飼料魚溶漿影響黃顙魚的膽汁酸代謝及脂肪沉積
2019-07-20 08:08:08高敏敏吳代武周露陽葉元土蔡春芳萍郁張銀照浦琴華
水生生物學(xué)報(bào) 2019年4期
關(guān)鍵詞:血清
高敏敏 吳代武 周露陽 何 杰 葉元土 蔡春芳 吳 萍郁 濃 孫 飛 張銀照 浦琴華
(1. 蘇州大學(xué)基礎(chǔ)醫(yī)學(xué)與生物科學(xué)學(xué)院, 蘇州 215123; 2. 浙江豐宇海洋生物制品有限公司, 舟山 316000;3. 浙江一星實(shí)業(yè)股份有限公司, 海鹽 314300)
膽汁酸(Bile acid, BA)是膽汁的重要組成成分,是膽固醇在代謝過程中所產(chǎn)生的一系列膽烷酸的總稱, 一般以鈉鹽或鉀鹽的形式存在, 稱為膽汁酸鹽。膽汁酸合成的經(jīng)典途徑是以膽固醇為原料, 通過一系列酶促反應(yīng)生成膽酸(Cholic acid, CA)和鵝脫氧膽酸(Chenodeoxycholic acid, CDCA), 膽固醇7α羥化酶(Cholesteral 7α hydroxylase, CYP7A1)是此反應(yīng)的限速酶[1]。合成的膽汁酸在肝毛細(xì)膽管膜處的膽鹽輸出泵(Bile salt export pump, BSEP)介導(dǎo)下形成膽汁流, 分泌到毛細(xì)膽管, 經(jīng)膽汁外排進(jìn)入小腸[2]。膽汁酸在回腸被重吸收入門靜脈, 進(jìn)入血循環(huán)。膽汁酸在肝細(xì)胞血竇面基底膜上的牛磺膽酸鈉協(xié)同轉(zhuǎn)運(yùn)蛋白(Na+-taurocholate contransport peptide, NTCP)介導(dǎo)下被肝細(xì)胞所攝取[3], 再經(jīng)BSEP被分泌入膽小管。膽汁酸從肝臟合成, 貯存于膽囊, 隨膽汁分泌入小腸, 在回腸被重吸收再通過門靜脈回到肝臟的循環(huán)過程稱為膽汁酸的腸肝循環(huán)[4]。膽汁酸腸肝循環(huán)是維持體內(nèi)膽汁酸池大小穩(wěn)定的重要調(diào)節(jié)環(huán)節(jié), 也是調(diào)節(jié)膽汁酸合成速率的重要機(jī)制。
牛磺酸是蛋氨酸、半胱氨酸的代謝終產(chǎn)物, 近年來的研究表明牛磺酸在哺乳動物體內(nèi)具有廣泛的生物學(xué)功能, 包括: 參與膽汁酸的合成和脂肪的代謝, 促進(jìn)脂肪的乳化作用和脂類物質(zhì)(如脂溶性維生素)的消化吸收等。肝臟合成的游離膽汁酸與牛磺酸結(jié)合成牛磺膽酸(Taurocholic acid)及牛磺鵝脫氧膽酸(Taurochenodeoxycholic acid)后, 與BSEP的親和力增強(qiáng), 促進(jìn)了膽汁酸的分泌[5]。隨著魚粉替代蛋白的研究, 牛磺酸在魚類中的研究也受到廣泛地關(guān)注, 但主要是探討其在生長性能、魚體免疫方面的影響[6], 尚無關(guān)于牛磺酸對魚類膽固醇、膽汁酸代謝的相關(guān)報(bào)道。膽固醇、膽汁酸的代謝平衡對魚體健康有重要影響, 高膽固醇會引起脂肪代謝障礙[7]; 而膽汁酸可有效促進(jìn)魚體脂肪的代謝,減少組織脂肪沉積[8]。因此, 研究牛磺酸對魚類膽固醇、膽汁酸代謝的具有重要意義。前期研究發(fā)現(xiàn)以魚粉生產(chǎn)過程中的壓榨液濃縮物——魚溶漿(Stickwater, SW)為蛋白源, 可顯著降低黃顙魚飼料產(chǎn)品中的魚粉需求量, 但同時, 魚溶漿會降低魚體脂肪沉積量[9], 因此本文深入探討魚溶漿對黃顙魚膽汁酸代謝及其組織分布的影響, 為魚溶漿對魚類膽固醇、膽汁酸代謝提供科學(xué)依據(jù)。黃顙魚(Pelteobagrus fulvidraco), 隸屬鲇形目(Siluriformes)、鲿科(Bagridae)、黃顙魚屬(Pelteobagrus), 是一種優(yōu)質(zhì)名貴經(jīng)濟(jì)魚類, 偏肉食性, 具有脂肪含量高的特點(diǎn), 是研究膽汁酸代謝的優(yōu)選對象。
1 材料與方法
1.1 試驗(yàn)材料
試驗(yàn)魚粉為Companfa Pesouera del Pacffico Centro S.A.(Lima, Peru)生產(chǎn)的超級蒸汽魚粉, 魚溶漿由浙江豐宇海洋生物制品有限公司提供, 制備過程見Wu等[9]。魚粉、魚溶漿主要營養(yǎng)成分見表1。魚溶漿中牛磺酸水平是魚粉的5.3倍。
1.2 試驗(yàn)日糧的制備
以30%魚粉日糧為對照(FM), 在無魚粉日糧中分別添加約8% (SW8)、17% (SW17)、25% (SW25)的魚溶漿(干物質(zhì)), 設(shè)計(jì)4組等氮等能日糧(表 2)。在試驗(yàn)配方中, 以混合油脂(魚油∶磷脂油∶豆油=1∶1∶2)平衡各試驗(yàn)日糧脂肪含量, 以磷酸二氫鈣保持各試驗(yàn)配方總磷含量, 大豆?jié)饪s蛋白、棉籽蛋白和雞肉粉同比例變化保持氨基酸平衡, 以米糠粕保持試驗(yàn)日糧配方比例平衡。玉米蛋白粉作為黃顙魚日糧色素來源而各在配方中保持一致。日糧的制作與Wu等[9]一致, 原料粉碎后過60目篩, 按照表 2進(jìn)行飼糧原料的配合。用混合機(jī)混合均勻后,用華祥牌HKj200制粒機(jī)加工成制成直徑1.5 mm,長3—5 mm的顆粒飼糧。所有的試驗(yàn)飼糧風(fēng)干后-20℃密封保存?zhèn)溆谩?/p>
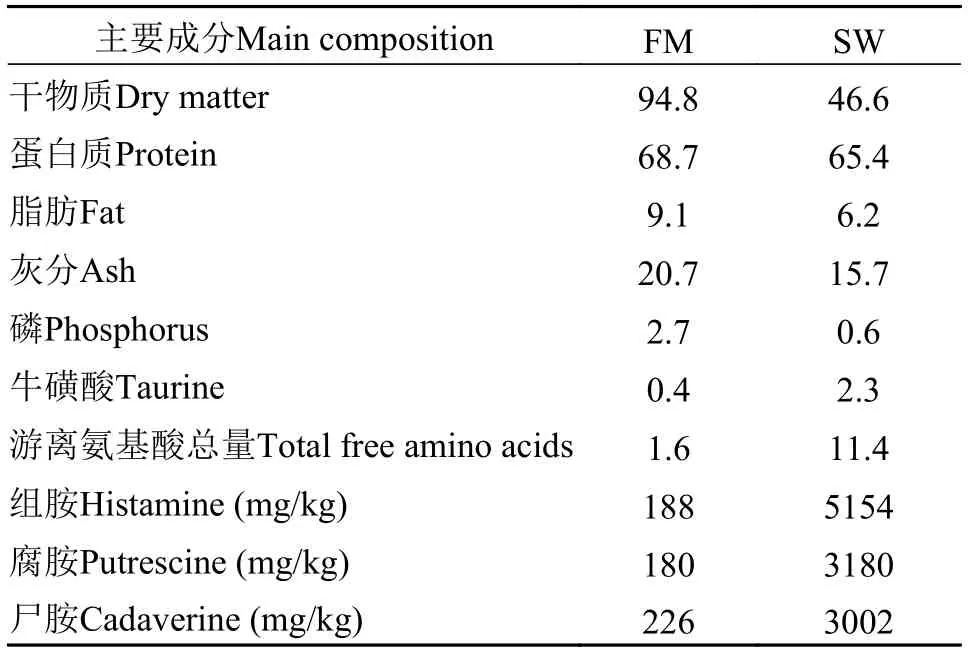
表 1 試驗(yàn)用魚粉、魚溶漿主要成分分析Tab. 1 Chemical composition of experimental fish meal and stickwater (g/100g)
試驗(yàn)日糧營養(yǎng)成分實(shí)測值見表 2, 各組間蛋白質(zhì)、脂肪、總磷、能量水平無顯著差異。SW8、SW17和SW25組組胺、腐胺、尸胺較FM組升高明顯, SW25組是FM組的21.5、14.0和10.5倍。SW17、SW25組試驗(yàn)日糧牛磺酸水平比FM組高55.6%和88.9%。日糧丙二醛、過氧化值無明顯差異, 僅酸價呈現(xiàn)一定程度升高, 表明油脂氧化程度差異性不顯著。
1.3 試驗(yàn)魚與養(yǎng)殖管理
養(yǎng)殖試驗(yàn)在浙江一星養(yǎng)殖基地池塘網(wǎng)箱中進(jìn)行, 在面積為40 m × 60 m的池塘中設(shè)置試驗(yàn)網(wǎng)箱(規(guī)格為1.5 m× 1.5 m × 2.0 m) 12個, 以海鹽縣長山河河水為水源。為保證池塘溶氧均勻, 池塘中間設(shè)置1臺1.5 kW的葉輪式增氧機(jī), 同時設(shè)置1臺2.2 kW的微孔增氧鼓風(fēng)機(jī)。每2個網(wǎng)箱之間, 在水下1.8 m的深度放置一個直徑為0.5 m的圓形微孔增氧盤(增氧盤由直徑20 mm納米曝氣管制成), 投喂期間關(guān)閉增氧設(shè)備, 投喂前使用微孔增氧1h, 其余時間使用葉輪式增氧。
試驗(yàn)用黃顙魚苗購自浙江省湖州農(nóng)業(yè)合作社,運(yùn)輸前停食24h。0.3%食鹽溶液浸泡15min消毒后,200尾/網(wǎng)分配到試驗(yàn)網(wǎng)箱中暫養(yǎng), 馴養(yǎng)2周。選取規(guī)格整齊的黃顙魚種, 0.3%食鹽溶液浸泡15min消毒后, 隨機(jī)分成4組, 每組設(shè)3個重復(fù)(n=3), 共12個試驗(yàn)網(wǎng)箱, 每個網(wǎng)箱46尾黃顙魚。黃顙魚初始體重為(15.67±0.11) g。
試驗(yàn)魚網(wǎng)箱適應(yīng)2周后開始正式投喂。每天投喂3次(5:30—7:00、12:00—13:30和18:00—20:00),日投喂量為體重的3%—5%, 三餐投喂比例2∶2∶3,每10天估算1次魚體增重, 調(diào)整投喂量, 正式投喂60d。每天6:00和18:00記錄池塘水溫, 試驗(yàn)期間水溫25.5—34.4℃。每5天測定水下30 cm的水環(huán)境,養(yǎng)殖期間水體溶解氧濃度>7.0 mg/L, pH 8.0—8.4,氨氮濃度<0.10 mg/L, 亞硝酸鹽濃度<0.005 mg/L,硫化物濃度<0.05 mg/L。
1.4 樣品采集
在試驗(yàn)正式開始時, 隨機(jī)抽取黃顙魚6尾, 作為初始樣本用于全魚體成分分析。正式養(yǎng)殖試驗(yàn)60d后, 停食24h, 進(jìn)行試驗(yàn)結(jié)束采樣工作。包括以下內(nèi)容: 每個網(wǎng)箱黃顙魚進(jìn)行稱重, 計(jì)算增重率。每網(wǎng)箱隨機(jī)取4尾魚保留全魚樣品, 用于體成分分析。隨機(jī)取10尾魚, 以1 mL無菌注射器自尾柄靜脈采血, 自然凝固30min, 3500 r/min、室溫離心10min,取上層血清混合, 液氮速凍后于-50℃冰箱保存, 用于血清生理指標(biāo)的分析; 解剖10尾魚取肝胰臟和腸道內(nèi)容物分別混合, 肝胰臟用于脂肪、膽汁酸測定,腸道內(nèi)容物膽汁酸含量分析; 每網(wǎng)箱隨機(jī)取3尾魚肝胰臟組織小塊置于滅酶EP管中, 液氮速凍, 于-80℃保存?zhèn)溆? 用于基因?qū)W分析。
1.5 樣品分析
主要營養(yǎng)成分原料、日糧、全魚樣品用粉碎機(jī)低溫粉碎均勻, 用LGJ-18B型冷凍干燥機(jī)干燥至恒重測定水分; 凱氏定氮法(GB 5009.5-2010)測定粗蛋白質(zhì)含量; 索氏抽提法(GB/T 14772-2008)測定粗脂肪含量; GB 5009.4-2010中方法測定粗灰分含量; 采用分光光度法(GB/T 6437-2002)測定總磷的含量; 能量采用XRY-1C型氧彈式熱量計(jì)測定。
牛磺酸的測定原料、日糧中氨基酸、牛磺酸含量的測定參考吳代武等[10]的分析方法。
生理指標(biāo)血清谷草轉(zhuǎn)氨酶(AST)、谷丙轉(zhuǎn)氨酶(ALT)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)、膽固醇(CHOL)、膽汁酸(BA)和甘油三酯(TAG)采用雅培C800全自動生化分析儀測定。
肝胰臟、腸道內(nèi)容物與生理鹽水1∶9, 組織勻漿, 5000 r/min離心10min。取上清液用雅培C800全自動生化分析儀測定膽汁酸含量。
熒光實(shí)時定量 PCR(qRT-PCR)取適量肝臟(不超過100 mg), 液氮研磨成粉末狀后, 加入1 mL Trizol (生工)完全覆蓋粉末, 待融化后快速轉(zhuǎn)移至1.5 mL RNAase free EP管中, 按照Trizol法提取樣本的總RNA。之后采用Prime ScriptTMRT reagent kit with gDNA Eraser (TaKaRa)將RNA反轉(zhuǎn)錄成cDNA, 反轉(zhuǎn)錄產(chǎn)物稀釋后于-20℃保存用于熒光定量分析。
基因序列依據(jù)高敏敏等[11]基因測序結(jié)果, 利用Primer Premier 5設(shè)計(jì)定量引物, 引物序列見表 3, 蘇州金唯智公司合成。熒光定量采用SYBR Premix ExTaqTMⅡ試劑盒(Takara, Japan), 反應(yīng)條件為: 預(yù)變性95℃ 30s, 變性95℃ 5s, 退火溫度20s, 40個循環(huán)。溶解曲線程序?yàn)? 95℃ 15s, 60℃ 30s, 20min,95℃ 15s。以DEPC水替代模板作為負(fù)對照, 采用2-ΔΔCt方法來測定目的基因的相對表達(dá)量[12]。
1.6 計(jì)算與分析
原始數(shù)據(jù)經(jīng)Excel 2003初步整理后, 用SPSS 19.0進(jìn)行單因素方差分析(One-Way ANOVA), 事后進(jìn)行Duncan氏多重比較分析試驗(yàn)數(shù)據(jù)的差異顯著性, 結(jié)果以平均值±標(biāo)準(zhǔn)差(Mean±SD)表示, 以P<0.05為差異顯著水平。
2 結(jié)果
2.1 黃顙魚器官組織膽汁酸含量與膽汁酸循環(huán)
黃顙魚肝胰臟、血清、腸道內(nèi)容物中總膽汁酸含量黃顙魚肝胰臟、血清、腸道中膽汁酸水平見圖 1。SW8和SW17組黃顙魚肝胰臟、血清、腸道內(nèi)容物中膽汁酸水平均與FM組無顯著差異, 而SW25組肝胰臟膽汁酸水平降低了76.3% (P<0.05), 血清、腸道內(nèi)容物中膽汁酸水平分別升高了125.7%、123.3%(P<0.05)。
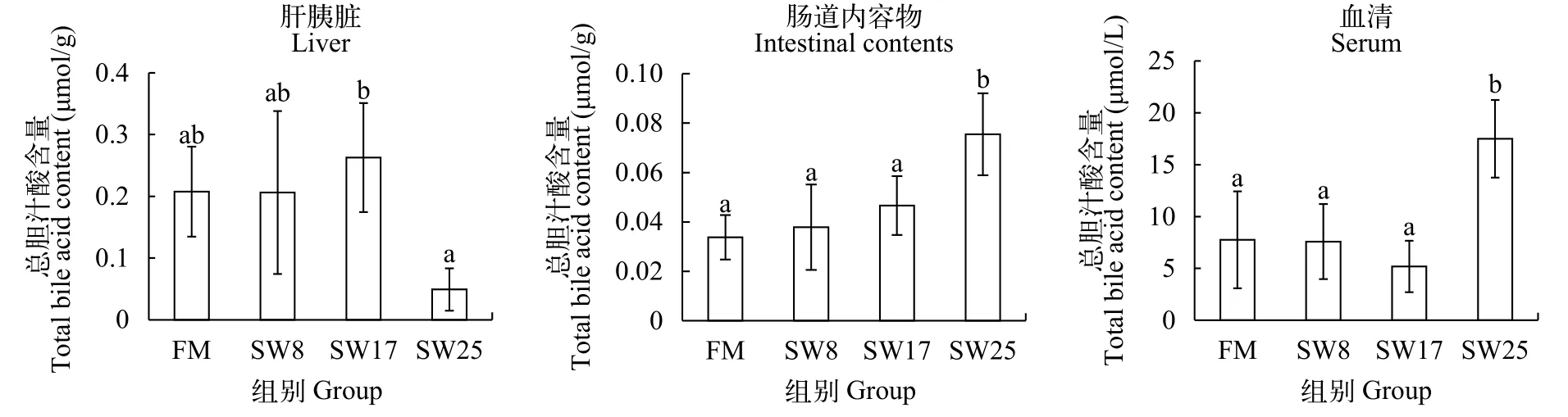
圖 1 魚溶漿對黃顙魚肝胰臟、血清、腸道內(nèi)容物膽汁酸水平的影響Fig. 1 Effects of stickwater on bile acid in liver, serum, intestine of yellow catfish
膽汁酸腸肝循環(huán)代謝關(guān)鍵基因的 mRNA 表達(dá)活性無魚粉日糧中添加8%—25%魚溶漿對黃顙魚肝胰臟CYP7A1 mRNA的表達(dá)量沒有顯著影響(P>0.05, 圖 2), 而BSEP、ABCC4、NTCPmRNA的表達(dá)量隨魚溶漿添加量增加而呈現(xiàn)上升趨勢,SW25組的表達(dá)量比FM組高16.9%、68.2%和222.8%,ABCC4、NTCP基因表達(dá)量變化顯著(P<0.05)。
結(jié)果表明, 無魚粉日糧中添加8%—17%的魚溶漿對黃顙魚肝胰臟膽汁酸的合成沒有顯著性的影響, 對膽汁酸的分泌作用有顯著性的影響。25%魚溶漿不影響膽汁酸的合成, 但會促進(jìn)肝胰臟膽汁酸向血液、腸道中轉(zhuǎn)移。
2.2 黃顙魚魚體脂肪沉積的變化
魚溶漿降低黃顙魚脂肪沉積與FM組相比,黃顙魚全魚蛋白質(zhì)含量不受日糧魚溶漿添加量影響(圖 3), 但全魚脂肪含量降低, SW25組變化顯著(P<0.05), 肝胰臟脂肪含量變化與之相似, SW17和SW25組變化顯著(P<0.05)。
黃顙魚血脂含量與轉(zhuǎn)氨酶活性各試驗(yàn)組血清AST和ALT無顯著差異(表 4), 表明肝胰臟細(xì)胞未受到損傷。SW8組血清CHOL最高, 但與FM組無顯著差異(P>0.05), SW17和SW25組CHOL比FM組低11.9%—16.6%。與FM組相比, 攝食魚溶漿日糧的黃顙魚血清TAG水平降低了32.5%—47.9%,SW17和SW25組變化顯著(P<0.05), 血清GLU、HDL和LDL水平無顯著差異。結(jié)果表明, 在日糧中添加25%魚溶漿促進(jìn)魚體脂肪的代謝過程, 降低脂肪沉積。
3 討論
3.1 魚溶漿對黃顙魚膽汁酸腸肝循環(huán)代謝的影響
前期研究發(fā)現(xiàn), 在黃顙魚日糧中以魚溶漿為蛋白源, 會降低魚體脂肪沉積[9], 在此基礎(chǔ)上, 本文深入探討魚溶漿對黃顙魚膽汁酸代謝及其組織分布的影響。結(jié)果表明, 與魚粉日糧相比, 無魚粉日糧中添加25%的魚溶漿, 肝胰臟膽汁酸水平降低了約76%, 而血清、腸道中膽汁酸水平升高了125%。試驗(yàn)結(jié)果表明魚溶漿的使用是黃顙魚膽汁酸代謝差異變化的主要原因, 而魚溶漿中哪些因素會導(dǎo)致膽汁酸代謝的差異? 依據(jù)魚溶漿、試驗(yàn)日糧中主要化學(xué)成分的分析, 主要包括小肽、牛黃素胺、生物胺、油脂氧化產(chǎn)物等含量在無魚粉日糧中存在較大的差異。
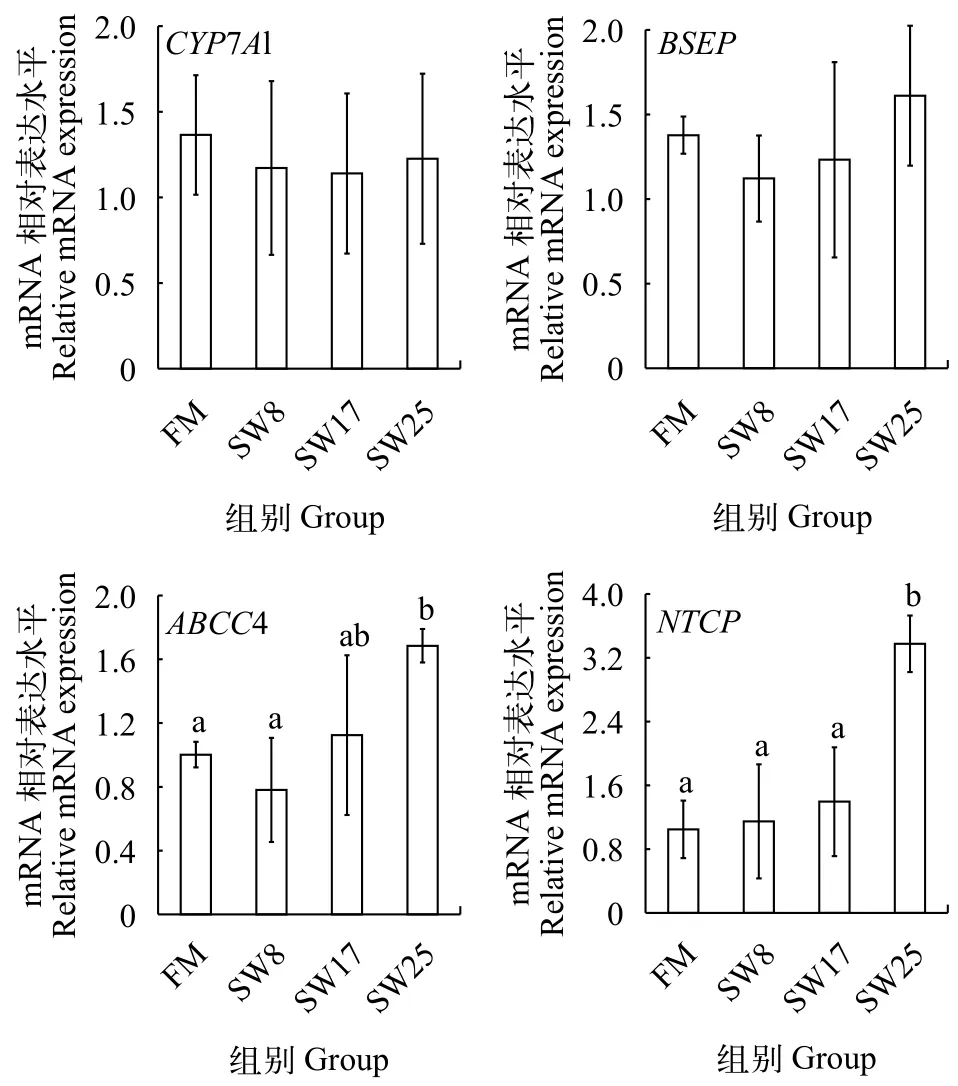
圖 2 魚溶漿對黃顙魚肝胰臟CYP7A1、BSEP、ABCC4、NTCP mRNA的表達(dá)量的影響Fig. 2 Effects of stickwater on the expression of CYP7A1, BSEP,ABCC4 and NTCP in liver
高敏敏等[11]的研究表明, 油脂氧化會導(dǎo)致血清膽汁酸水平降低, 在本試驗(yàn)中油脂氧化程度差異較小, 表明油脂氧化不是本試驗(yàn)中黃顙魚膽汁酸代謝差異性變化的影響因素。各組試驗(yàn)魚血清AST和ALT無顯著差異(表 4), 表明黃顙魚肝胰臟沒有出現(xiàn)損傷, 也不是膽汁酸代謝差異的主要原因。
與魚粉相比, 魚溶漿主要含溶解于水中的蛋白質(zhì)、小肽、生物胺、游離氨基酸等(表 1)。本研究在日糧中添加25%魚溶漿, 黃顙魚肝胰臟BSEP表達(dá)量上調(diào)、腸道膽汁酸水平增加, 是否受組胺等生物胺的影響有待進(jìn)一步研究。目前很少有生物胺對魚類膽汁酸代謝的影響見報(bào)道, 僅何杰等[13]發(fā)現(xiàn)在日糧中添加84.60 mg/kg的組胺不影響黃顙魚血清膽汁酸、膽固醇含量。有研究表明牛磺酸在膽汁酸代謝過程中具有重要的生物學(xué)功能[14—16]。牛磺酸在魚體大量以游離形態(tài)存在, 原料魚在蒸煮過程中牛磺酸更多的進(jìn)入壓榨液, 所以魚溶漿牛磺酸含量遠(yuǎn)高于魚粉(表 1)。研究表明, 肝臟合成的游離膽汁酸與牛磺酸結(jié)合形成牛磺膽酸(Taurocholic acid)及牛磺鵝脫氧膽酸(Taurochenodeoxycholic acid), 增加日糧牛磺酸供應(yīng), 魚體牛磺膽酸、牛磺鵝脫氧膽酸水平會顯著升高[11,17]。肝臟合成的膽汁酸依靠肝毛細(xì)膽管膜處BSEP分泌到毛細(xì)膽管[2],與游離膽汁酸相比, 結(jié)合膽汁酸與BSEP的親和力增強(qiáng)[5,18]。增加日糧魚溶漿添加量, 提高牛磺酸供應(yīng), 可能導(dǎo)致魚體牛磺膽酸、牛磺鵝脫氧膽酸水平會升高, 導(dǎo)致肝胰臟BSEPmRNA的表達(dá)量上調(diào)。BSEPmRNA的上調(diào)促進(jìn)了肝胰臟膽汁酸向膽管分泌, 引起腸道膽汁酸水平的升高, 大量研究也驗(yàn)證了日糧供應(yīng)牛磺酸會促進(jìn)膽汁酸的分泌作用[14—16]。膽汁酸的腸肝循環(huán)是維持體內(nèi)膽汁酸池大小穩(wěn)定的重要調(diào)節(jié)環(huán)節(jié)。陳文等[16]的研究表明, 牛磺酸會促進(jìn)膽汁酸隨糞便的排出, 機(jī)體以膽固醇的降解來補(bǔ)充膽汁酸的排出, 從而起到降低血清CHOL的作用, 意味著魚溶漿的使用會導(dǎo)致腸道膽汁酸的流失,這也會在一定程度上促進(jìn)肝胰臟膽汁酸向腸道的補(bǔ)充分泌。
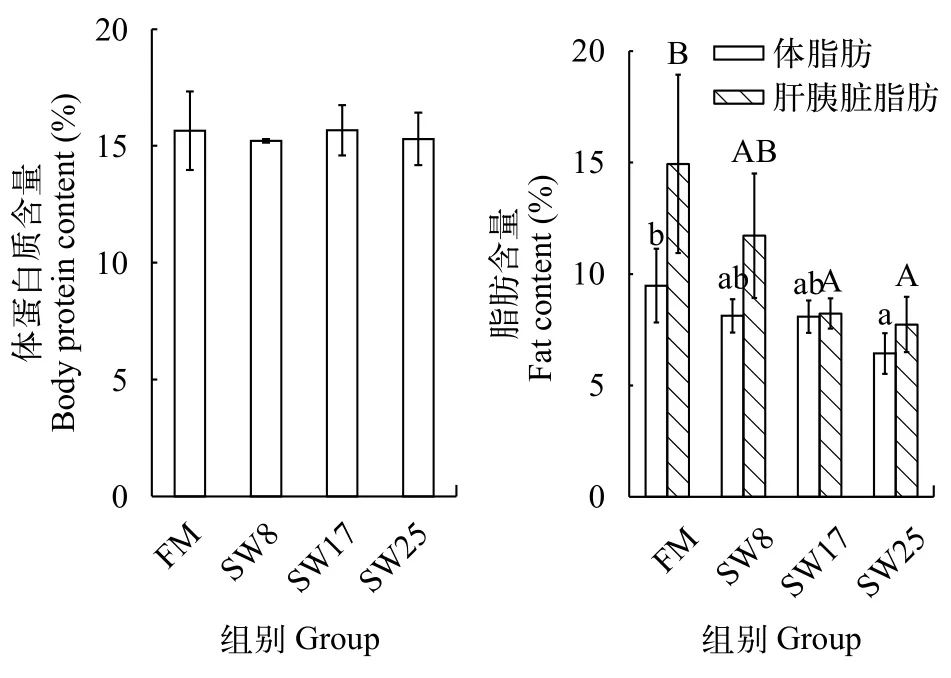
圖 3 魚溶漿對黃顙魚體成分的影響Fig. 3 Effects of stickwater on weight gain and body composition of yellow catfish
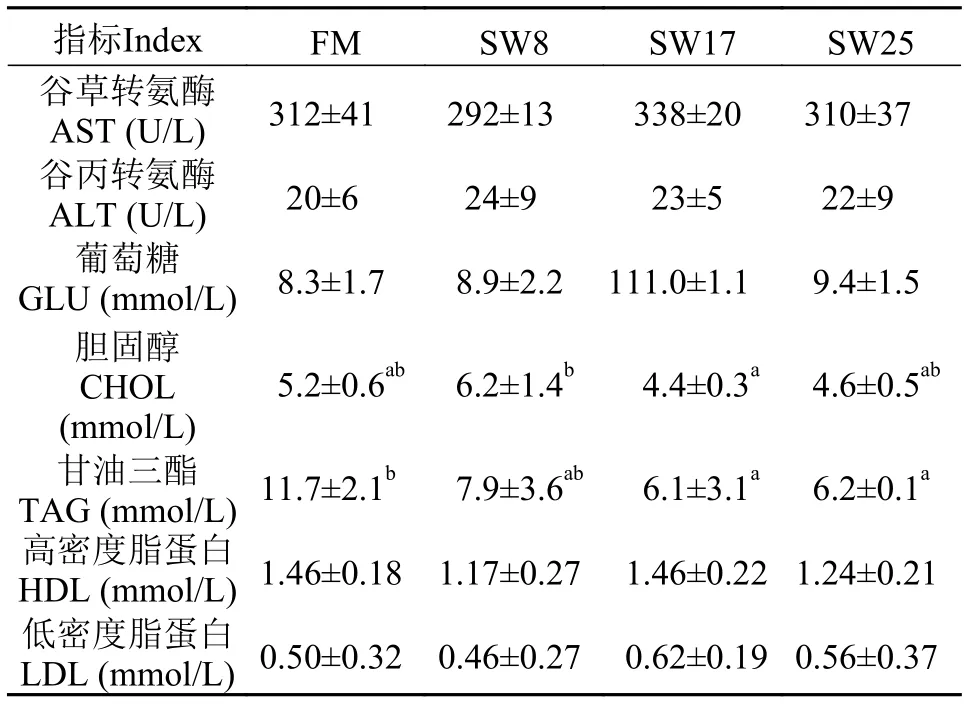
表 4 黃顙魚血清生理指標(biāo)Tab. 4 Serum physiological indexes of yellow catfish
ATP-結(jié)合盒轉(zhuǎn)運(yùn)蛋白C4 (ATP Binding Cassette Transporter C4, ABCC4), 也稱多藥耐藥相關(guān)蛋白4 (Multidrug resistance-associated protein 4,MRP4), 位于肝細(xì)胞基底膜上, 大量動物實(shí)驗(yàn)證明ABCC4對膽汁酸向血液轉(zhuǎn)移具有重要生理作用[19,20]。在哺乳動物的研究發(fā)現(xiàn), 肝臟ABCC4蛋白表達(dá)上調(diào)使膽汁酸改道向血液轉(zhuǎn)移, 最終隨尿液排出體外[21,22]。與BSEP相比, ABCC4與結(jié)合膽汁酸親和力更強(qiáng)[23],故ABCC4表達(dá)量隨魚溶漿添加量增加而上調(diào),25%魚溶漿影響顯著, 導(dǎo)致了肝胰臟膽汁酸向血液轉(zhuǎn)移, 血清膽汁酸水平顯著高于魚粉組。
血循環(huán)中的膽汁酸可在肝細(xì)胞血竇面基底膜上的NTCP介導(dǎo)下被肝細(xì)胞所攝取[3], 日糧添加25%的魚溶漿導(dǎo)致肝胰臟膽汁酸向腸道、血液轉(zhuǎn)移、降低了肝胰臟膽汁酸的含量, 使NTCPmRNA的表達(dá)量上調(diào), 促進(jìn)了膽汁酸的重吸收作用。為了驗(yàn)證肝胰臟膽汁酸水平的降低是否會促進(jìn)膽汁酸的從頭合成, 我們分析了膽汁酸合成限速酶[1]——CYP7A1 mRNA的表達(dá)水平, 結(jié)果發(fā)現(xiàn)魚溶漿并不會影響膽汁酸的合成作用。Liaset等[24]發(fā)現(xiàn)高牛磺酸的酶解魚漿有降低CYP7A1 mRNA表達(dá)量的趨勢, 而膽汁酸的合成量沒有變化。陳文等[16,25]認(rèn)為牛磺酸僅在有外源膽酸鹽的條件下才會誘導(dǎo)CYP7A1的大量表達(dá), 但其誘導(dǎo)機(jī)制尚不清楚。總之, 魚溶漿促進(jìn)肝胰臟膽汁酸向腸道、血液的分泌, 而膽汁酸的合成能力沒有增強(qiáng), 當(dāng)肝胰臟從血液重吸收的膽汁酸不足以彌補(bǔ)自身損失時, 便導(dǎo)致了肝胰臟膽汁酸水平的下降。
3.2 魚溶漿對黃顙魚脂肪沉積代謝的影響
膽固醇、膽汁酸的代謝平衡對魚體健康有重要影響, 高膽固醇會引起脂肪代謝障礙[7,26]; 而膽汁酸可有效乳化脂肪, 促進(jìn)魚體脂肪的代謝, 減少組織脂肪沉積[8,27]。TAG、CHOL是體內(nèi)重要的脂類物質(zhì), 血清CHOL的增高常伴隨TAG的增高[28]。脂肪組織是體內(nèi)最大的CHOL儲存池, 而血清CHOL水平反映了脂肪吸收、代謝狀況[29]。血清CHOL過高會引起脂肪代謝障礙, 陳強(qiáng)等[26]發(fā)現(xiàn)血清CHOL增加會造成肝臟CHOL的過度積累, 病變肝臟(花肝、土肝、綠肝等)的比例上升。在本研究中, 添加25%的魚溶漿, 黃顙魚血清TAG、CHOL均低于魚粉組, TAG降低顯著, 表明魚溶漿可有效促進(jìn)脂肪的代謝, 而全魚和肝胰臟的脂肪含量降低,意味著魚溶漿對降低魚體脂肪沉積有顯著影響。前期研究也發(fā)現(xiàn)草魚日糧中魚溶漿添加量增加, 血清CHOL、TAG呈降低趨勢[30], 與本研究結(jié)果相似。這可能與魚溶漿中牛磺酸含量高有關(guān), 研究表明牛磺酸參與膽汁酸的合成和脂肪的代謝, 促進(jìn)脂肪的乳化作用和脂類物質(zhì)(如脂溶性維生素)的消化吸收等, Li等[31]的研究表明牛磺酸可有效降低血清CHOL和TAG水平。
HDL和LDL是CHOL的主要運(yùn)輸者[32], 分別將CHOL運(yùn)入和運(yùn)出肝臟, 血清LDL與CHOL水平正相關(guān)[28], 草魚日糧中添加6%的魚溶漿使血清HDL升高而LDL降低, 但未出現(xiàn)顯著變化[30], 在本試驗(yàn)中HDL和LDL無顯著變化, 表明25%的魚溶漿對血清HDL和LDL影響不顯著。
4 結(jié)論
無魚粉日糧中添加25% 魚溶漿(干物質(zhì)), 對黃顙魚膽汁酸的合成沒有顯著影響, 但促進(jìn)了膽汁酸從肝胰臟向血液、腸道中轉(zhuǎn)移; 牛磺酸可能是引起膽汁酸代謝變化的主要因素之一; 魚溶漿促進(jìn)了魚體脂肪的能量代謝作用, 降低全魚、肝胰臟脂肪沉積量。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年2期)2021-03-29 05:31:08
昆明醫(yī)科大學(xué)學(xué)報(bào)(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
昆明醫(yī)科大學(xué)學(xué)報(bào)(2020年12期)2021-01-26 00:44:04
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業(yè)科學(xué)(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫(yī)學(xué)院學(xué)報(bào)(2015年5期)2015-12-05 08:22:29
