不同規格蝦夷扇貝捕后耐干露特性比較
2019-07-23 00:33:12李亞烜閆麗新田元勇劉俊榮
水產科學 2019年4期
李亞烜,閆麗新,于 笛,田元勇,劉俊榮
( 1.大連海洋大學 食品科學與工程學院,遼寧 大連 116023;2.遼寧省海洋水產科學研究院,遼寧 大連 116023 )
貝類是海洋生物中一個龐大的經濟資源類群,貝類養殖是我國海水養殖第一大產業。北方地區主要養殖品種有蝦夷扇貝(Patinopectenyessoensis)、海灣扇貝(Argopectenirradians)和櫛孔扇貝(Chlamysfarreri)。大部分扇貝加工品以干品和凍品為主,市場價格最高的往往是活品,因此活品保藏極具研究意義。活品蝦夷扇貝作為一種高端海產品,采捕及捕后因各種脅迫因素導致其代謝紊亂從而影響活品品質。近年來,國內有關活品蝦夷扇貝捕后品質的研究陸續有所報道,最初研究從海水養殖蝦夷扇貝供應鏈可追溯管理體系的探討展開[1];隨后,有關活品蝦夷扇貝的感官品質及其評價方法、活品流通過程中的風味品質變化規律等陸續見報道[2-3];此外,對蝦夷扇貝橫紋肌和平滑肌的蛋白分布及理化性質和從腸道菌群角度針對活品供應鏈各環節安全性的研究也有報道[4-5]。
活品扇貝離水早期頻繁暴露在空氣中,環境波動使扇貝正常代謝受到干擾,捕后影響因素包括貯藏方式、蓄養溫度、機械振動等[6],因此扇貝耐干露特性對后期活品貯藏意義較大。通常,市售扇貝除了有不同品種還有不同的規格,而且規格的影響一直被忽視。筆者針對4月大連產蝦夷扇貝,以規格為研究切入點,以離水早期干露為關鍵環節,通過跟蹤pH、三磷酸腺苷及關聯物、糖原、核苷酸能荷以及氨基酸等指標,對蝦夷扇貝規格與早期耐干露特性的關聯開展探索研究,以期為貝類活品保藏提供科學依據。
1 材料與方法
1.1 材料、規格分級與質量組成
1.1.1 試驗材料
試驗用扇貝均為4月采捕于北緯39°底播的蝦夷扇貝,自凈化基地現場挑選健康扇貝,樣品用泡沫箱密封(冰袋降溫)于3 h內運回實驗室。到達實驗室之后立即進行分選和貯藏處理,剔除活力弱及已死亡的蝦夷扇貝。
1.1.2 規格分級
分級方法包括外形尺寸測量及質量稱量。用游標卡尺對殼高、殼長和殼寬進行測量[7],用分析天平稱量單貝質量,將其按殼長分成(105.8±1.5) mm、(98.4±2.3) mm及(90.5±1.7) mm 3個規格組(圖1)。
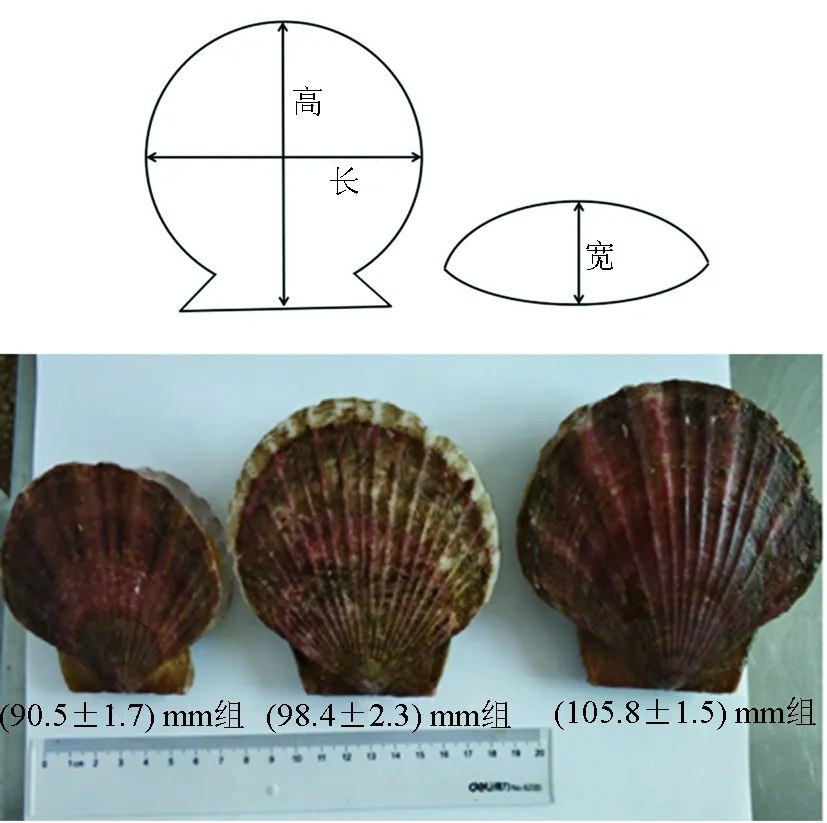
圖1 扇貝外形尺寸及底播蝦夷扇貝商品規格分類
1.1.3 質量組成
每次取6枚活品扇貝,解剖后分別取貝殼、體腔液、閉殼肌及其他軟體組織,取軟體部分置于覆蓋紗布的燒杯上,靜置5 min收集體腔液,用分析天平分別稱量體腔液、橫紋肌以及軟體組織的質量。
1.1.4 其他指標
肥滿度/%=m2/m1×100%
存活率/%=n2/n1×100%
式中,m1為鮮貝整體質量[8],m2為鮮貝軟體質量;n1為扇貝總數,n2為各規格存活扇貝數。
采用單因素方差分析法分析數據間的差異。
1.2 干露處置及采樣
將不同規格組的扇貝分別置于不同的托盤中,用濕布覆蓋并在冰箱中4 ℃貯藏,于0、1、2、3 d后取樣,每12 h更換濕布。每次取6枚蝦夷扇貝,將殼、軟體組織以及外套膜液分離;外套膜液通過雙層紗布過濾收集隨即置于-40 ℃保藏;全部軟體組織立即液氮速凍于-40 ℃保藏。
1.3 化學分析
1.3.1 pH
各組分別取6枚扇貝,剖開后取軟體部分置于覆蓋紗布的燒杯上,靜置5 min用精密pH計測定體腔液pH;取2.0 g扇貝橫紋肌,加入20 mmol/L的碘乙酸鈉10 mL,冰浴下用玻璃棒搗碎,靜置25 min后,用精密pH計測定閉殼肌pH。每組3個重復。
1.3.2 核苷酸系關聯物
取1.0 g扇貝橫紋肌,加入10 mL 5%高氯酸溶液,冰浴條件下搗碎10 min,用2 mol/L KOH調節pH至2.0~3.5,定容至20 mL,3000 r/min離心5 min,取上清液,并用0.45 μm濾膜過濾,取4 mL濾液加入1 mL 0.1 mol/L磷酸緩沖液后待分析。采用高效液相色譜法進行分析。色譜柱:SinoChrom ODS-BP 5 μm,4.6 mm×250 mm(大連伊力特公司);檢測器:二極管陣列檢測器;檢測波長:254 nm;溫度:35 ℃;流動相流速:0.7 mL/min;進樣量:0.02 mL。流動相A:0.05 mol/L K2HPO4-KH2PO4緩沖液(pH 6.5);流動相B:流動相A∶甲醇溶液=8∶2。
1.3.3 腺苷酸能荷
腺苷酸能荷值是用來反映動物受環境脅迫程度的指標,可用于反應扇貝的鮮活程度,按下式計算:
AEC=(2ATP+ADP)/[2(ATP+ADP+AMP)]
式中,AEC為腺苷酸能荷含量,ATP為三磷酸腺苷含量,ADP為二磷酸腺苷含量,AMP為一磷酸腺苷含量。
1.3.4 糖原含量
蒽酮比色法于620 nm測吸光度后計算葡萄糖含量。糖原含量為葡萄糖含量的1.11倍[9]。
1.3.5 氨基酸
將閉殼肌抽提物與10% TCA 1∶1混勻(10 000 r/min離心10 min),用3% TCA稀釋樣品,吸取上清液通過0.45 μm的(水系)針孔過濾膜,濾液采用日立L-8900全自動氨基酸分析儀測定閉殼肌的游離氨基酸。采用酸水解法測定體腔液中氨基酸含量[10],首先取1 g體腔液于水解管中,加入10 mL 6 mol/L HCl、2滴苯酚,-20 ℃放置5 min,取出后吹入氮氣封瓶,110 ℃放置24 h,冷卻后過濾,濾液定容至50 mL,蒸發除去鹽酸,用0.02 mol/L HCl稀釋樣品,通過0.45 μm的(水系)針孔過濾膜,濾液采用日立L-8900全自動氨基酸分析儀進行氨基酸的測定。
1.4 儀器設備
PHS 3C精密pH計(上海精密科學儀器有限公司),BS224S分析天平(北京賽多利斯儀器系統有限公司),BS224S型精密電子天平(北京賽多利斯儀器系統有限公司),Z326K型離心機(HERMLE Labortechnik GmbH),BD314型冰柜(海爾集團青島電冰柜總廠),電磁爐(九陽股份有限公司),SH 2型磁力攪拌器(常州國華電器有限公司)。
2 結 果
2.1 春季產蝦夷扇貝一般規格及肥滿度
大連春季(4月)的蝦夷扇貝基本可以分為大、中、小3個規格,外形尺寸(殼高×殼長×殼寬)依次為(105.3±1.4)mm×(105.8±1.5) mm×(24.2±1.2) mm、(95.7±2.1) mm×(98.4±2.3) mm×(21.7±1.6) mm和(89.8±2.2) mm×(90.5±1.7) mm×(19.6±1.3) mm(表1)。由此可見,區別不同規格的主要指標是殼高與殼長,二者均依次以10 mm左右由大到小遞減;相對而言,殼寬的遞減幅度僅約為2 mm。本研究以殼長指標代替各規格組。各組蝦夷扇貝平均質量由大到小依次為(123.81±7.53) g、(96.68±12.60) g和(77.02±6.83) g。各組單貝平均質量相差20~30 g。
由大到小3規格組蝦夷扇貝肥滿度依次為35.1%、42.6%、35.2%。扇貝殼占總質量百分比最大,(105.8±1.5) mm組為56.6%,(98.4±2.3) mm組為46.9%,(90.5±1.7) mm組為56.5%;各組非可食軟體部位(軟體+體腔液)占總質量達26.9%~35.9%,由大到小3規格組依次為26.9%、35.9%和28.1%;閉殼肌占比小于10%,分別為7.8%、9.6%和7.02%,(98.4±2.3) mm組最高,接近10%(表2)。
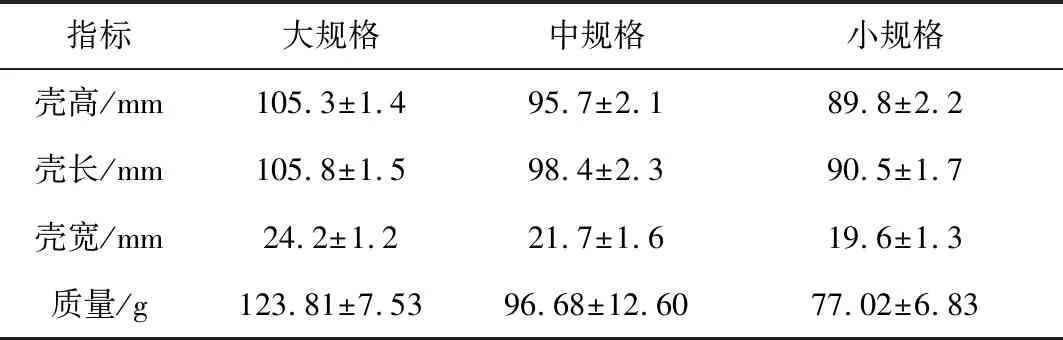
表1 大連春季(4月)底播蝦夷扇貝一般規格分類情況

表2 大連春季(4月)底播蝦夷扇貝各規格組的質量組成及肥滿度
2.2 干露期間閉殼肌的生化變化
2.2.1 閉殼肌pH
3個規格組扇貝肌肉pH變化均呈下降趨勢(圖2)。(105.8±1.5) mm組的肌肉初始pH與(98.4±2.3) mm組幾乎相同,(90.5±1.7) mm組最高;干露3 d后,(98.4±2.3) mm組肌肉pH最低,為6.65,其次是(105.8±1.5) mm組,為6.75,(90.5±1.7) mm組最高,為6.86。干露過程中,較大規格扇貝閉殼肌pH始終低于小規格扇貝。
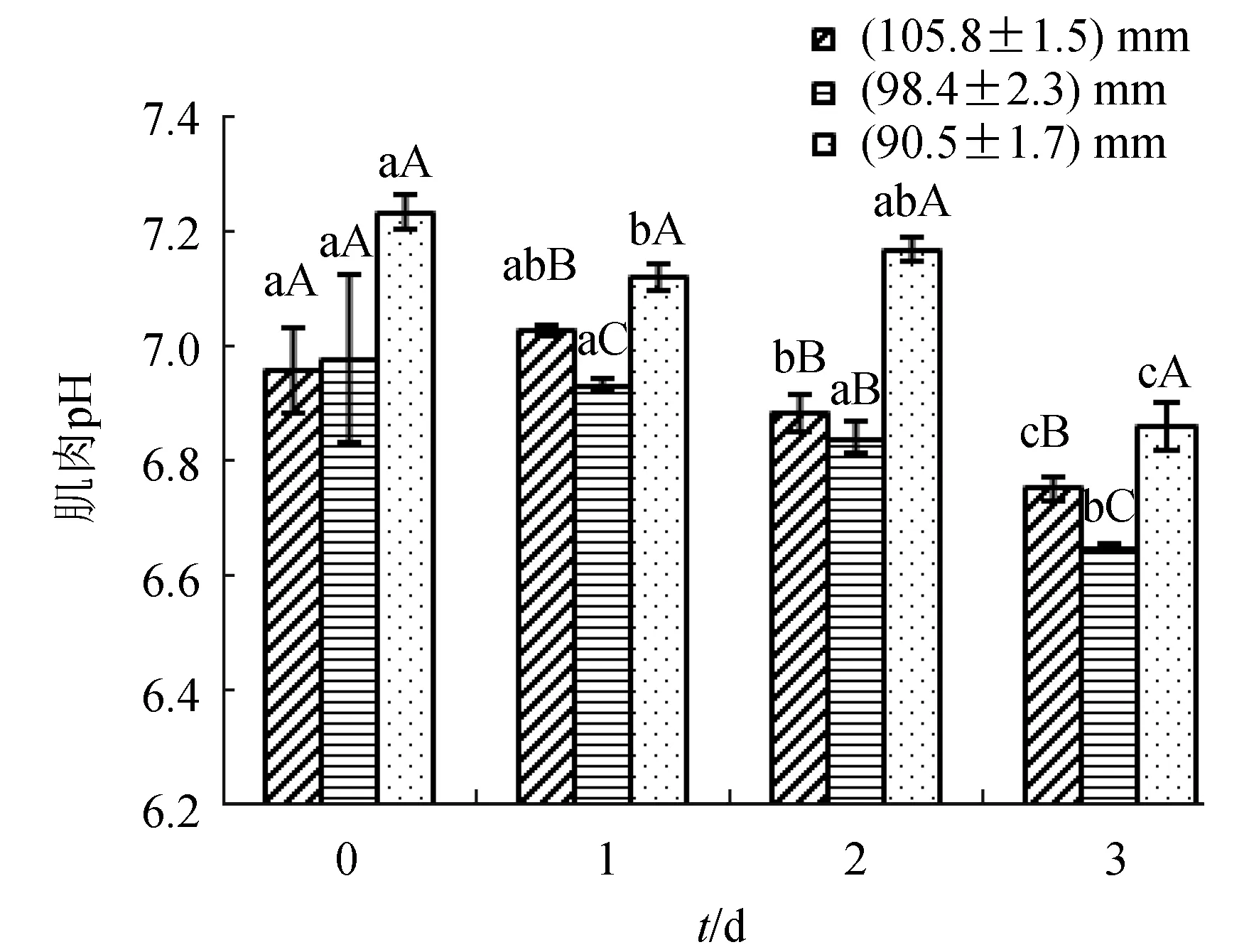
圖2 不同規格的蝦夷扇貝在捕后干露期間肌肉pH的變化不同小寫字母表示同組不同時間差異顯著(P<0.05),不同大寫字母表示不同組相同時間差異顯著(P<0.05).下同.
2.2.2 閉殼肌核苷酸代謝
2.2.2.1 三磷酸腺苷及關聯物
不同規格扇貝的初始三磷酸腺苷含量有差異,(105.8±1.5) mm組為3.227 μmol/g,(98.4±2.3) mm組為3.88 μmol/g,(90.5±1.7) mm組為4.426 μmol/g(圖3);捕后貯藏期間,3種規格扇貝三磷酸腺苷含量均呈現下降趨勢,干露第3 d,較大規格扇貝閉殼肌的三磷酸腺苷含量幾乎耗盡,(105.8±1.5) mm組為0.222 μmol/g,(98.4±2.3) mm組為0.246 μmol/g,而小規格扇貝(90.5±1.7) mm組三磷酸腺苷含量為2.075 μmol/g,仍較高。3種規格扇貝初始二磷酸腺苷含量較低,(105.8±1.5) mm組為0.742 μmol/g,(98.4±2.3) mm組為1.247 μmol/g,(90.5±1.7) mm組為1.080 μmol/g;貯藏期間三磷酸腺苷不斷降解為二磷酸腺苷,干露3 d后,(105.8±1.5) mm組、(98.4±2.3) mm組二磷酸腺苷含量分別為0.84 μmol/g和1.39 μmol/g,(90.5±1.7) mm組為2.342 μmol/g,明顯高于(105.8±1.5) mm組和(98.4±2.3) mm組。3種規格扇貝一磷酸腺苷初始含量均很低,在貯藏期間大部分三磷酸腺苷和二磷酸腺苷轉化為不提供能量的一磷酸腺苷,其中(105.8±1.5) mm組一磷酸腺苷含量為2.33 μmol/g,(98.4±2.3) mm組為3.23 μmol/g,(90.5±1.7) mm組為1.33 μmol/g。

圖3 干露時間對不同規格蝦夷扇貝閉殼肌三磷酸腺苷及關聯物的影響
2.2.2.2 核苷酸能荷變化及存活率
干露3 d,3種規格扇貝核苷酸能荷均呈下降趨勢,其中(105.8±1.5) mm組、(98.4±2.3) mm組下降尤其明顯(圖4)。由小到大3種規格組的初始核苷酸能荷分別為0.877、0.835和0.856;干露2 d,(105.8±1.5) mm組和(98.4±2.3) mm組核苷酸能荷有明顯的下降,分別為0.55和0.65;干露3 d,(105.8±1.5) mm組、(98.4±2.3) mm組核苷酸能荷已經低于0.2,而此時(90.5±1.7) mm組接近0.6。
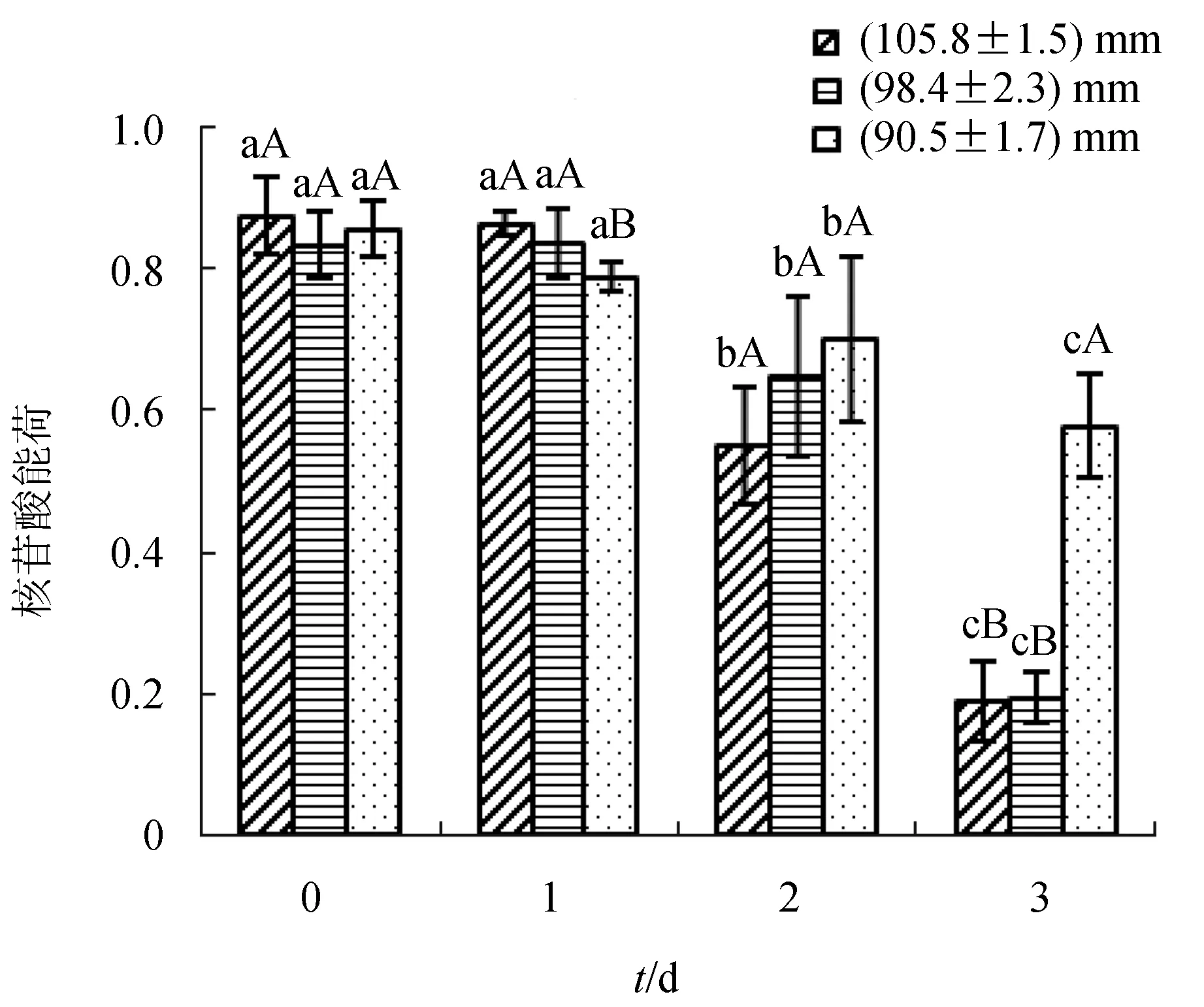
圖4 干露時間對不同規格蝦夷扇貝核苷酸能荷的影響
干露2 d,3種規格扇貝存活率始終保持在100%;干露3 d,(105.8±1.5) mm組和(98.4±2.3) mm組扇貝存活率低于50%,(89.8±2.2) mm組規格扇貝存活率為67%(表3)。

表3 蝦夷扇貝捕后干露期間每日存活率 %
2.2.3 閉殼肌糖原含量變化
干露貯藏期間,(105.8±1.5) mm組、(98.4±2.3) mm組糖原在干露2 d呈現下降趨勢,(90.5±1.7) mm組下降不明顯,干露3 d時糖原含量幾乎與前1 d相同;干露貯藏前2 d,3種規格扇貝差異不明顯,干露3 d,(105.8±1.5) mm組、(98.4±2.3) mm組規格扇貝糖原低于(90.5±1.7) mm組(圖5)。

圖5 不同規格蝦夷扇貝捕后干露期間糖原含量變化
2.2.4 閉殼肌游離氨基酸含量變化
大連春季(4月)的蝦夷扇貝閉殼肌游離氨基酸中,甘氨酸和精氨酸含量明顯高于其他氨基酸,其次是谷氨酸和丙氨酸;各組甘氨酸含量分別為:(105.8±1.5) mm組81.99 nmol/g,(98.4±2.3) mm組139.82 nmol/g,(90.5±1.7) mm組174.39 nmol/g,干露3 d后各規格組甘氨酸含量分別為:(105.8±1.5) mm組100.55 nmol/g,(98.4±2.3) mm組149.16 nmol/g,(90.5±1.7) mm組182.25 nmol/g(表4)。
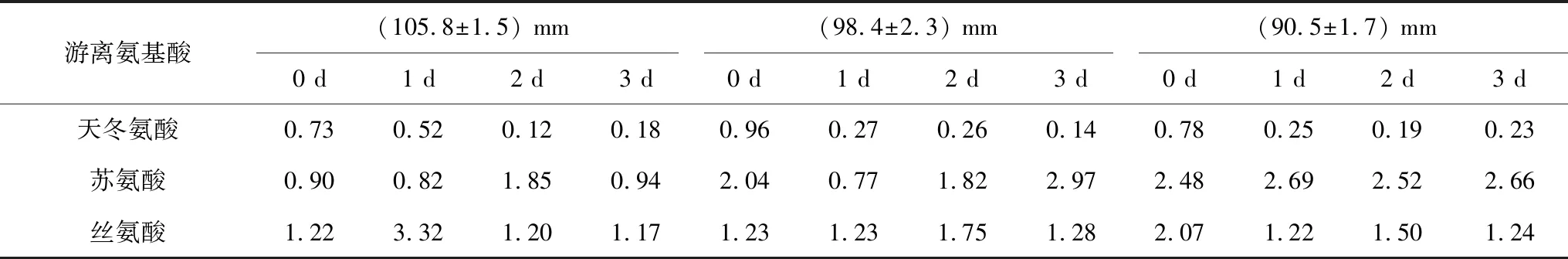
表4 大連春季(4月)蝦夷扇貝閉殼肌游離氨基酸分布及干露變化 nmol/g
續表4

游離氨基酸(105.8±1.5) mm(98.4±2.3) mm(90.5±1.7) mm0 d1 d2 d3 d0 d1 d2 d3 d0 d1 d2 d3 d谷氨酸4.997.497.464.367.117.246.997.675.7910.156.399.48甘氨酸81.99128.08192.30100.55139.80159.90198.20149.16174.39173.94199.50182.25丙氨酸4.469.0811.768.386.433.9510.5019.317.7518.959.5927.77纈氨酸0.640.971.050.541.010.520.811.552.272.011.992.09甲硫氨酸0.540.710.970.490.730.110.691.652.422.221.761.92異亮氨酸0.180.440.3000.410.180.261.271.521.361.491.42亮氨酸0.330.470.410.100.420.210.311.892.272.292.092.38賴氨酸0.790.320.460.220.450.350.300.610.962.210.471.10組氨酸0.230.160.660.210.260.160.300.450.641.610.690.74精氨酸28.9927.5235.7023.5934.3638.6738.3331.2431.2136.0135.4228.85氨1.792.462.504.131.702.411.943.241.712.552.473.28
2.3 干露期間體腔液的生化變化
2.3.1 體腔液pH
3種規格扇貝在貯藏前2 d體腔液pH有下降趨勢但是不明顯,干露3 d,(105.8±1.5) mm組和(90.5±1.7) mm組規格扇貝的體腔液pH有所上升(圖6)。
2.3.2 體腔液含氮物組成
體腔液中含氮物組成見表5,其中氨含量較高,其他氨基酸含量均較低且無明顯變化。

圖6 不同規格的蝦夷扇貝在捕后干露期間體腔液pH的變化

表5 體腔液含氮物組成 mg/g
3 討 論
3.1 扇貝規格分級方法及其質量組成的特點
扇貝的商業規格分級有多維參數作為考量。本研究主要從外形尺寸及單貝質量兩方面對規格進行常規分級。另外,貝齡也是判斷商品價值的一個重要參考指標,且與規格有密切關聯。貝齡的判斷可通過使用雙目顯微鏡計數在韌帶上的生長區并且遵循Mason等[11]描述的程序來確定年齡。閉殼肌是扇貝品質研究的焦點,國內外對其活品品質尤為關注[12-14],作為重要的可食部位,閉殼肌質量占比不到10%,其中(98.4±2.3) mm組扇貝閉殼肌占總質量的9.6%,(105.8±1.5) mm組和(90.5±1.7) mm組占比分別為7.8%和7.02%,由此可知,(98.4±2.3) mm組扇貝閉殼肌出成率優于其他兩組。扇貝不可食部位占比較高,這些副產物的開發利用對于扇貝產業也非常重要,例如近年來蝦夷扇貝殼開發利用主要集中在工藝品制造、醫藥衛生、輕工業等領域[15];軟體組織(扇貝內臟團、裙邊以及性腺)雖食用價值不高,但扇貝性腺可制備多肽[16],扇貝裙邊營養成分豐富[16],內臟團含有大量生物活性物質,具有重要的藥用價值[17-18]。本研究中,(98.4±2.3) mm組扇貝殼占比最低為46.9%,(105.8±1.5) mm組與(90.5±1.7) mm組分別為56.6%和56.5%;(98.4±2.3) mm組扇貝其他軟體部分部位占比最高為35.9%,(90.5±1.7) mm組與(105.8±1.5) mm組分別為28.1%和26.9%。(105.8±1.5) mm組和(90.5±1.7) mm組扇貝肥滿度相近,分別為35.1%和35.2%;(98.4±2.3) mm組具有貝殼占比小且軟體部位占比大的特點,因此大連春季蝦夷扇貝(98.4±2.3) mm組規格最飽滿。對占總質量比例不到8%的體腔液關注度較少,其氨基酸等營養成分值得進一步研究。
3.2 捕后初期脅迫應激下的活品蝦夷扇貝生化代謝特性
3.2.1 規格大小與活品扇貝中的三磷酸腺苷及其降解
扇貝離水后初期頻繁暴露在空氣中,相對于捕前生長狀態面臨急劇的脅迫。閉殼肌是扇貝主要的可食部位,同時也是肌肉能量代謝的重要場所,肌肉中三磷酸腺苷含量能夠較好地反映扇貝的生理狀態[19]。三磷酸腺苷在水解酶作用下釋放出其中能量,轉化為二磷酸腺苷;相反二磷酸腺苷接受能量和一個磷酸基團可合成為一個三磷酸腺苷;二磷酸腺苷與一磷酸腺苷相互轉化原理相同。干露初始,小規格扇貝三磷酸腺苷含量高于較大規格扇貝,閉殼肌的三磷酸腺苷含量與規格大小成反比,可能是由于采捕及運輸期間面臨機械碰撞及密度、濕度、溫度及氧氣等脅迫因子的急劇變化,較大規格扇貝面臨的應激反應更加強烈所致。隨著干露時間的增加,蝦夷扇貝受到低溫缺氧等脅迫加強,三磷酸腺苷被逐步消耗,各組中三磷酸腺苷含量均明顯下降,干露3 d時,(105.8±1.5) mm組、(98.4±2.3) mm組扇貝的閉殼肌三磷酸腺苷含量幾乎耗盡,而(90.5±1.7) mm組含量尚保持為2.075 μmol/g,其含量明顯高于(105.8±1.5) mm組、(98.4±2.3) mm組,可見小規格扇貝比大規格扇貝具有更好的耐干露特性。隨著干露時間的延長,三磷酸腺苷逐漸被消耗分解,相應地,3種規格扇貝二磷酸腺苷和一磷酸腺苷含量逐漸積累,干露3 d后,大部分三磷酸腺苷轉化為二磷酸腺苷,(90.5±1.7) mm組扇貝二磷酸腺苷明顯高于其他兩組,進一步說明了在貯藏過程中小規格扇貝狀態明顯好于較大規格扇貝。隨貯藏時間的延長,二磷酸腺苷與三磷酸腺苷逐漸降解為一磷酸腺苷,干露貯藏至3 d時扇貝開始出現嚴重縮邊甚至死亡,(105.8±1.5) mm組、(98.4±2.3) mm組扇貝狀態不好,而(90.5±1.7) mm組扇貝在貯藏3 d時狀態仍然較好,其肌肉中一磷酸腺苷含量低于其他兩組,由此也可看出,小規格扇貝比大規格扇貝具有更強的耐干露特性。
核苷酸能荷這一指標被廣泛應用于評價貝類的生理狀態,能夠靈敏地反映貝類所受到的脅迫[20]。核苷酸能荷值分析顯示,干露初始時,3組扇貝狀態良好均可繁殖;干露3 d后,(105.8±1.5) mm組、(98.4±2.3) mm組扇貝已處于無法繁殖的不可逆狀態,(90.5±1.7) mm組扇貝處于緩慢生長但可恢復繁殖的狀態。相同規格不同干露時間之間比較,干露2 d和3 d時核苷酸能荷與之前均呈現顯著差異;相同甘露時間不同規格比較,干露2 d時,(90.5±1.7) mm組扇貝核苷酸能荷略高于其他兩組,干露3 d時,(90.5±1.7) mm組扇貝核苷酸能荷顯著高于其他兩組。干露期間每日存活率可體現出不同規格扇貝的耐干露特性,根據所得數據可看出,在貯藏過程中,(90.5±1.7) mm組耐干露特性優于其他兩組。
3.2.2 規格大小與活品扇貝中的糖原及其降解
貝類肌肉中,糖原直接氧化分解產生能量供給肌肉收縮,因此,糖原的變化情況是反映貝類生理狀態的重要指標,其儲備情況對扇貝捕后的品質很重要[21]。在干露貯藏期間,較大規格扇貝糖原含量有下降趨勢,小規格扇貝下降不明顯。干露3 d時,(105.8±1.5) mm組、(98.4±2.3) mm組扇貝糖原含量低于(90.5±1.7) mm組,可能因為(105.8±1.5) mm組和(98.4±2.3) mm組扇貝耐干露特性不如(90.5±1.7) mm組,貯藏期間前兩組的三磷酸腺苷降解為不提供能量的一磷酸腺苷,而糖原則成為供能物質,消耗較多所致。通常情況下,糖酵解會導致酸性代謝產物積累,肌肉組織pH略有下降[22]。在本研究中,干露期間低環境供氧下,扇貝肌肉進行無氧呼吸,隨著章魚堿等酸性糖酵解產物積累,導致扇貝的肌肉pH降低。試驗結果也顯示,干露3 d,各規格組扇貝肌肉pH均呈現降低趨勢;不同規格之間,初始時3種規格扇貝pH差異不顯著,其他時間,(90.5±1.7) mm組均與其他兩組存在顯著差異。
3.2.3 規格與其他干露貯藏變化
貝類肌肉中的游離氨基酸與核苷酸關聯物均為含氮呈味物質,這些呈味物質的組成和含量的不同,會使原料呈現出各具特色的特征滋味,其中谷氨酸是海產貝類中共有的鮮味貢獻者,甘氨酸是貝類中共有的甜味貢獻者[23]。含量最高的甘氨酸在干露2 d時含量呈現上升趨勢,干露3 d時含量下降;其余氨基酸含量變化趨勢不明顯。不同規格之間比較,小規格扇貝甘氨酸初始含量優于較大規格扇貝;干露3 d后,小規格扇貝甘氨酸含量仍然優于較大規格扇貝。
貝類是開放性系統,不存在特異性體液免疫,體腔液構成貝類以血細胞為基礎的細胞和體液系統,在貝類免疫學領域具有重要研究價值[24]。本研究對干露處理下的厭氧代謝所引起的體腔液pH變化也做了初步分析,整體來看,3 d干露期間,體腔液pH無明顯變化,(105.8±1.5) mm組pH在干露3 d的上升,可以解釋為氨氮積累所致,與干露3 d出現的死亡現象相對應。
貝類體腔液在蒸煮加工過程中,通常作為副產物被排放[25-26]。其中不乏兼具營養美味的含氮物,基于這個考慮,重點對體腔液的含氮物進行了分析。分析結果顯示,體腔液中氨含量較高,可能由于貝類處在干露條件下,代謝產生的氨氮積累無法消除所致。其他處置條件下體腔液的含氮物分布及變化,及有機物和礦物質分布情況有待進一步研究。
4 結 論
經過對不同規格蝦夷扇貝捕后耐干露特性的比較,主要得出結論:
(1)大連春季蝦夷扇貝體質量為70~130 g/個,扇貝不可食部位(殼+軟體部位)占總質量比例為90%,主要可食部位(閉殼肌)占比不到10%;不同規格間外形尺寸的差異主要表現在殼高與殼長,殼厚不明顯;
(2)中等規格(98.4±2.3) mm組(約90 g/個)相比其他兩組,在肥滿度及閉殼肌出成率方面具有突出優勢;
(3)小規格(90.5±1.7) mm組扇貝(約70 g/個)在耐捕后脅迫方面有明顯優勢,初始三磷酸腺苷含量、干露3 d后三磷酸腺苷含量和核苷酸能荷值與較大規格扇貝[(105.8±1.5) mm組和(98.4±2.3) mm組]相比有明顯優勢。
