植物彈狀病毒研究進展
2019-07-22 09:50:46張守杰耿軍紀國才王黎宋春燕李松堅
河北農業科學 2019年2期
關鍵詞:植物
張守杰,耿軍,紀國才,王黎,宋春燕,李松堅*
(1.青島市農業技術推廣站,山東 青島 266071;2.濱城區水產局,山東 濱州 256600;3.青島市農產品質量檢測中心,山東青島 266041;4.嶗山區農業農村局,山東 青島 266100)
彈狀病毒(Rhabdviridae)是一類負義單鏈RNA病毒,因病毒粒子形態似子彈或棒狀而得名[1]。該類病毒寄生宿主很廣,可感染植物、昆蟲、水生生物以及變溫和溫血脊椎動物[2,3]。根據侵染寄主的不同,可以將彈狀病毒分為植物彈狀病毒和動物彈狀病毒兩大類。其中,植物彈狀病毒可以侵染小麥、馬鈴薯、大麥、萵苣和蘭花等多種植物,為害宿主健康,給農業生產造成經濟損失[4,5]。
1 植物彈狀病毒的分類
在國際病毒分類委員會(ICTV)2017年發布的報告中,彈狀病毒共包括131個種,分屬于18個屬。其中,植物彈狀病毒有4個屬,分別是細胞質彈狀病毒屬 (Cytorhabdovirus)、雙分體彈狀病毒屬 (Dichorhavirus)、細胞核彈狀病毒屬(Nucleorhabdovirus)和巨脈病毒屬(Varicosavirus)[6];其他14個屬為動物彈狀病毒。在植物彈狀病毒中,雙分體彈狀病毒屬和巨脈病毒屬為最新添加的2個屬[7,8],其歸類依據是它們分別與細胞核彈狀病毒屬和細胞質彈狀病毒屬的基因組序列有顯著的相似性。雙分體彈狀病毒屬中共有2個種,分別是咖啡環斑病毒(Coffee ringspot virus,CoRSV)和蘭花斑點病毒(Orchid fleck virus,OFV)。巨脈病毒屬中只有萵苣巨脈病毒(Lettuce big veinassociated virus,LBVaV)1個種。此外,根據病毒在宿主細胞中復制場所的不同,可以將植物彈狀病毒分為細胞質彈狀病毒屬和細胞核彈狀病毒屬。其中,細胞質彈狀病毒在細胞質中復制,細胞核彈狀病毒在細胞核中復制。以馬鈴薯黃矮病毒(Potato yellow dwarf virus,PYDV)為代表種的細胞核彈狀病毒共含有10個種。以萵苣壞死黃化病毒(Lettuce necrotic yellows virus,LNYV)、北方禾谷花葉病毒(Northern cereal mosaic virus,NCMV)和大麥黃條點花葉病毒(Barley yellow striate mosaic virus,BYSMV)等為代表種的細胞質彈狀病毒共含有11個種。
2 植物彈狀病毒的基因組結構及蛋白功能
2.1 基因組結構
植物彈狀病毒為負義單鏈RNA病毒[9],其基因組大小為10.8~16.1 kb,占病毒粒子重量的1%~2%。病毒粒子長度100~460 nm,直徑45~100 nm。子彈狀或者棒狀的病毒粒體,由螺旋狀的核衣殼組成。核衣殼由核糖核蛋白(RNP)復合物組成,直徑在30~70 nm,占病毒粒子重量的65%~75%。核糖核蛋白復合物包含基因組RNA、核蛋白(Nucleoprotein,N)、聚合酶相關磷蛋白(Phosphoprotein,P)和RNA依賴性RNA聚合酶(RNA polymerase,L),它們緊密結合在一起,共同發揮作用。核糖核蛋白復合物外側包被有一層脂質包膜和基質蛋白(Matrix protein,M),其中在脂質包膜上結合有病毒包膜糖蛋白(Glycoprotein,G)的三聚體,這些三聚體長度5~10 nm,直徑約3 nm,組成了包被在病毒粒體最外側的一層突起(圖1)。病毒內部還含有5'端加帽酶、鳥苷酸及甲基轉移酶、蛋白激酶(病毒或寄主編碼)、三磷酸核苷酶和二磷酸核苷激酶等多種酶類。
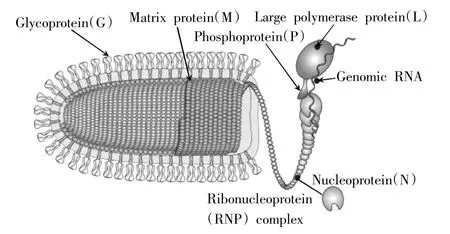
圖1 植物彈狀病毒結構[5]Fig.1 The structure of plant rhabdviridae
現有研究表明,植物彈狀病毒的病毒體通常含有單分子的線性負義單鏈RNA,即使有些彈狀病毒是雙分體,也都是每條RNA獨立包衣的。線性負義單鏈RNA具有3′末端的游離羥基和未聚腺苷酸化的5′三磷酸,末端具有編碼轉錄和復制起始信號的倒序互補序列。全長正向RNA是復制過程中的中間體,約占病毒RNA群體的5%,是構成病毒RNA群體的重要組成部分。與全長負義RNA基因組一樣,反向基因組RNA也會與N緊密結合在一起,而不是以裸露的RNA形式出現。
2.2 蛋白功能
植物彈狀病毒基因組一般含有5個能夠編碼結構蛋白的基因,分別是N、P、M、G和L蛋白編碼基因[10],這些基因通常依照 3′-N-P-M-G-L-5′的順序排列(圖2)[11]。N、P和L蛋白主要參與病毒的轉錄與復制。其中,N蛋白分子量為47~62 kDa,是病毒核衣殼的主要組成成分,可以通過影響轉錄信號的識別過程調節基因組的轉錄和復制,還可以誘導細胞介導的免疫應答反應[11,12]。L聚合酶分子量為220~240kDa,是病毒核衣殼的組分之一,主要負責催化mRNA 5′端加帽等過程[13]。P蛋白是病毒聚合酶的輔助因子,分子量為20~30 kDa,主要功能是介導L蛋白在N-RNA模板上正確定位并連接。在N蛋白合成期間,P蛋白可以作為分子伴侶,通過形成N-P復合物與RNA結合而阻止N蛋白的自我聚集[14,15]。研究表明,植物彈狀病毒的P蛋白在逃避宿主的先天抗病毒反應中具有基礎性作用,可以引起細胞介導的免疫反應。M基質蛋白分子量為20~30 kDa,屬于功能性蛋白,可以調節基因組RNA的轉錄,是病毒粒體的內部組分。M蛋白還可以促進植物彈狀病毒的芽生過程[16~18],參與核質間的運輸和基因的表達。糖蛋白G是病毒的主要抗原蛋白,以三聚體的形式覆蓋在病毒表面,主要負責與宿主細胞受體的結合,可以誘導和結合病毒中和抗體并引發細胞介導的免疫反應[19,20]。目前,普遍認為N、G蛋白與昆蟲介體的傳毒有關。多數植物彈狀病毒還會編碼多個輔助蛋白,其功能在很大程度上是未知的。一般認為,輔助蛋白具有參與病毒在細胞間的移動、調節干擾素應答、逃避免疫等作用。
3 植物彈狀病毒的生物學特性
彈狀病毒是極少數既可以侵染植物,又可以侵染動物的病毒之一。多數植物彈狀病毒以持久增殖的方式進行傳播,通常不會在植物中垂直傳播,但是在昆蟲中可以經卵傳播。植物彈狀病毒主要的傳毒介體有蚜蟲、葉蟬和飛虱等[21],部分病毒還可以通過植物的汁液或宿主的體液進行機械傳播。通過稻飛虱和葉蟬傳播的植物彈狀病毒,不能通過機械傳播的方式侵染植物,但可以通過維管穿刺接種。Louie[22]研究顯示,以蚜蟲為介體的植物彈狀病毒通過摩擦接種傳播的成功率不高。植物彈狀病毒的傳毒效率為5%~100%,差異較大[23~25]。病毒的獲毒時間也存在顯著差異,短的幾分鐘,長的可以達到數天。與在其介體中可以復制的其他病毒一樣,彈狀病毒的潛伏期也存在一定差異,短者為3d,長者達60 d以上[26,27]。有學者發現,玉米花葉病毒(Maize mosaic virus,MMV)能夠侵染其介體玉米花翅飛虱(P.maidis)的包括神經組織在內的大多數組織[28,29]。最近對植物彈狀病毒昆蟲介體的免疫熒光顯微鏡研究結果表明,MMV對P.maidis神經系統的侵染較對其他組織的侵染更廣泛且迅速,MMV可以從介體生物的中腸向前憩室和食管移動,并通過神經系統從這些器官移動到唾液腺,從而避開P.maidis的傳播障礙[30]。這種嗜神經途徑與已經發現的雙翅目宿主中的一些侵染脊椎動物的動物彈狀病毒[31]有些類似。Romoser等[32]研究了其他2種蟲媒病毒在其介體中的傳播途徑,認為內臟肌肉和氣管可以促進蟲媒病毒通過蚊子的腸中層進行移動。盡管在植物彈狀病毒領域已取得較多的研究成果,但目前植物彈狀病毒的傳播途徑和作用機理仍未十分清楚,需要進一步研究。
4 結語
近年來,多種植物彈狀病毒在我國被相繼發現并報道[33,34],且部分植物彈狀病毒的田間為害有蔓延趨勢[35],給農業生產帶來了巨大經濟損失。目前研究較多的植物彈狀病毒(如北方禾谷花葉病毒)以及近幾年在我國新發生的幾種植物彈狀病毒(如大麥黃條點花葉病毒)都有一個共同的寄主——小麥。而中國北方生態密集型小麥栽培范圍較廣,這使得植物彈狀病毒具備了在某些地區暴發并造成嚴重破壞的可能。同時,植物彈狀病毒為一種RNA病毒,其基因組非常容易發生突變,這提高了病毒的適應能力,增加了防治的難度[36],亟需加強防治措施研究。對于植物病毒來講,阻斷其傳播途徑是防止病毒侵染寄主的主要途徑之一。目前,普遍認為彈狀病毒的G蛋白和N蛋白與病毒在介體內的傳播有關。因此,進一步研究植物彈狀病毒蛋白與介體昆蟲蛋白之間的互作關系,明確病毒蛋白在病毒傳播中的作用,對于阻斷病毒傳播,進而防控植物彈狀病毒甚至動物彈狀病毒引起的病毒病具有重要意義。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
