食源性致病菌等溫擴增檢測技術的研究進展
2019-07-27 07:07:18吳朦晨張明洲俞曉平
分析測試學報 2019年7期
關鍵詞:檢測
葛 航,吳朦晨,張明洲,俞曉平
(中國計量大學 生命科學學院 浙江省生物計量及檢驗檢疫技術重點實驗室,浙江 杭州 310018)
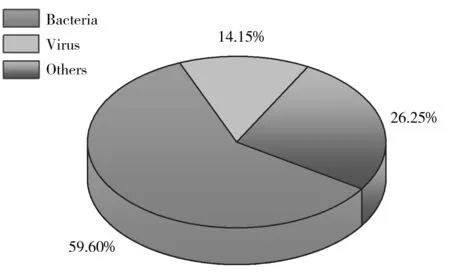
圖1 世界范圍內食源性風險因子危害程度分析[1]Fig.1 Global burden of diseases caused by food-borne factors[1]the percentage was calculated by dividing number of deaths caused by certain factor over total number of deaths
微生物與食品安全有著密切的關系,有益微生物被廣泛應用于食品制造與加工過程;有害微生物則會導致食品腐敗變質或直接危害人類健康,引起食品安全事件。世界衛生組織最新的食源性疾病調查報告顯示,全世界因食源性風險因子致病的人數超過6億人,死亡人數超過41萬人,其中因細菌導致死亡的人數超過24萬人,占總死亡人數的59.60%[1](圖1)。據國家衛生計生委統計,2011~2016年我國因微生物引起的食物中毒事件為2 446起,占食物中毒事件的25.5%,致病人數達4.82萬人,占總致病人數的43.3%,其中以腸炎沙門氏菌(Salmonellaenteritidis)和副溶血弧菌(Vibrioparahaemolyticus)為代表的食源性致病菌是引發食品安全事件的主要因子[2]。2018年8月25日發生在桂林的百人以上集體食物中毒事件,就是因參會人員食用了被沙門氏菌污染的食物所致。對食源性致病菌進行現場或在線快速精準檢測是保障公眾安全的重要手段,無論是對突發性食品安全事件的快速響應,或是對高危害性致病菌的有效防控,都需要相應的快速檢測方法作為支撐。然而,細菌微小的個體和復雜的種類,使得對其準確鑒定具有一定的困難,這也凸顯了開發快速、準確而又高效的致病菌檢測方法的必要性和迫切性。
1 食源性致病菌核酸檢測方法
微生物的傳統鑒別方法(培養法)一般基于其形態和生化特性,需要經過選擇性增菌、分離菌株、擴增培養、制片觀察形態等常規生化鑒定和毒素鑒定步驟。培養法雖然判定準確性高、穩定性強,但耗時通常為5~6天,且對檢驗者的經驗和技巧要求較高。因此,培養法在一些需要做出快速反應或檢測量大的現場或在線檢測中局限較大。除此之外,有些細菌能夠進入不可培養狀態(Viable but non culturable,VBNC),即菌體具有活性但無法被培養,這種情況會導致所檢測的細菌數量被低估[3]。因此,基于形態觀察和生化鑒定的傳統方法雖然可作為最終的仲裁方法,卻無法在有限的時間內完成對大量樣品的篩查,恰恰相反,這正是以聚合酶鏈式反應(Polymerase chain reaction,PCR)技術為代表的致病菌核酸檢測方法所具備的優點。
PCR方法以細菌特異性核酸序列作為檢測目標,簡化了判斷細菌種類的標準,并通過指數級的擴增,提高了檢測靈敏度。PCR技術的結果非常可靠,一方面,細菌的特征基因具有非常高的特異性,在其他細菌種類中可能不存在或者序列不同,即使是種間較為保守的基因,如16S基因,也在不同種類間具有多樣性的特定區域,序列完全相同的可能性極低;另一方面,通過DNA測序技術,能夠獲得詳細的序列堿基組成和排列信息,為判斷是否檢測到靶標基因提供可靠證據。同時,以核酸為靶標的檢測方法檢測通量高、檢測時間短,在檢測效率上有著明顯的優勢。由于PCR的以上諸多優點,相關研究在近十年呈現快速增長的趨勢(圖2),在國標中一些致病菌的檢測方法也采用了PCR方法(表1)。
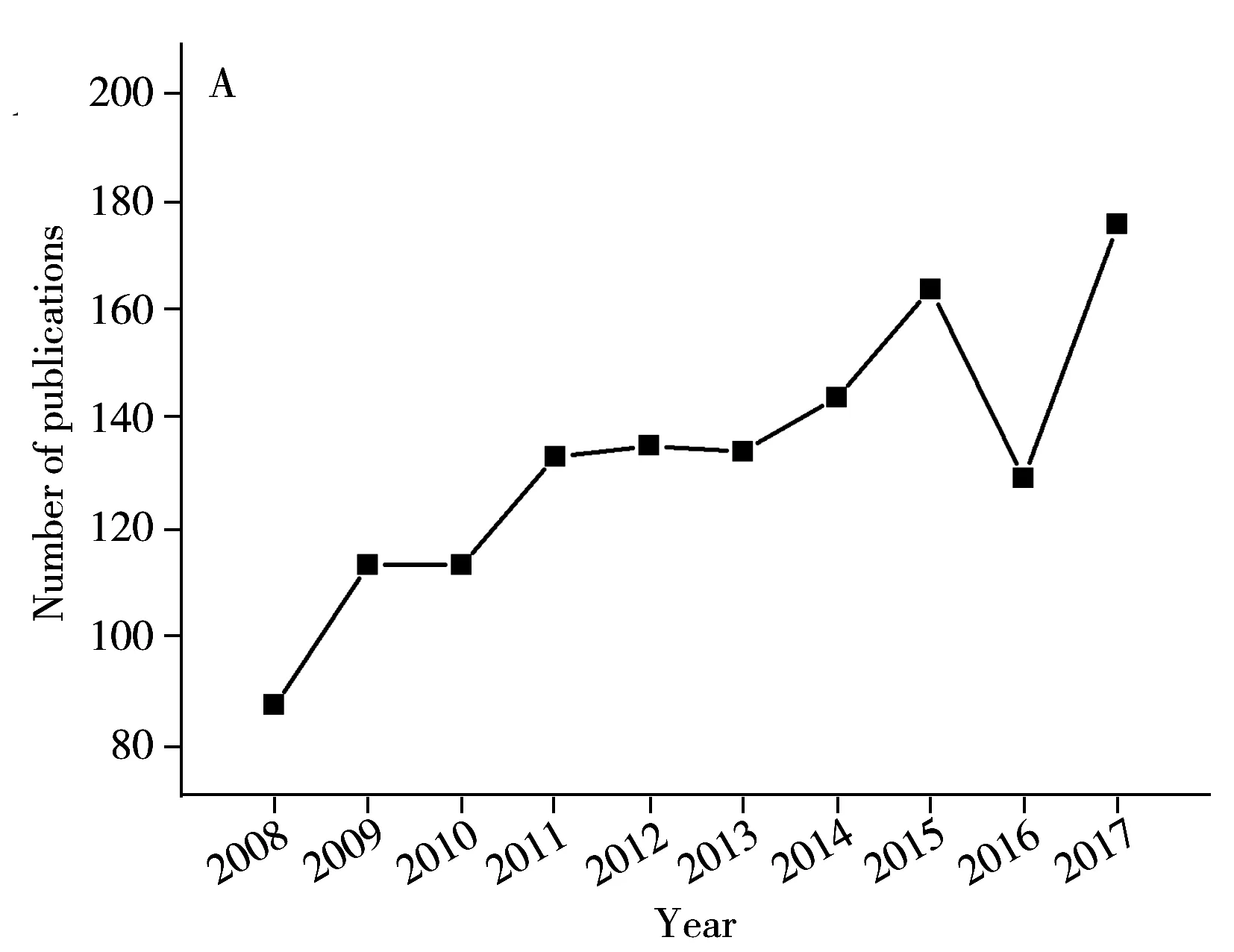
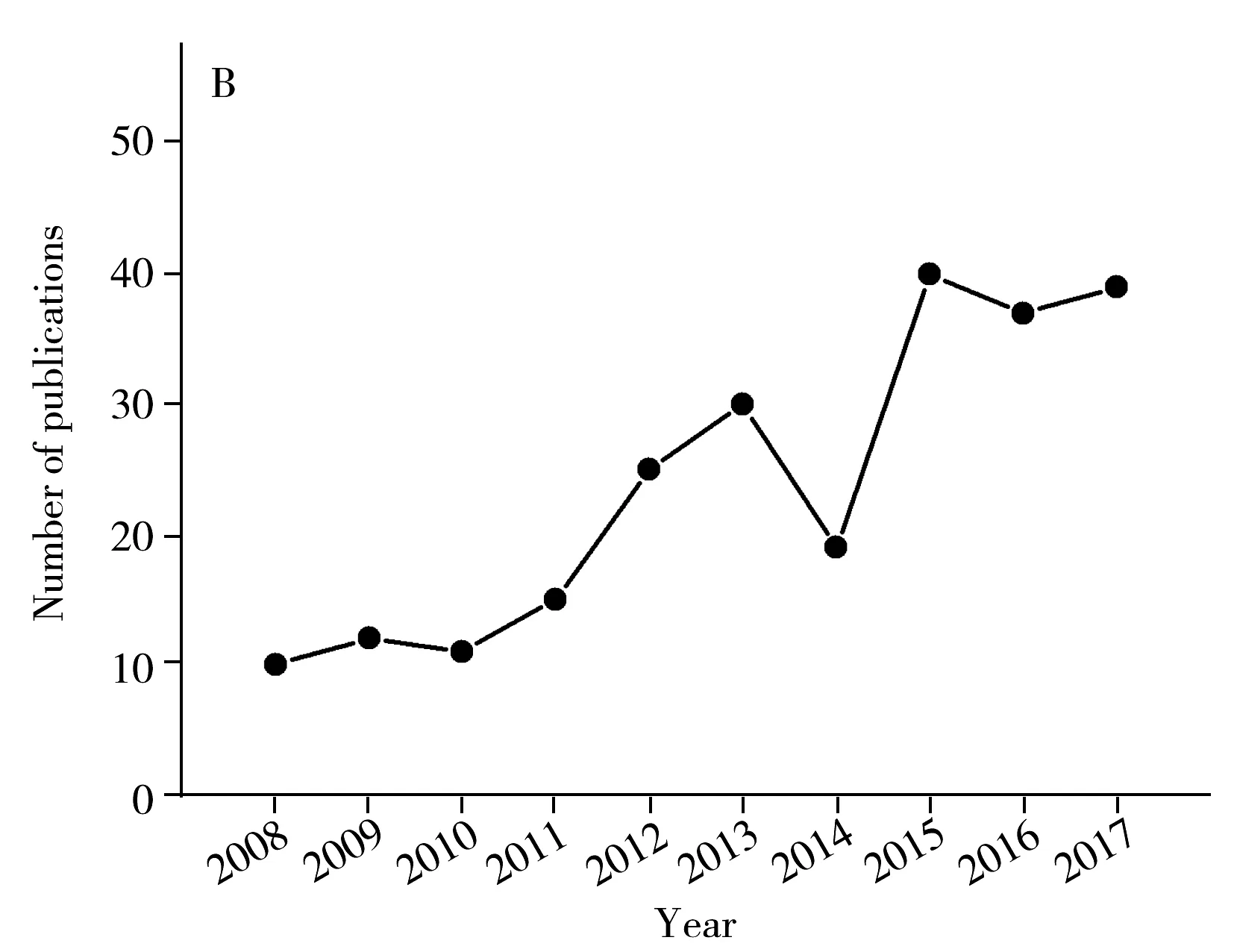
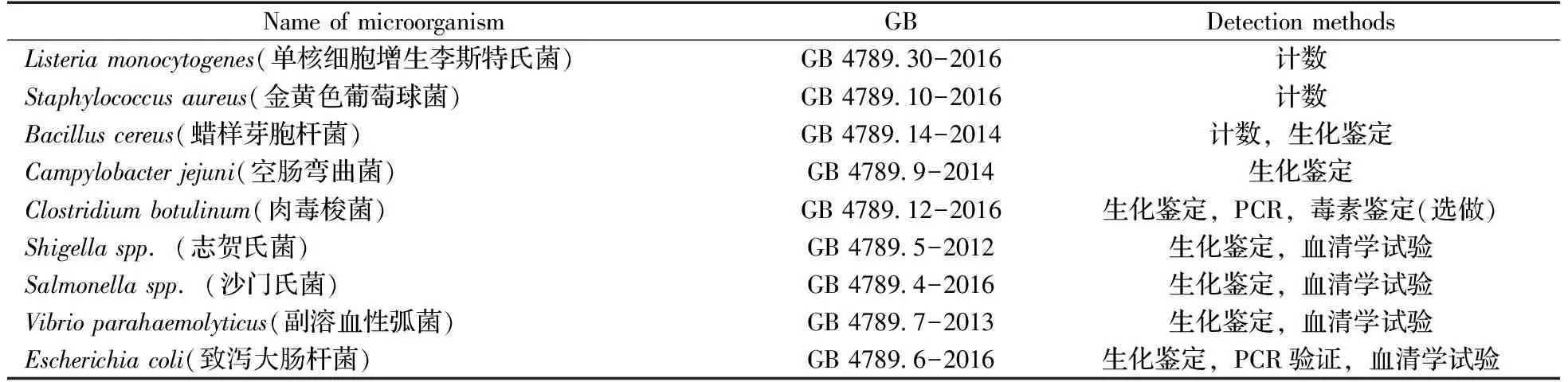
圖2 沙門氏菌PCR檢測方法研究情況Fig.2 Publications of Salmonella spp.detection methods targeting specific genes(data were derived from Web of Science)A.PCR based methods for Salmonella spp.(沙門氏菌核酸擴增方法研究數量);B.isothermal amplification based methods for Salmonella spp.(沙門氏菌等溫擴增方法研究數量)
2 等溫擴增技術
經典的PCR方法能夠在幾小時內完成對目標核酸的指數級擴增,具有檢測時間短、適用范圍廣的優勢,但也存在明顯的缺陷。PCR過程需要經歷變性、退火、延伸3個步驟的多次循環,而且每一個步驟都需要在特定的溫度下進行,通常分別為94、55、72 ℃。變溫循環要求PCR儀器配備快速變溫單元,導致儀器不便移動和攜帶,而實際應用中又存在著大量現場或在線檢測的需求,由此催生了多種可在恒溫條件下進行的PCR技術,稱為等溫擴增技術,如環介導等溫擴增(Loop-mediated isothermal amplification,LAMP)、滾環擴增(Rolling circle amplification,RCA)、鏈置換擴增(Strand displacement amplification,SDA)、切口酶信號擴增(Nicking enzyme signal amplification,NESA)和核酸外切酶Ⅲ輔助擴增(Exonuclease Ⅲ assisted amplification)技術等[4-6],詳見表2。這些技術不需要復雜的溫度變化,在一個恒定溫度下就可以實現對檢測目標或者信號的擴增,極大地降低了對儀器的需求,具有現場應用或在線檢測的潛力。

表2 常見5種等溫擴增方法比較Table 2 Comparison of 5 isothermal amplification methods
2.1 環介導等溫擴增技術(LAMP)
LAMP技術是目前最為常用的病原菌現場檢測技術,在2000年首次提出[7]。與傳統PCR不同,LAMP技術針對模板6個不同區域的位置,設計可以自發形成環狀結構的引物,在不斷復制中將線性的目標DNA序列無限拓展為之字形的長鏈結構。盡管LAMP的反應過程非常復雜,但是結果的判斷卻簡便、可視。這是由于反應產生的大量焦磷酸離子與鎂離子形成了難溶于水的焦磷酸鎂沉淀,使得肉眼即可定性判別檢測結果。但更精確的定量判斷則需要借助于濁度計或使用能識別DNA雙鏈的核酸染料[8]。目前,研究人員已經成功地將LAMP技術應用于多種致病微生物的檢測中,包括單增李斯特菌(Listeriamonocytogenes)[9-11]、沙門氏菌(Salmonellaspp.)[12-14]、空腸彎曲桿菌(Campylobacterjejuni)[15-17]、產志賀毒素大腸桿菌(EscherichiacoliO157)[18]、副溶血性弧菌(Vibrioparahaemolyticus)[19-21]和霍亂弧菌(Vibriocholerae)[22-24]等。LAMP方法擴增速度快、靈敏度高、無需變溫,在所有等溫擴增方法中應用最廣。但是,LAMP的擴增產物組成復雜,多種長度片段共存的產物起不到再次驗證擴增特異性的作用;此外,60 ℃的反應溫度仍然需要裝置維持;極快的擴增速率也增加了假陽性的風險。這些缺點是LAMP技術所亟待解決的。

圖3 滾環擴增技術(RCA)原理示意圖Fig.3 Principle of rolling circle amplification
2.2 滾環擴增技術(RCA)
RCA的原理與滾環復制有很多相似之處,滾環復制是噬菌體和病毒常見的環狀雙鏈DNA復制方式,由具有3端至5端外切酶活性的DNA聚合酶Ⅲ在切口處同步進行新鏈合成和原有鏈去除[25]。RCA技術只使用單側引物,結合在環形待檢測的核酸序列上,在聚合酶的作用下,不斷循環復制,形成不斷重復并且與環形模板互補的DNA長鏈[26];待測核酸非環狀類型時,可將鎖狀引物(Padlock probe)與待測線型核酸結合,使用DNA連接酶連成環形,以鎖狀引物的序列代替核酸序列進行擴增,進而使用相應策略檢測引物的數量(圖3)。Zhou等[27]巧妙地將一小段雙鏈結構加入到鎖狀引物中,形成啞鈴型。由于雙鏈結構的存在,RCA擴增的長鏈DNA中也會形成多個雙鏈結構,在反應體系中加入嵌入型熒光染料,即可對雙鏈結構的數目做出估計,達到對目標核酸序列定量分析的目的。Guo等[28]將RCA與氯高鐵血紅素的氧化還原反應結合,用于大腸桿菌(Escherichiacoli)的定量測定。Teng等[19]先用抗體與適配體混合的夾板式方法捕獲目標菌體,再通過RCA技術放大信號,實現了對魚肉中副溶血弧菌的精準測定。由于鎖狀引物的序列是人為設計的,因此RCA的產物序列具有很強的拓展性,可以根據實際需要來調整,以便形成具有信號產生功能或者催化活性的DNA高級結構,也易于實現指數級擴增。此外,RCA的產物在空間上是連續的,因此是唯一可進行原位擴增的等溫擴增策略,特別適合在芯片表面進行非均相體系擴增。但是,線性單鏈DNA需與鎖狀引物進行連接反應后再開始擴增反應,這決定了RCA容易受到復雜溶液體系的干擾,所以RCA在均相體系中的檢測效果不如非均相體系[28]。

圖4 鏈置換擴增技術(SDA)原理示意圖Fig.4 Principle of strand displacement amplification
2.3 鏈置換擴增技術(SDA)
SDA是一種通過建立循環置換機制實現擴增特定核酸序列的方法,與LAMP相比,SDA對反應溫度要求低,在室溫條件下也能進行(圖4)。SDA通常產生較短的DNA單鏈,與嵌入型熒光染料、Tagman探針或分子信標(Molecular beacon,MB)等結合,在擴增目標序列的同時放大檢測信號,為定量檢測提供基礎。SDA單鏈的產生可以由具有3端至5端外切酶活性的DNA聚合酶引發[29],也可以由基于熱力學穩定性的Toehold置換引發[30-31]。由于產物為單鏈DNA,具備識別其他核酸序列的能力,因此SDA也是實現DNA邏輯運算的常用手段[32]。通常情況下,SDA中的靶標在啟動一次置換反應后,無法被回收,因而只能實現線性擴增,但在切口酶(Nicking enzyme)幫助下,也可實現指數級的快速擴增[33]。Tan等[34]將兩段與目標序列互補的DNA通過切口酶的結合序列串聯起來,以堿基配對原則與目標序列結合,在聚合酶作用下形成平末端DNA雙鏈后,切口酶結合到新形成的雙鏈識別原件上,此時可剪開一個缺口,釋放剛合成的單鏈(目標序列),用于下一輪循環;與此同時,被切口酶切割的雙鏈結構可在聚合酶作用下重新形成目標序列,達到指數級擴增效果。然而,指數級SDA需要多個DNA結構單元配合,體系較為復雜,容易受到干擾;SDA反應的觸發需要DNA單鏈,無法直接對雙鏈DNA擴增。這些缺點限制了SDA的大范圍應用,目前尚無基于SDA的商業化檢測產品。
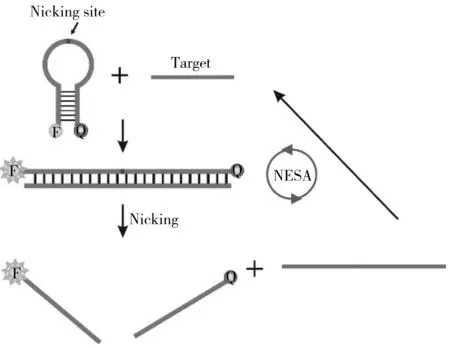
圖5 切口酶擴增技術(NESA)原理示意圖Fig.5 Principle of nicking enzyme signal amplification

圖6 核酸外切酶擴增技術原理示意圖Fig.6 Principle of exonuclease Ⅲ assisted amplification
2.4 切口酶信號擴增技術(NESA)
切口酶(Nicking enzyme)與限制性內切酶相似,能夠識別特異的核酸序列,但切口酶只催化一條鏈的磷酸二酯鍵斷裂。切口酶的這一特性主要用于均相檢測中核酸靶標的回收以及信號的產生。切口酶介導的擴增體系由靶標識別元件、信號產生元件、切口酶識別序列組成,通過切口酶切割信號探針并回收靶標用于下一輪信號產生(圖5)。Li等[35]設計了一種特殊的分子信標,信標的環形部分堿基序列與需要檢測的目標序列互補,并且其中含有切口酶的識別序列。信標與目標序列互補后產生的雙鏈結構,在熱力學穩定性上要高于分子信標的莖環結構。因此,當目標序列出現在檢測體系中時,會使分子信標的構象發生改變,打開原來的莖環結構,形成新的雙鏈DNA結構,進而被切口酶切割。切割后,雙鏈之間的氫鍵數目顯著減少,穩定性大大降低,在55 ℃的條件下會自動解離,釋放目標序列,用于結合下一個分子信標,形成擴增循環。目前已經有采用類似策略檢測腸炎沙門氏菌(Salmonellaenteritidis)16S rRNA序列的報道[36]。切口酶等溫擴增方法的最大困難在于酶切識別位點的引入。通常情況下,核酸靶標中不含有識別序列,需要結合其他策略來生成,這限制了NESA的應用[37]。NESA更適合作為SDA的輔助單元,配合進行信號的二次放大[38]。
2.5 核酸外切酶Ⅲ輔助擴增技術
除了切口酶,核酸外切酶Ⅲ(Exonuclease Ⅲ)也能通過類似的策略,達到持續放大信號的目的[39]。核酸外切酶Ⅲ具有3端到5端脫氧核糖核酸外切酶活性,與切口酶相比,核酸外切酶不需要特定的識別序列即可工作,并且只作用于DNA雙鏈的平末端或者3端方向凹陷的粘性末端,因此適用面更廣(圖6)。Zuo等[40]對常規的分子信標結構做了些許調整,在3端猝滅基團后增加了若干堿基,形成凸出型的3端粘性末端,以避免信標本身被核酸外切酶Ⅲ降解。當信標結合目標DNA序列形成雙鏈結構時,3端變為平末端類型,堿基逐個被核酸外切酶剝離,熒光基團也因為猝滅基團的遠離而發光。當剩余的堿基數量不足以維持穩定的雙鏈結構時,5端含有熒光基團的短鏈解離,釋放目標序列進行下一個循環。Luo等[41]將捕獲探針固定在電極表面,利用核酸外切酶Ⅲ持續地降解與目標結合的捕獲探針,使得電極表面的DNA數量持續減少,導電性增加,據此來檢測常見的腸道細菌特征DNA序列。核酸外切酶Ⅲ只識別粘性末端類型而不區分DNA序列的特點,便于在非均相體系中同時降解不同序列的DNA雙鏈,所以主要用于固相材料捕獲靶標DNA后信號的產生。特別當與石墨烯等納米材料結合時,核酸外切酶Ⅲ可以快速釋放熒光基團,從而解除石墨烯對熒光的猝滅效應,產生信號[42]。但是,經核酸外切酶Ⅲ降解后的單鏈DNA分解為單個游離核苷酸,無法進行回收,因此難以實現循環,一般只能實現信號的線性擴增。
3 等溫擴增存在的共性問題
等溫擴增技術在近十多年間的發展非常迅速,LAMP、RCA、NESA、SDA等核酸高效擴增方法的出現,有效地縮短了檢測所需時間,也降低了對復雜檢測設備的依賴,為致病微生物的現場實時檢測提供了良好的基礎。目前,等溫擴增技術已經成為致病微生物快速檢測的重要方法之一,常見的食源性致病菌,均有眾多成熟的等溫擴增檢測方法報道,以及基于等溫擴增原理開發的商業化試劑盒[43-46]。但隨著人們對食品安全問題關注程度的逐漸提高,以及政府部門監管力度的逐漸加強,致病菌檢測的需求急劇增長,并且出現了不少新變化和新問題。對此,等溫擴增方法還需要在一些共性問題上加以改進,以適應這些實際需求。
3.1 活性識別
等溫擴增所檢測的物質是致病菌DNA,其性質相對穩定,細菌死亡后DNA不會立即降解,因此仍可以被擴增手段檢測到。嚴格意義上來說,等溫擴增所得到的結果,只能證明樣品中細菌DNA的存在,而無法獲知細胞的生存狀態,這可能導致對細菌危害風險的誤判。
3.2 非特異性擴增
快速是等溫擴增的關鍵特點,也是實現其高靈敏度的重要原因。然而,短時間內對目標基因的高速率復制,在顯著提升方法靈敏度、縮短檢測時間的同時,也大大增加了等溫擴增對于非靶標DNA的敏感性,導致非特異性擴增概率上升,產生假陽性。例如,在LAMP操作過程中,只要反應體系與外界空氣有過短暫接觸,就非常容易受到氣溶膠污染,導致擴增產物中含有大量的非目標基因片段。
3.3 前處理步驟
復雜的溶液成分會影響氫鍵的穩定性,從而降低引物對靶基因的識別效率。因此,樣品的前處理對于等溫擴增技術來說非常關鍵。細菌DNA必須經過富集、提取和純化,減弱其他因素的干擾,才能被高效識別和擴增,否則將會引起背景值偏高或信號值減弱,導致信噪比降低。由于前處理主要依賴人工操作,在某些情況下,前處理所消耗的時間已經超過了等溫擴增所需時間,成為等溫擴增進一步快速化的制約因素[47]。
3.4 檢測通量
為了進一步提高等溫擴增的效率,更好地適應現場或在線檢測,不少研究都致力于在單個等溫擴增反應中實現多個核酸靶標的檢測,即多重等溫擴增策略[48](圖7)。但是,單純通過增加多組引物達到同時擴增多個目的片段的方式具有較大局限性。增加引物組數必然導致體系的復雜化,不同組引物之間存在交叉組合的可能,增加了獲得非特異性條帶的幾率。目前,多重等溫擴增策略能同時檢測的靶標一般為2到3種,3種以上的多重反應往往由于引物組之間的干擾導致靈敏度不佳。
3.5 解決措施
在細菌活性區分問題上,研究發現,提取DNA前用疊氮溴化丙錠(Propidium monoazide,PMA)處理細菌,可以實現對活菌的選擇性擴增。死亡細菌通常會喪失對細胞壁通透性的調控能力,使得PMA可以自由通過,并結合死亡細菌核區內的基因組DNA,阻止其被DNA聚合酶結合[13]。基于此原理,馬骉等[49]構建了PMA-LAMP方法,成功區分了樣品中死亡的和具有活性的副溶血弧菌。但是,在某些情況下,細胞壁的通透性并不能實時反映細菌的活性,此時PMA并不能很好地發揮區分活性的作用[50]。
在非特異性擴增問題上,張明洲等[51]優化了鈣黃綠素、氯化錳配比及反應液鎂離子濃度,降低了染料對于酶擴增效率的影響,從而省略了擴增完成后加入染料的步驟,使得污染幾率顯著降低。也有研究者從LAMP產物的識別角度出發,利用分子信標的特異性識別功能,只對目標序列產生信號[52],雖然在一定程度上降低了假陽性率,但是污染引發的非特異性擴增依然存在,并且會對目標基因的擴增效率產生影響。
在前處理問題上,高通量的前處理裝置已經逐步用于食品樣品的快速檢測,例如基于納米級中空纖維膜的農獸藥高通量萃取裝置[53]。細菌核酸提取的高通量、自動化前處理方法和處理裝置的研究也有不少報道,這有助于進一步縮短等溫擴增的檢測時間,減少人工操作步驟,是等溫擴增規模化檢測的重要基礎[54]。
在檢測通量問題上,伴隨著等溫擴增芯片化的研究趨勢(圖7),等溫擴增的體系得以不斷縮小,單位面積可容納的獨立反應個數得到增加,這為解決檢測通量問題提供了新的角度。Zhou等[55]設計了基于離心作用的LAMP微流控芯片,通過在圓盤形平面上構建獨立反應通道來增加通量,可以同時檢測10種常見的食源性致病菌。
4 結 論
綜上所述,雖然等溫擴增已有將近20年的歷史,但因其準確性高、檢測用時短、便攜易操作的特點。目前依然作為快速檢測和在線檢測的主要手段被廣泛應用于食源性致病微生物的檢驗檢疫中。以LAMP和NEAR為代表的商業化現場檢測產品的成功,也證明了等溫擴增在微生物現場檢測中的價值。等溫擴增的相關研究持續受到較高的關注,其技術缺陷也正在被逐步克服,芯片化、高通量的等溫擴增方法及其裝置正在逐漸成為熱點,隨著技術體系的不斷完善,其應用前景將會更加廣闊。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
