復合微生物菌劑對棉花生理特性及根際土壤微生物和化學性質的影響①
2019-07-26 02:57:58宋以玲陳士更肖承澤李玉環蘇秀榮丁方軍
土壤 2019年3期
關鍵詞:生長
宋以玲,于 建,陳士更,肖承澤,李玉環,蘇秀榮,丁方軍,
復合微生物菌劑對棉花生理特性及根際土壤微生物和化學性質的影響①
宋以玲1,2,于 建1,2,陳士更1,2,肖承澤1,2,李玉環3,蘇秀榮4,丁方軍1,2,3*
(1 山東農大肥業科技有限公司,山東泰安 271000;2 山東省腐植酸高效利用工程技術研究中心,山東泰安 271000;3 山東農業大學資源與環境學院,山東泰安 271018;4 山東農業大學化學學院,山東泰安 271018)
為了系統地闡述復合微生物菌劑在改善土壤化學性質、生物學性質和促進植物生長方面的效果及機理,采用盆栽試驗,研究常規和滅菌條件下不同用量的微生物菌劑對棉花生長、生理代謝、根際土壤生物學特性和養分含量的影響。結果表明:施用復合微生物菌劑能夠提高苗期棉花葉片光合色素總量,葉片和根系內抗氧化物酶(SOD、POD、CAT)活性,降低過氧化物質含量(MDA、O2? ?),進而提高了棉花的光合性能和抗氧化能力;降低了土壤pH,提高了根際土壤細菌和放線菌數,而降低了根際土壤真菌數,同時提高了根際土壤脲酶、中性磷酸酶、蔗糖酶和脫氫酶活性,最終提高了根際土壤有效養分和有機質含量,改善了根際微域環境,促進了棉花苗期生物量的累積和抗病性。除根際土壤養分含量外,常規復合微生物菌劑對各指標的改善效果優于滅菌復合微生物菌劑,且用量為10 L/667m2時效果較明顯。因此,復合微生物菌劑可通過改變土壤pH,改善根際土壤微生物群落的數量和結構來改變土壤酶活性和有效養分含量,優化根際生長環境,增強植物抗逆性和光合能力,最終實現養地增產的效果。此外,復合微生物菌劑中的有益活菌在改土、促生方面具有極其重要的積極效應。
復合微生物菌劑;棉花;抗氧化系統;土壤微生物;土壤酶
隨著我國經濟的快速發展,農業生產中化肥、農藥投入量日益增多,對土壤、水體、大氣與生態平衡造成了嚴重的破壞,土壤退化、污染嚴重,對作物產量、品質、食品安全性和人體健康也造成了嚴重的威脅[1]。隨著人們環境安全意識的提升,開始注重食品的安全性和品質,因此我國開始大力推進新型肥料的使用,實施減少化肥、農藥投入量的“雙減政策”。復合微生物菌劑本身含有大量高效有益活菌,施入土壤后在植物根際定殖,同時激活土著微生物,能在一定程度上降解污染環境的有機物[2];復合微生物菌劑還含有大量活性物質和有機質等營養成分,具有提高土壤微生物豐富度、優化土壤團粒結構、調節根部營養環境等作用;此外,在其生命活動過程中還能產生各類植物生長刺激素,刺激植物生長[3-4]。因此,在自然生態系統中,微生物與植物的有益共生在防治植物病害和促進植物生長等方面發揮了重要的作用[5]。丁莉萍等[6]研究發現復合菌劑在提高樟樹苗移栽成活率的同時,提高了土壤中有效養分含量,促進樟樹苗對養分的吸收和利用,為樟樹苗移栽后的生長打下了堅實的基礎。前人大量研究表明:復合微生物菌劑可提高果園土壤微生物多樣性,促進重茬果園土壤微生物生態系統的恢復[7];通過提高對花生根際化感物質的降解,來減輕花生網斑病、焦斑病和褐斑病的發病率,緩解連作障礙的同時,減少藥物使用帶來的環境污染[8];還可通過提高鹽堿土耕層土壤脫鹽率,降低土壤鹽分含量,減輕鹽害脅迫,促進苜蓿生長[9]。因此,應用微生物菌劑來調控土壤微生物群落結構和多樣性、改善土壤營養條件、增強植物抗逆能力、減輕病蟲害的發生,實現農業健康持續發展具有深遠的意義。
棉花是我國僅次于糧食的第二大作物,也是關系國計民生的戰略物資。然而,棉花主產區多為貧瘠土壤區,為了提高產量,人們加大了化肥的投入量,隨著生產周期的推移,導致土壤板結、土壤團粒結構和通透性變差、耕地肥力下降、連作障礙等諸多問題[10]。研究表明:合理配施有機無機肥利于棉花的增產和增質,同時對土壤環境的改善也具有重要作用[11]。微生物肥料在與有機肥混施條件下提高了棉花產量、土壤肥力,降低了土壤容重,且微生物肥料與有機肥混施效果優于單一使用[12]。然而有關復合微生物菌劑對根際土壤微生物群落與土壤酶活性的影響及促進植物生長的機理等研究較少。因此,本試驗以棉花為供試植物,通過盆栽試驗,分析常規和滅菌條件下不同用量的復合微生物菌劑對棉花生長、生理代謝及土壤養分、酶活性和土壤微生物數量的影響,研究復合微生物菌劑的改土、促生效果及機理,對比常規與滅菌條件下的差異,同時探討適宜的推薦用量,為復合微生物菌劑的大面積推廣應用提供科學依據。
1 材料與方法
1.1 供試材料
試驗于2017年7月15日—8月30日在山東省泰安市肥城市農大肥業科技有限公司蔬菜大棚內實施,供試土壤基本理化性狀:堿解氮60.38 mg/kg,全氮 0.93 g/kg,有效磷 31.74 mg/kg,速效鉀 95.09 mg/kg,有機質9.83 g/kg;供試作物:魯棉研28號;供試肥料:農大肥業科技有限公司微生物肥料車間提供的常規復合微生物菌劑(有效活菌數≥10億/ml;有效氮磷鉀含量≥6%)。滅菌復合微生物菌劑:常規復合微生物菌劑在120 ℃條件下滅菌30 min后獲得。
1.2 試驗設計
常規和滅菌復合微生物菌劑各設2個施用量,不施肥處理為空白對照,復合微生物菌劑做追肥施入,共設5個處理,分別為: CK,不施肥;T1,滅菌復合微生物菌劑 5 L/667m2;T2,滅菌復合微生物菌劑 10 L/667m2;T3,常規復合微生物菌劑 5 L/667m2;T4,常規復合微生物菌劑 10 L/667m2,每個處理設6次重復,采用隨機區組試驗;種播棉花,兩片真葉后間苗定植,每盆留3棵棉花幼苗,總共30盆。定植后統一管理,每隔5 d處理1次,每個處理所需肥料總量溶于1.8 L水中稀釋,即每盆施入300 ml稀釋液,空白用等量蒸餾水,共處理8次,直至收獲。
1.3 測定項目與方法
1.3.1 生長指標和產量的測定 棉花幼苗生長期用直尺、游標卡尺和手持SPAD-502 PLUS葉綠素儀測定棉花株高、莖粗和葉片SPAD值;處理45 d后收獲,將植株地上部和地下部分開,根系先用5 mmol/L CaCl2沖洗,再用蒸餾水沖洗干凈,稱鮮物質量;然后在110 ℃下殺青,80 ℃烘干至恒重,測干物質量。
1.3.2 植株生理指標測定 光合色素含量的測定:用 95% 的乙醇研磨,25 ml定容后,分別在665、649和470 nm下測定吸光值[13];超氧化物歧化酶(SOD)活性測定:采用氮藍四唑(NBT)法[14];過氧化物酶(POD)活性測定:采用愈創木酚法;過氧化氫酶(CAT)活性測定:采用紫外吸收法[15];丙二醛(MDA)含量測定:采用硫代巴比妥酸法[16];超氧陰離子(O2? ?)產生速率測定:采用羥氨氧化法[17]。
1.3.3 土壤各項指標測定 用5︰1的水土比浸提法測土壤pH;采用平板菌落計數法測根際土壤細菌數、真菌數和放線菌數[18];采用重鉻酸鉀容量法測土壤有機質;采用凱氏定氮法測全氮;采用堿擴散法測堿解氮;采用碳酸氫鈉溶解-鉬銻抗比色法測有效磷;采用乙酸銨溶解-火焰光度計法測有效鉀[17];采用苯酚-次氯酸鈉比色法測脲酶活性,以24 h后1 g土壤中NH4+-N的質量(mg)表示;采用滴定法測過氧化氫酶活性,用20 min后1 g土壤消耗0.02 mol/LKMnO4的ml數表示;采用3,5-二硝基水楊酸比色法測蔗糖酶活性,以24 h后1 g土壤中所含葡萄糖的質量(mg)表示;采用磷酸苯二鈉比色法測土壤磷酸酶活性,以24 h后1 g土壤中釋放出的酚并轉化為磷的質量(mg)表示;采用氯化三苯基四氮唑法測土壤脫氫酶活性,結果以1 g土壤中三苯基甲臢(TPF)的質量(μg)表示[19]。
1.4 數據處理
采用Excel 2003軟件處理數據和繪表,采用DPS 7.05軟件進行統計分析,采用最小顯著極差法(LSD)進行差異顯著性檢驗(< 0.05)。
2 結果與分析
2.1 復合微生物菌劑對棉花生長的影響
由表1可知,不同處理對生長25、35和45 d后棉花株高、莖粗和SPAD值的影響不同,整體趨勢為常規復合微生物菌劑處理優于滅菌復合微生物菌劑。與CK相比,常規和滅菌復合微生物菌劑均顯著提高了不同測定時期棉花葉片SPAD值,35 d時效果最明顯,與CK相比,T1、T2、T3和T4處理分別顯著提高了11.02%、16.53%、15.82% 和13.17%;在25 d時,僅常規復合微生物菌劑顯著提高了棉花株高和莖粗,不同用量下差異不顯著;隨著處理時間的推移和次數的增多,到35 d時,滅菌復合微生物菌劑用量達10 L/667m2時,其株高和莖粗開始顯著高于CK;直至處理45 d后,不同用量的滅菌復合微生物菌劑均顯著提高了棉花株高和莖粗;此外,在棉花整個苗期生育階段,常規復合微生物菌劑處理的株高和莖粗均顯著高于CK處理。
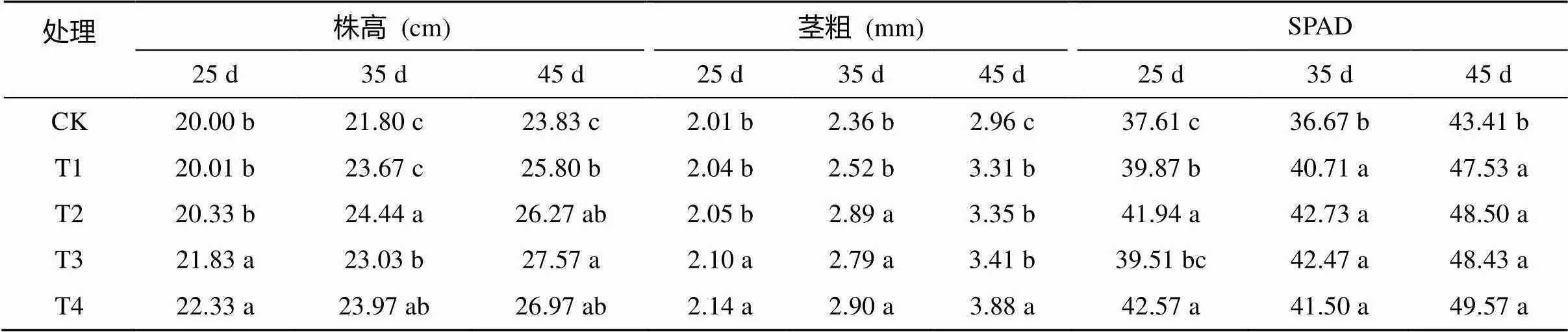
表1 不同處理對棉花株高、莖粗和SPAD值的影響
注:同列數據小寫字母不同表示處理間差異達到<0.05 顯著水平,下同。
2.2 復合微生物菌劑對棉花葉片內光合色素含量的影響
處理35 d后取棉花倒4葉測定葉片內光合色素含量,由表2可知,與CK相比,常規和滅菌復合微生物菌劑均顯著提高了棉花葉片內葉綠素a和葉綠素b含量以及葉綠素a/b,對類胡蘿卜素含量的影響不顯著;其中,T1、T2、T3和T4的葉綠素a分別提高了21.60%、19.20%、25.60% 和36.00%,葉綠素b分別提高了18.60%、16.28%、13.95% 和25.58%,葉綠素a/b分別提高了25.98%、20.08%、23.62% 和31.50%。以上結果表明常規復合微生物菌劑對光合色素的提高能力優于滅菌復合微生物菌劑,但不同用量的復合微生物菌劑對光合色素含量的影響差異較小。
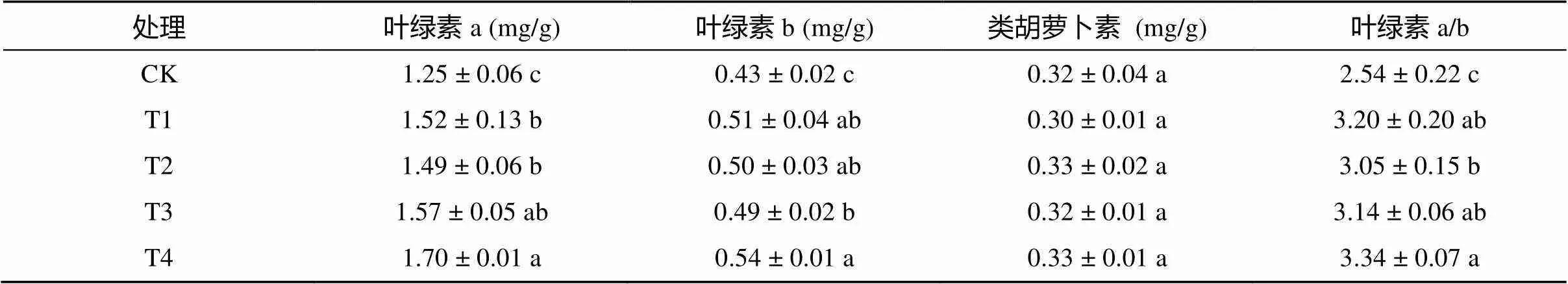
表2 不同處理對棉花葉片內光合色素含量的影響
2.3 復合微生物菌劑對棉花生物量的影響
棉花幼苗生長45 d后收獲測得幼苗植株生物量如表3所示,復合微生物菌劑可顯著提高棉花幼苗地上部和地下部干、鮮物質量,且干、鮮物質量的變化趨勢相同。棉花地上部生物產量隨復合微生物菌劑用量的增多而增多,而地下部受復合微生物菌劑用量的影響不明顯。等量條件下,常規復合微生物菌劑處理的棉花植株地上部與地下部生物量均顯著高于滅菌復合微生物菌劑,T3與T1 相比,地上部鮮物質量、干物質量,地下部鮮物質量、干物質量分別顯著提高了50.50%、63.20%和5.10%、12.09%;T4與T2相比也分別顯著提高了17.76%、18.26% 和7.28%、11.96%。表明復合微生物菌劑可促進幼苗期棉花生物量的累積,進而為后期生殖生長提供充足的物質保障。
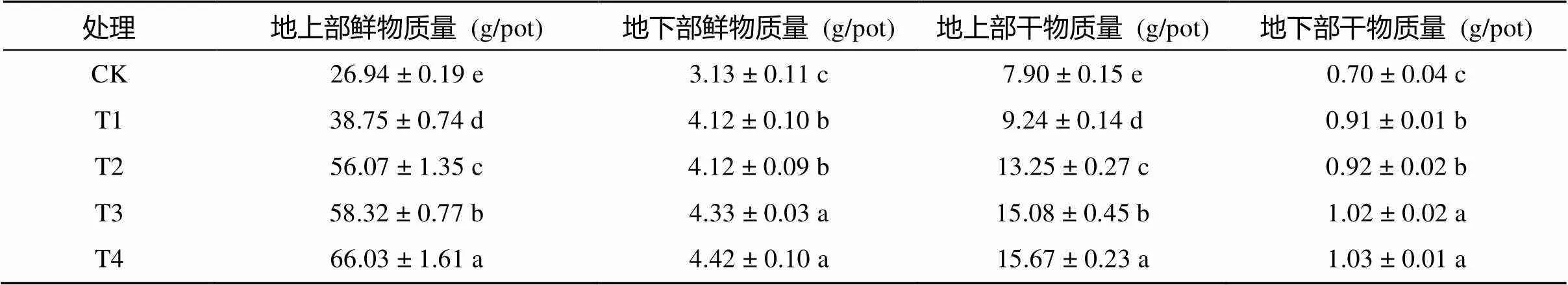
表3 不同處理對棉花幼苗生物量的影響
2.4 復合微生物菌劑對棉花葉片和根系內抗氧化物酶活性的影響
由圖1可見,復合微生物菌劑顯著提高了棉花葉片和根系內SOD、POD和CAT活性,且各處理葉片SOD和CAT活性高于根系,而POD活性與之相反。由圖1A可知,常規復合微生物菌劑對葉片和根系內SOD活性的提高能力較滅菌復合微生物菌劑明顯,T3較T1分別顯著提高了3.41% 和37.58%,T4較T2分別顯著提高了4.14% 和19.56%,這表明常規復合微生物菌劑對根系SOD活性的提高能力大于葉片,而不同用量間的差異不顯著。由圖1B可見,POD活性變化趨勢與SOD相似,與T1相比,T3處理葉片和根系內POD活性分別顯著提高了44.43% 和3.29%;而T4與T2相比,僅葉片內顯著提高了8.61%,根系內差異不顯著,這表明常規復合微生物菌劑對葉片POD活性的提高能力較根系明顯,然而用量升高后酶活性卻有所降低,但差異不顯著。由圖1C可見,與滅菌復合微生物菌劑處理相比,常規復合微生物菌劑處理降低了葉片CAT活性,差異不顯著;而根系內活性顯著提高,T3比T1提高了98.46%,T4比T2提高了63.96%,這表明常規復合微生物菌劑對根系CAT活性的提高能力較葉片明顯,且隨用量的升高而升高。因此復合微生物菌劑可通過提高棉花體內抗氧化物酶的活性來緩解逆境脅迫下的氧化損傷,提高植物抵抗逆境脅迫的能力,且常規復合微生物菌劑的效果優于滅菌復合微生物菌劑,而不同用量間差異不顯著。
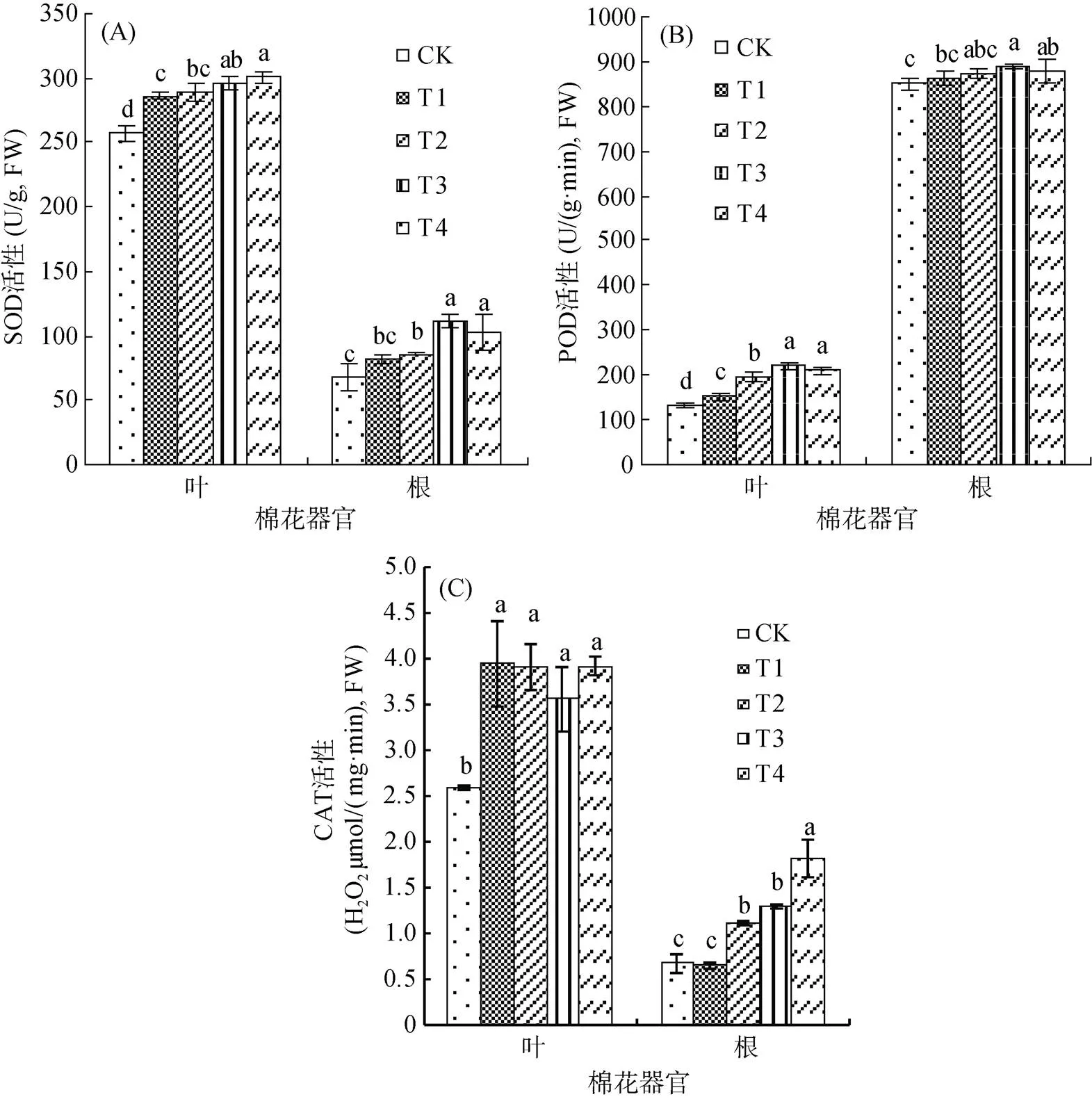
(圖柱上方小寫字母不同表示處理間差異達到P<0.05 顯著水平,下同)
2.5 復合微生物菌劑對棉花葉片和根系內過氧化物質含量的影響
植物在生長過程中遭受逆境脅迫時體內代謝平衡發生紊亂,會累積大量有害過氧化物質(MDA、H2O2、O2? ?),致使氧化還原平衡受損。由圖2可見,復合微生物菌劑顯著降低了棉花葉片和根系內MDA含量和超氧陰離子產生速率,但降低能力與復合微生物菌劑的種類和用量有關。由圖2A可見,葉片內,T1、T2和T3處理間差異不顯著,而T4與各處理相比存在顯著性差異,T4與T2相比,顯著降低了47.18%;根系內,滅菌條件下隨用量的升高,MDA累積量逐漸降低,常規條件下卻出現了相反的趨勢,T3比T1顯著降低了20.00%,而T4與T2相比有所升高,但差異不顯著。由圖2B可見,常規復合微生物菌劑對葉片和根系內超氧陰離子產生速率的降低能力高于滅菌復合微生物菌劑,且均隨用量的增加而降低,T3與T1相比,分別降低了14.16% 和22.34%,T4與T2相比,分別降低了11.93% 和5.91%,且僅根系內T3與T1間存在顯著性差異。以上結果表明,復合微生物菌劑可通過提高棉花體內抗氧化物酶的活性來促進過氧化物質的分解,降低植物體內過氧化物質的積累量,保證植物體內正常的氧化還原電位,減輕膜質氧化損傷,最終提高植物抵抗逆境脅迫能力。
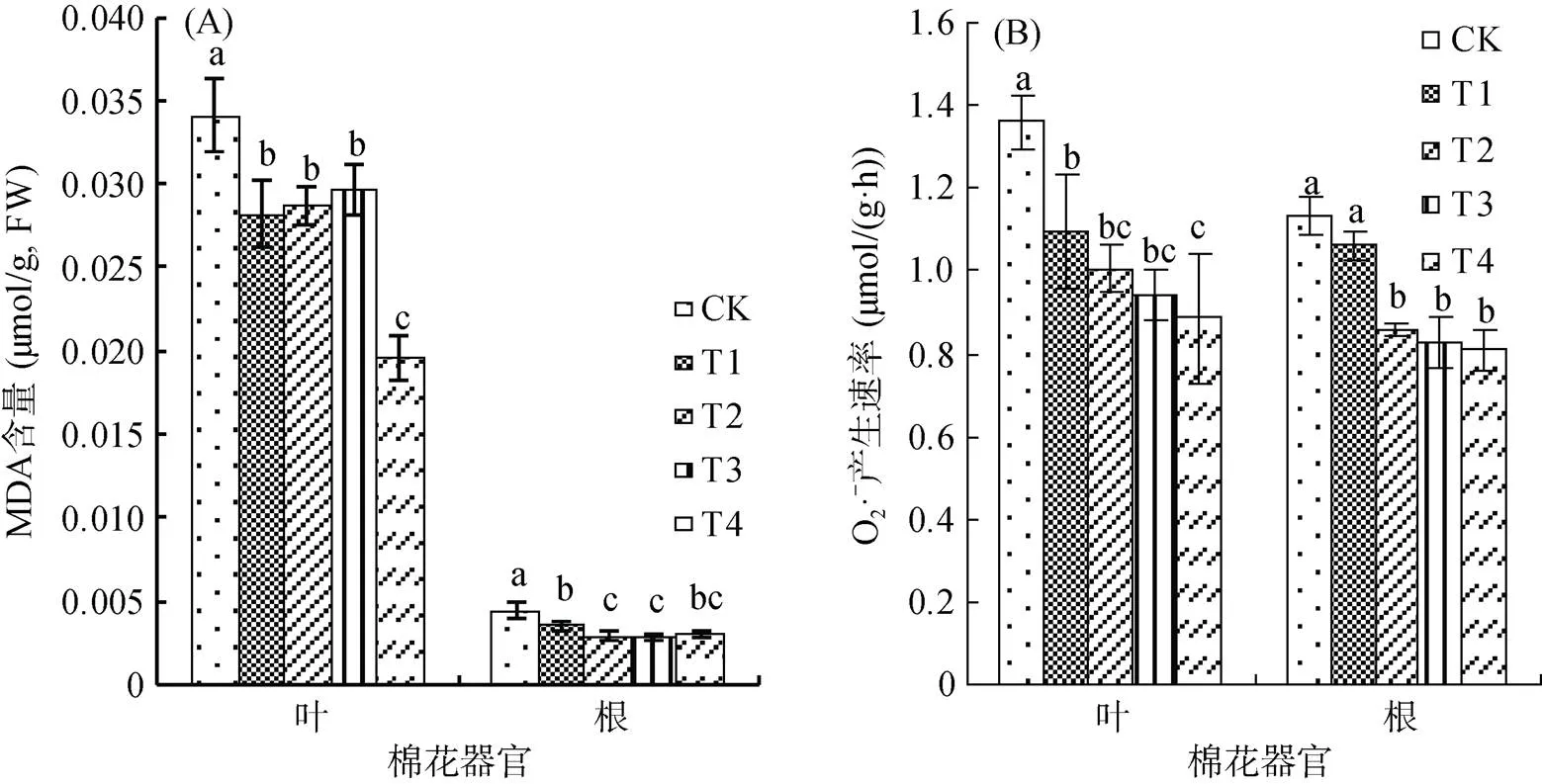
圖2 不同處理對棉花葉片和根系內丙二醛(MDA)含量和超氧陰離子(O2? ?)產生速率的影響
2.6 復合微生物菌劑對棉花根際土壤pH的影響
土壤酸堿度與根際微生物和根系分泌物息息相關,由圖3可見,復合微生物菌劑顯著降低了根際土壤pH,與CK相比,T1、T2、T3和T4分別降低了4.27%、12.07%、5.00% 和13.98%,即常規復合微生物菌劑的效果優于滅菌復合微生物菌劑,且用量對根際土壤pH的影響較菌劑是否滅菌更為顯著,當用量達到10 L/667m2時,土壤的pH降到了7.0 ~ 7.2,把土壤從堿性調到了中性。
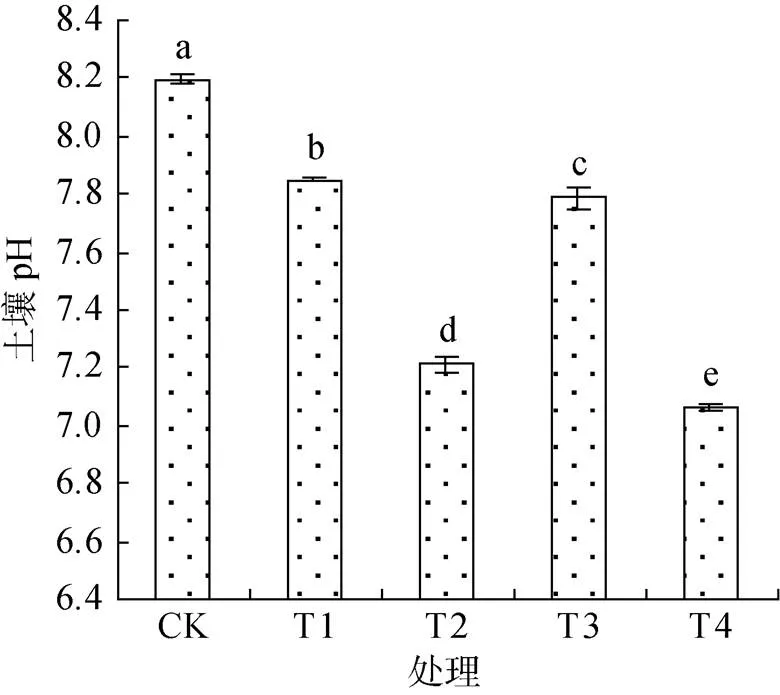
圖3 不同處理對棉花根際土壤pH的影響
2.7 復合微生物菌劑對棉花根際土壤微生物數量的影響
由圖4可見,各處理土壤細菌數>放線菌數>真菌數,添加復合微生物菌劑后土壤微生物總量呈上升趨勢。由圖4A可見,處理30 d時,與CK相比,除T1處理外,T2、T3和T4處理的根際土壤細菌數分別顯著提高了19.05%、30.95% 和103.10%;到處理45 d時,與CK相比,各處理分別顯著提高了22.42%、19.89%、48.28% 和58.59%,均高于30 d的細菌數,且常規復合微生物菌劑與滅菌復合微生物菌劑間存在顯著性差異,而不同用量間差異不顯著。由圖4B可見,復合微生物菌劑處理降低了根際土壤真菌數,處理30 d時,T1和T4與CK相比,顯著降低了10.17% 和23.38%;到處理45 d時,除T2外,其余各處理與CK相比,分別顯著降低了10.53%、11.99% 和23.82%,與30 d相比,各處理真菌數均有所降低,且兩次結果均為復合微生物菌劑用量越高,真菌數越低,等量條件下,常規復合微生物菌劑的根際土壤真菌數低于滅菌復合微生物菌劑。由圖4C可見,復合微生物菌劑提高了根際土壤放線菌數,處理30 d時,與CK相比,除T1顯著提高了38.17%,其余處理間差異不顯著;而到45 d時,除T4與CK相比差異不顯著外,其余處理顯著提高了13.11%、17.10% 和20.14%。以上結果表明根際土壤細菌數和真菌數受常規復合微生物菌劑的影響效果較滅菌復合微生物菌劑明顯,且用量越高效果越顯著,而放線菌數的變化趨勢與之相反。
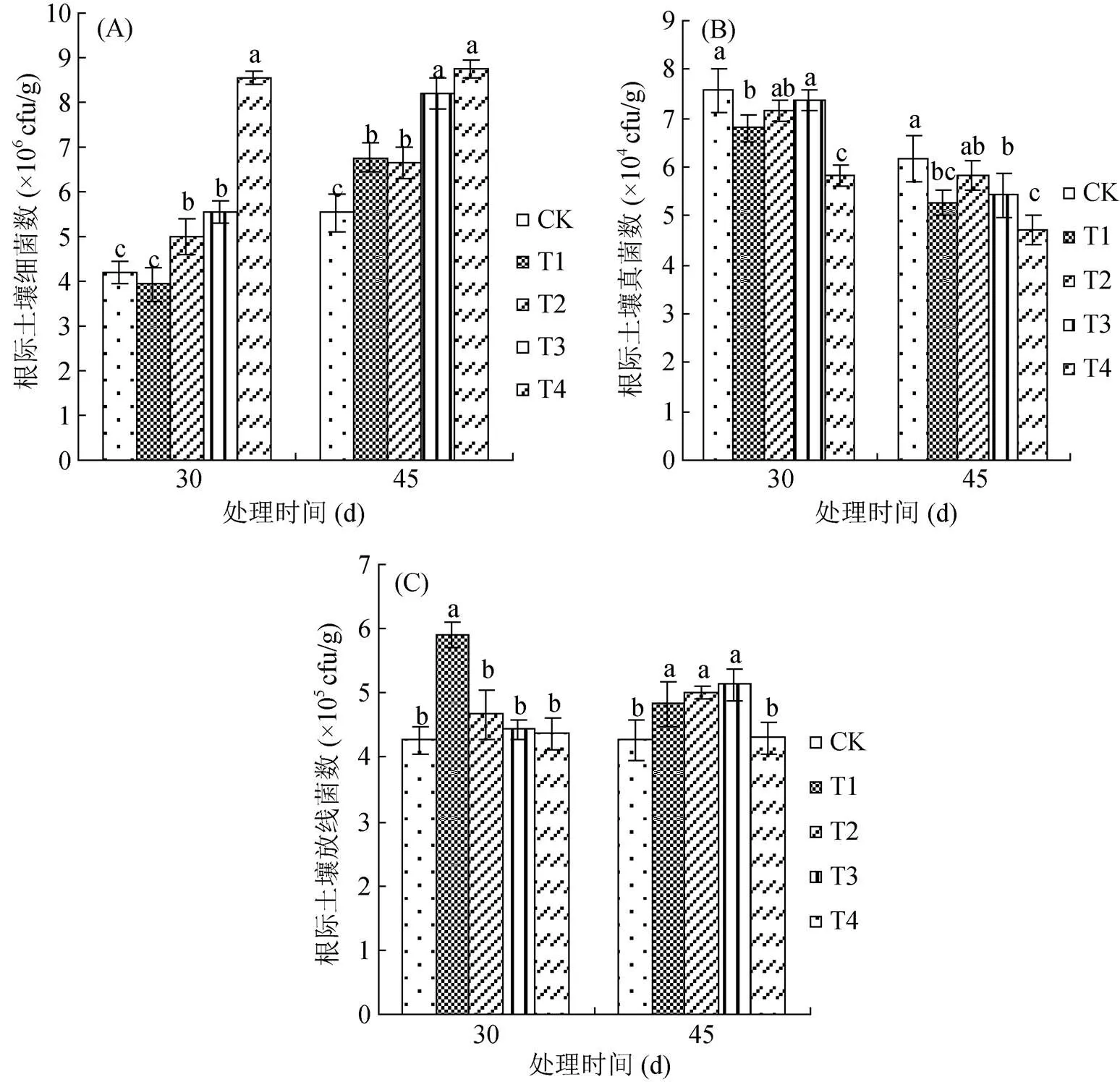
圖4 不同處理對棉花根際土壤細菌(A)、真菌(B)和放線菌數(C)的影響
2.8 復合微生物菌劑對棉花根際土壤酶活性的影響
由表4可知,復合微生物菌劑提高了土壤脲酶、脫氫酶、中性磷酸酶和蔗糖酶活性,除脲酶外,其余各酶活性與CK相比,均存在顯著性差異。除T2外,T1、T3和T4脲酶活性與CK相比,也分別顯著提高了7.55%、11.32% 和5.67%,即復合微生物肥料用量越大,對脲酶活性的提高能力越小。與CK相比,T1、T2、T3和T4的土壤脫氫酶活性分別提高了7.91%、15.42%、20.56% 和21.34%,土壤中性磷酸酶活性分別提高了36.69%、40.77%、42.47% 和67.61%,土壤蔗糖酶活性分別提高了24.64%、67.91%、37.31% 和82.09%,而土壤過氧化氫酶活性卻顯著降低了3.22%、6.44%、4.51%和6.77%。以上結果表明等量條件下常規復合微生物菌劑對土壤各酶活性的影響較滅菌復合微生物菌劑明顯,脫氫酶、中性磷酸酶和蔗糖酶活性均隨復合微生物菌劑用量的升高而升高,而土壤脲酶和過氧化氫酶活性的變化趨勢與之相反。
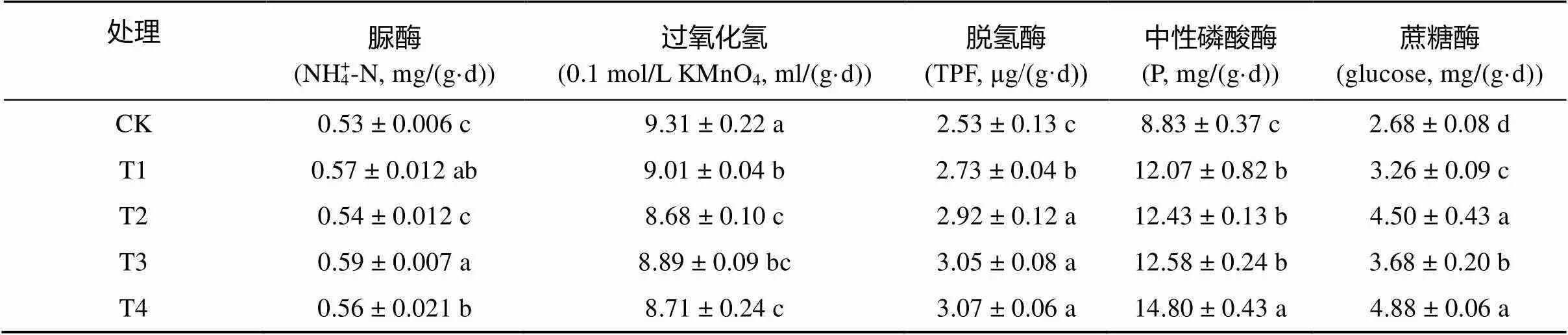
表4 不同處理對棉花根際土壤酶活性的影響
2.9 復合微生物菌劑對棉花根際土壤養分含量的影響
由表5可知,復合微生物菌劑提高了土壤養分及有機質含量,除堿解氮外,其余養分含量與CK相比均存在顯著性差異,其中T1、T2、T3和T4的全氮含量分別提高了32.84%、76.12%、55.22%和67.16%,速效鉀分別提高了8.99%、50.16%、5.07% 和41.33%,有效磷分別提高了15.30%、13.07%、7.32%和12.42%,有機質分別提高了5.52%、15.13%、10.94% 和9.41%,除T1外,堿解氮含量也分別顯著提高了33.28%、8.15% 和18.57%。這表明土壤養分含量隨復合微生物菌劑用量的增加而增加,且等量條件下,滅菌復合微生物菌劑對土壤速效鉀和有效磷的提高比例高于常規復合微生物菌劑,當用量為10 L/667m2時,堿解氮、全氮和有機質含量也表現出了相同的變化趨勢。
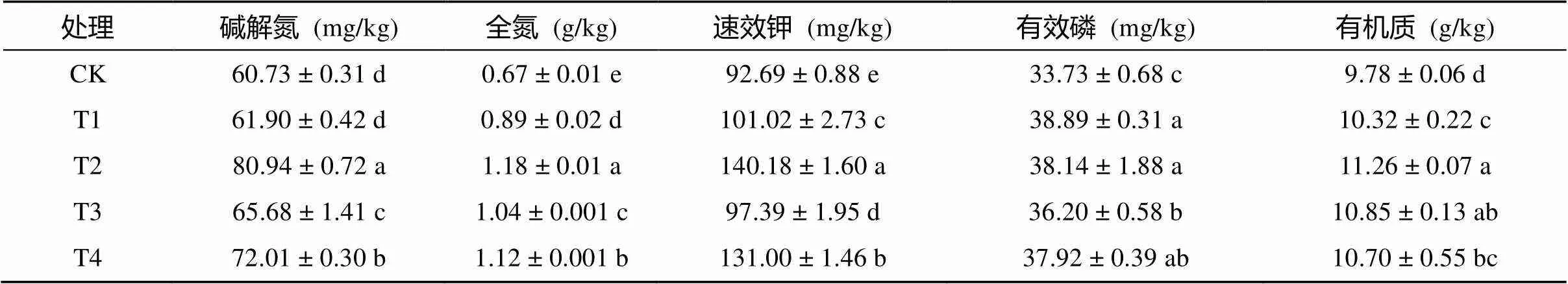
表5 不同處理對棉花根際土壤養分含量的影響
3 討論
在自然生態系統中,80% 以上的高等植物和微生物菌種處于共生關系,微生物可通過在土壤中形成龐大的菌絲網絡來增加植物根系的吸收面積,從而增加根系對營養元素的吸收進而促進植物生長[20]。本研究發現復合微生物菌劑提高了幼苗期棉花葉片內光合色素總量和SPAD值,提高棉花的光合能力,促進碳水化合物的合成和轉運,進而提高了棉花的株高和莖粗。因此,處理45 d后收獲棉花幼苗發現復合微生物菌劑用量越多,棉花地上部生物量越多,而地下部生物量受復合微生物菌劑用量的影響不明顯;此外,常規復合微生物菌劑添加量達10 L/667m2時,效果最顯著。王其傳等[21]研究發現,微生物菌劑可同時提高辣椒葉片葉綠素總量和凈光合速率( Pn),提高光合電子傳遞效率和植株凈光合能力,最終促進辣椒生長,提高產量;弓新國等[22]研究也發現苗期和大田期施用 FZB42 菌劑均可促進烤煙生長,減少病害發生,提高煙葉產量、質量。常規復合微生物菌劑對各指標的影響優于滅菌復合微生物菌劑,可能是常規復合微生物菌劑在為棉花生長提供所需養分的同時還含有大量高活菌,這些活菌在發酵過程中可產生各種有利于植物生長的代謝產物,如赤霉素(GA)、細胞分裂素(CTK)和吲哚乙酸(IAA)等促進植株生長的植物生長調節劑以及各類維生素、氨基酸和有機酸類物質,這些代謝產物對植物生長發育具有一定的調節促進作用[23-24];此外,微生物的活動和其代謝產物還可進一步活化土壤養分,改良土壤酸堿度和土壤通氣性,進而提高土壤的保水保肥能力,最終促進苗期棉花的營養生長,為后期生殖生長和養分累積提供充足的物質保障。何瑞成等[25]研究表明,土壤微生物數量和活性與水稻有效穗數、每穗總粒數、千粒重以及產量具有很強的正相關性,改善土壤環境的同時提高作物產量。
植物體內防御酶活性與植物抗性關系密切,前人研究表明,微生物菌劑在提高小麥產量的同時,還能顯著降低小麥紋枯病、根腐病和全蝕病的發病率[26]。本研究發現復合微生物菌劑可顯著提高棉花葉片和根系內SOD、POD和CAT活性,降低MDA含量和超氧陰離子產生速率,常規復合微生物菌劑效果優于滅菌復合微生物菌劑,且用量為10 L/667m2的效果優于5 L/667m2。這表明復合微生物菌劑可通過提高棉花體內抗氧化物酶的活性來促進過氧化物質的分解,降低植物體內過氧化物質的累積量,保證體內正常的氧化還原電位,減輕膜質氧化損傷,最終提高植物抗逆境脅迫能力。陳沖等[27]研究同樣發現施用復合微生物菌劑可提高煙草葉片內SOD和POD活性,降低MDA的含量,提高煙草的抗逆性。這可能是因為復合微生物菌劑所含的大量有益活菌物質和多種天然發酵活性物質能夠在根區土壤繁殖形成有利于作物生長的微生物優勢菌群,這些菌群在繁殖代謝過程中優化根際微域環境,促進植物根系對營養物質的吸收和生長,進而增強植物抵抗逆境脅迫的能力。常規復合微生物菌劑處理優于滅菌復合微生物菌劑的原因,一方面可能是常規復合微生物菌劑在促進植物對氮、磷、鉀等大量元素吸收利用的同時,還能通過微生物的分泌代謝活化提高土壤中難溶不易被作物吸收的Fe、Mn、Cu、Zn等微量元素的生物有效性,而POD 和 CAT 是典型的亞鐵血紅素酶,所以其活性與Fe含量高低有很強的相關性[28],Cu、Zn、Mn 和 Fe是 SOD 活性中心的金屬元素[29],因而可通過提高棉花體內微量元素含量來進一步激活其根系和葉片內的抗氧化物酶活性;另一方面常規復合微生物菌劑中的功能菌可作為誘導因子,激活并增強植株體內抗氧化物酶以及與抗病相關蛋白的活性來提高植物的抗性[30],因此遭受外界不良環境影響時,植物體會通過外界的信號源,激發自身已有的強大調控系統來抵御不良環境。
土壤微生物活性的強弱關系土壤新陳代謝的快慢,土壤內各種生化反應都離不開微生物的參與,微生物菌劑可通過改善土壤或基質的理化性質,促進植物生長,提高產量,改善品質[31-32]。已有研究表明,在黃瓜不同生育期施用枯草芽胞桿菌 BSD2、放線菌 S-317、木霉 T36 等多種有益微生物配成液態復合制劑后降低了根際真菌數,而細菌和放線菌數明顯提高,改善土壤微生物的群落結構和土壤營養條件,增強植株的抗逆能力[33];復合微生物菌劑還可通過降低反硝化細菌、真菌數量和提高硝化細菌、氨化細菌、固氮菌、纖維素分解菌、放線菌數量來改變土壤生態環境[34]。本研究發現施入復合微生物菌劑后土壤微生物總量呈上升趨勢,隨處理時間的推移土壤細菌數逐漸增多,且常規復合微生物菌劑處理顯著高于滅菌復合微生物菌劑處理,而根際土壤真菌數的變化趨勢與之相反;同時提高了根際土壤放線菌數,且在處理30 d時,僅滅菌復合微生物菌劑用量為5 L/667m2時效果顯著,并隨時間的推移,除常規復合微生物菌劑用量為10 L/667m2處理外,其余處理均顯著高于CK,這與孔慶宇等[35]和尹淑麗等[36]研究結果相似。曹云等[37]研究發現在棉隆熏蒸結合微生物有機肥處理土壤的微生物數量和酶活性恢復速度和強度較普通有機肥處理強,病原菌數量及病原菌與真菌數量比值低,微生物碳源利用多樣性指數最高。出現上述結果的原因,首先復合微生物菌劑含有大量氨基酸、多肽、單糖和多糖等有機成分和氮、磷、鉀等營養成分,可為土著微生物的生長繁殖提供所需的氮源和能源[38];其次,常規復合微生物菌劑還含有大量有益微生物,施入土壤后在棉花根際定殖,從而提高根際微生物的豐富度和多樣性,因此常規復合微生物菌劑處理的根際微生物總量高于滅菌復合微生物菌劑;第三,微生物在生長繁殖過程中既存在互利共生也存在相互競爭的作用,當根際微域的某一群落變為優勢種群時,必然會影響到其他群落的生長繁殖,所以當根際土壤細菌數顯著增多時,其真菌數會有所降低。第四,土傳病原菌主要為真菌型病原菌,常規復合微生物菌劑中所含的芽孢類桿菌、乳酸菌和酵母菌等多種有益菌可通過自身的代謝分泌來殺死根際土壤中原有病原菌,同時其分泌物可在植物根系形成保護膜,防止病原菌的入侵,減輕土傳病害,降低根際土壤有害病原菌數[39],最終把“真菌型”土壤轉化為“細菌型”土壤。
土壤酶是土壤有機質分解與養分轉化和循環的驅動力,是土壤質量和生態穩定性的重要指標[40],不但影響土壤養分有效性,還能促進作物的生長發育和根系活力的提高[41]。已有研究表明,微生物菌劑的施用會影響土壤酶活性、土壤生化反應、土壤養分循環和作物對養分的吸收利用能力[42-43]。本研究發現,施入復合微生物菌劑提高了土壤脲酶、脫氫酶、中性磷酸酶和蔗糖酶活性,但土壤脲酶活性隨肥料用量的增多而降低,其余酶活性均隨肥料用量的增多而升高,而土壤過氧化氫酶活性卻有所降低;此外,常規復合微生物菌劑對土壤各酶活性的影響較滅菌復合微生物菌劑明顯。張美存等[44]研究表明,向草坪土壤基質中施入放線菌、枯草芽孢桿菌和蘇云金芽孢桿菌均可顯著提高土壤蔗糖酶、脲酶、多酚氧化酶和過氧化物酶活性,本研究結果與此相似;曹恩琿等[45]研究也表明復合微生物菌劑在降低番茄土壤酸度,增強土壤脲酶、磷酸酶、纖維素酶及過氧化氫酶活性的同時,還顯著提高了土壤有機質和有效養分含量。此外,土壤水分、有機質、氮素和有效磷等物質也是影響土壤微生物群落結構的最主要環境因素[46-47]。本研究發現土壤養分含量隨復合微生物菌劑施入量的增加而增加,且滅菌復合微生物菌劑對土壤速效鉀和有效磷的提高比例高于常規復合微生物菌劑。這首先是復合微生物菌劑本身含有大量的養分和有機質,施入土壤后提高了土壤養分及有機質含量,因而土壤養分含量隨復合微生物菌劑施入量的增加而增加;其次,在充足的氮源、碳源供應下又加速了根際土壤微生物的繁殖和代謝,進而提高了微生物分泌物和生理活性物質的產生,在提高土壤酶活性的同時活化土壤中難溶不易移動的營養元素,以此提高土壤養分有效性[48]。然而常規復合微生物菌劑對土壤各酶活性的影響較滅菌復合微生物菌劑明顯的原因可能是:復合微生物菌劑中的高活性微生物改善了土壤微環境,一定程度上激活了土壤中的土著微生物,并且常規復合微生物菌劑對根際土壤微生物豐富度和多樣性的提高能力優于滅菌型復合微生物菌劑,這些微生物在繁殖代謝過程中加速有機化合物的分解,提供了酶促反應的底物,進而提高了胞外酶釋放量和與提高酶活性有關物質的分泌量[48]。土壤脲酶活性隨肥料用量的增多而降低的原因可能是:當肥料用量增多時,土壤供氮水平提高,為了減少氮素損失提高氮素利用率,微生物可通過改變自身的代謝分泌及根系的分泌來調整脲酶的活性。滅菌復合微生物菌劑處理的養分和有機質含量高于常規復合微生物菌劑的原因首先可能是高溫滅菌時,復合微生物菌劑中有部分水分的蒸發,使養分含量升高;其次,棉花在生長過程中需要不斷從土壤中吸收水分和營養物質,并且本試驗條件下,常規復合微生物菌劑處理的棉花生物量顯著高于滅菌復合微生物菌劑處理,因而加大了對土壤養分的索取,所以收獲后土壤養分含量相對較低。
4 結論
常規和滅菌復合微生物菌劑均可提高幼苗期棉花葉片內光合色素含量,葉片和根系內SOD、POD和CAT等抗氧化物酶活性,降低MDA和O2??等過氧化物質累積,進而提高棉花的光合性能和抗氧化損傷能力,提高植物抗逆性,促進碳水化合物的合成、轉運和累積,最終提高了棉花的株高、莖粗和生物量的累積,常規復合微生物菌劑處理效果優于滅菌復合微生物菌劑,但不同用量間的差異較小。
常規和滅菌復合微生物菌劑均提高了根際土壤細菌數和放線菌數,且隨處理時間的推移,其效果越明顯,其中細菌數隨用量的升高而增多,但放線菌數的變化與之相反,而根際土壤真菌數顯著降低。因此改善了根際微生物生態平衡,優化了根際微域環境,促進了微生物和根系分泌,提高了土壤脲酶、脫氫酶、中性磷酸酶和蔗糖酶活性,進而提高了土壤有效氮、磷、鉀和有機質含量,最終促進了棉花苗期生長和生物量的累積,為后期的生殖生長提供充足的物質保障。
[1] 葉邦興, 唐海明, 湯小明, 等. 中國農田污染的現狀及防治對策初探[J]. 中國農學通報, 2010, 26(7): 295–298
[2] 張雪峰,胡濱. 微生物肥料對土壤生態環境修復效應的影響[J]. 綠色科技, 2011(2): 98–100
[3] 郝彩琴, 劉美杰, 陳海燕. 菊科植物生物活性研究概述[J]. 西北林學院學報, 2010, 25(2): 45–49
[4] 張曼麗, 陳劍山, 柳曉磊, 等. 水溶肥配施微生物菌劑對海南設施哈密瓜產量及土壤的影響[J]. 中國農技推廣, 2017, 33(2): 54–56
[5] Spadaro D, Gullino M L. Improving the efficacy of biocontrol agents against soilborne pathogens[J]. Crop Prot, 2005, 24(7): 601–613
[6] 丁莉萍, 孫維紅, 李朋飛. 復合菌劑對樟樹苗移栽生長及圃地土壤的影響[J]. 生物技術進展, 2017, 7(3): 236– 240
[7] 肖蓉, 孫海峰, 聶園軍, 等. 復合微生物菌劑對重茬蘋果園土壤細菌群落的影響[J]. 微生物學通報, 2016, 43(2): 301–309
[8] 趙亞慧, 華雪潔, 杜海巖, 等. 花生化感物質降解菌和抗連作拮抗菌復合菌劑應用效果的研究[J]. 土壤通報, 2016, 47(3): 599–604
[9] 逄煥成, 李玉義, 嚴慧峻, 等.微生物菌劑對鹽堿土理化和生物性狀影響的研究[J]. 農業環境科學學報, 2009, 28(5): 951–955
[10] 馮國藝, 張謙, 雷曉鵬, 等. 濱海鹽堿地不同適性棉花品種的篩選及其性狀特征[J]. 河北農業科學, 2017, 21(3): 25–29
[11] 孫凱寧, 于君寶, 王克安, 等. 不同肥料類型對濱海鹽堿地棉花生長及肥料利用率的影響[J]. 中國農學通, 2015, 31(15): 91–96
[12] 孫中濤, 姚良同, 孫鳳鳴, 等. 微生物肥料對棉田土壤生態與棉花生長的影響[J]. 中國生態農業學報, 2005, 13(3): 54–56
[13] Knudson L L, Tibbitts T W, Edwards G E. Measurement of ozone injury by determination of leaf chlorophyll concentration[J]. Plant Physiology, 1977, 60(4): 606–608
[14] 李合生.植物生理生化實驗原理和技術[M].北京: 高等教育出版社, 2000: 167–169
[15] Heath R L, Packer L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation[J]. Archives Biochemistry and Biophysics1968, 125(1): 189–198
[16] Shi Q H, Zhu Z J. Effects of exogenous salicylic acid on manganese toxicity, element contents and antioxidative system in cucumber[J]. Environmental and Experimental Botany, 2008, 63(3): 317–326
[17] 何紹江, 陳雯莉.微生物實驗[M]. 北京: 中國農業出版社, 2007: 45–48
[18] 鮑士旦. 土壤農化分析[M]. 北京: 中國農業出版社, 2005: 14–114
[19] 關松蔭. 土壤酶及其研究法[M]. 北京:農業出版社, 1986: 294–297
[20] 陳梅梅, 陳保冬, 王新軍,等.不同磷水平土壤接種叢枝菌根真菌對植物生長和養分吸收的影響[J]. 生態學報, 2009, 29(4): 1980–1986
[21] 王其傳, 孫錦, 束勝, 等. 微生物菌劑對日光溫室辣椒生長和光合特性的影響[J]. 南京農業大學學報, 2012, 35(6): 7–12
[22] 弓新國, 胡曉峰, 陳俊秋, 等. 不同時期施用微生物菌劑對烤煙生長和產量質量的影響[J].中國土壤與肥料, 2014(6): 106–110
[23] 陳慧君. 微生物肥料菌種應用與效果分析[D]. 北京: 中國農業科學院, 2013.
[24] Vessey J K. Plant growth promoting rhizobacteria as biofertilizers[J]. Plant soil, 2003, 255(2): 571–586
[25] 何瑞成, 吳景貴. 有機物料對原生鹽堿地土壤生物學性質的影響[J]. 土壤學報, 2018, 55(3): 774–782
[26] 常娜, 張雪嬌, 馬璐璐, 等.微生物菌劑對小麥生長及土傳病害預防效果的影響[J]. 作物雜志, 2017, 176(1): 155–160
[27] 陳沖, 王小東, 郭傳濱, 等. 微生物菌劑不同用量對烤煙生長和葉片關鍵酶活性的影響[J]. 山東農業科學, 2015, 47(9): 73–76
[28] Ranieri A, Castagna A, Baldan B, et al. Iron deficiency differently affects peroxidase isoforms in sunflower[J]. Journal of Experimental Botany, 2001, 52, 354: 25–35
[29] Sevilla F, del Rio L A, Hellin E. Superoxide dismutases from a citrus plant: Presence of two iron-containing isoenzymes in leaves of lemon trees (L) [J]. Journal of Plant Physiology, 1984, 116(5): 381–387
[30] 丁傳雨, 喬煥英, 沈其榮, 等.生物有機肥對茄子青枯病的防治及其機理探討[J]. 中國農業科學, 2012, 45(2): 239–245
[31] 常勃. 微生物菌劑對礦區復墾土壤生物活性和油菜生長的影響[D]. 太原: 山西大學, 2013
[32] 侯樂梅, 孟瑞青, 乜蘭春, 等. 不同微生物菌劑對基質酶活性和番茄產量及品質的影響[J]. 應用生態學報, 2016, 27(8): 2520–2526
[33] 麻耀華, 尹淑麗, 張麗萍, 等.復合微生態制劑對黃瓜根際土壤微生物數量和酶活性的影響[J]. 植物保護, 2012, 38(2): 46–50
[34] 葛紅蓮, 李亞茹. 復合菌肥對黃瓜根際微生物區系的影響[J].農村經濟與科技, 2017, 28(9): 60–61
[35] 孔慶宇, 秦嗣軍, 張英霞, 等. EM菌劑對甜櫻桃幼苗根際微生物區系及根系呼吸的影響[J]. 沈陽農業大學學報, 2013, 44(4): 409–412
[36] 尹淑麗, 張麗萍, 張根偉, 等.復合微生態菌劑對黃瓜根際土壤微生物數量及酶活的影響[J]. 微生物學雜志, 2012, 32(1): 23–27
[37] 曹云, 宋修超, 郭德杰, 等.棉隆熏蒸與微生物有機肥聯用對西瓜枯萎病的防控研究[J]. 土壤,2018, 50(1): 93–100
[38] 陳劍山, 李鵬, 張曼麗, 等.氨基酸水溶肥與微生物菌劑混用抑制豇豆枯萎病的效果[J]. 中國植保導刊, 2015, 35(8): 52–53
[39] 雷先德, 李金文, 徐秀玲, 等.微生物菌劑對菠菜生長特性及土壤微生物多樣性的影響[J]. 中國生態農業學報, 2012, 20(4): 488–494
[40] DeForest J L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA[J]. Soil Biology and Biochemistry, 2009, 41(6): 1180–1186
[41] 朱金峰, 王小東, 郭傳濱, 等. 施用微生物菌劑對土壤關鍵酶活性和烤煙根系生長的影響[J]. 江西農業學報, 2015, 27(9): 31–35
[42] 曹慧, 孫輝, 楊浩, 等.土壤酶活性及其對土壤質量的指示研究進展[J]. 應用與環境生物學報, 2003, 9(1): 105– 109
[43] 杜廣紅, 周曉琳, 馬鳴超, 等. 不同施肥處理對土壤微生物學特性和作物產量的影響[J]. 中國土壤與肥料, 2012(5): 22–26
[44] 張美存, 程田, 多立安, 等. 微生物菌劑對草坪植物高羊茅生長與土壤酶活性的影響[J]. 生態學報, 2017, 37(14): 4763–4769
[45] 曹恩琿, 侯憲文, 李光義, 等. 復合菌劑對盆栽番茄土壤理化性質及微生物活性的影響[J]. 生態環境學報, 2011, 20(5): 875–880
[46] 楊東偉, 章明奎, 張鵬啟, 等. 水田改果園后土壤微生物學特性演變[J]. 土壤學報, 2018, 55(1): 182–193
[47] 吳松芹, 汪成忠, 李夢莎. 模擬氮沉降對濱海濕地土壤微生物功能多樣性的影響[J]. 土壤, 2017, 49(6): 1153– 1158
[48] 曹丹, 宗良綱, 肖峻, 等.生物肥對有機黃瓜生長及土壤生物學特性的影響[J]. 應用生態學報, 2010, 21(10): 2587–2592
Effects of Complex Microbial Agent on Cotton Physiological Characteristics, Microorganism and Physicochemical Properties in Rhizosphere Soil
SONG Yiling1,2, YU Jian1,2, CHEN Shigeng1, 2, XIAO Chengze1,2, LI Yuhuan3, SU Xiurong4, DING Fangjun1,2,3*
(1 Fertilizer Science & Technology Co. Ltd., Shandong Agricultural University, Taian, Shandong 271000, China; 2 Engineering & Technology Research Center of High Efficient Utilization of Humic Acid of Shandong Province, Taian, Shandong 271000, China; 3 College of Resources and Environment, Shandong Agricultural University, Taian, Shandong 271018, China; 4 College of Chemistry, Shandong Agricultural University, Taian, Shandong 271018, China)
To elucidate the effects and mechanism of complex microbial agent on the improvement of soilchemical and biological properties and the promotion of plant growth, the effects of different dosages of convention and sterilization complex microbial agents on growth, physiological metabolism of cotton and biological characteristics and nutrient contents in rhizosphere soil were investigated. Results showed that adding complex microbial agent increased chlorophyll content in cotton leaves, enhanced the activities of antioxidant enzymes (SOD, POD, CAT), reduced the contents of reactive oxygen species (MDA, O2? ?) in both leaves and roots, then increased photosynthetic performance and the ability to resist oxidative stress. Besides, it reduced soil pH, increased the amounts of soil bacteria and actinomycetes, decreased the number of soil fungi, increased the activities of soil urease, phosphatase, invertase and dehydrogenase, increased the contents of available nutrients and organic matter in soil, thus, improved the rhizosphere environment, and promoted cotton biomass accumulation and disease resistance. The above indexes except soil nutrient contents influenced by convention complex microbial agent were better than sterilization complex microbial agent, and the dosage of 10 L/667m2got the more obvious effect.In summary, the application of complex microbial agent could change soil enzyme activities and available nutrients by improving the number and structure of soil microbial community and soil pH, thus optimize the rhizosphere environment and increasing plant stress-resistance and photosynthetic ability, and consequently enrich soil and increase crop production. What’s more, the beneficial live bacteria in complex microbial agent had a particularly important positive effect on plant growth and soil improvement.
Complex microbial agent; Cotton; Antioxidant system; Soil microorganism; Soil enzyme
泰山產業領軍人才工程項目(LJNY201612)和山東省重點研發計劃項目(2015GNC110025)資助。
(sdndfyjs@163.com)
宋以玲(1989—),女,山東淄博人,碩士研究生,主要從事新型肥料的研發與應用研究。E - mail: songyiling0915@163.com
S144.1;S562
A
10.13758/j.cnki.tr.2019.03.009
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
