答與調節基因。
4 結論
(1)輕度鉛脅迫下,鹽膚木通過調節細胞內水解酶與磷酸酶相關基因抵御脅迫,而在重度鉛脅迫下,則是氧化還原酶相關基因起到主要調節作用。
(2)鹽膚木在鉛脅迫下細胞受損,其自身通過調節細胞內細胞器、細胞質等相關的細胞運動以應對逆境。
(3)核糖體代謝通路是鹽膚木適應鉛脅迫的主要代謝通路,核糖體相關基因是鹽膚木應對鉛脅迫的主要調節基因。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
