棘孢木霉JM -1菌株對麥根腐離蠕孢的拮抗機制
2019-08-03 07:16:06張眉吳斌徐德坤王升吉辛相啟姜珊珊
山東農業科學 2019年3期
張眉 吳斌 徐德坤 王升吉 辛相啟 姜珊珊
摘要:為了研究木霉菌JM-l菌株對麥根腐離蠕孢CY6菌株的拮抗機制,本研究結合形態學和分子生物學鑒定方法,明確JM-l菌株的分類地位,并采用平板對峙培養法和顯微觀察,分析JM-l菌株對CY6菌株的拮抗作用。結果表明,木霉菌JM-l菌株為棘孢木霉(Trichoderma a.sperellurn),可通過空間和營養競爭抑制CY6菌株的生長,抑菌率為77.22%;顯微觀察發現,棘孢木霉JM-l對CY6菌株有明顯的重寄生作用,同時,JM-l菌株可產生某些抗生物質,使CY6菌株的菌絲、分生孢子發生變形和消解。因此,棘孢木霉JM-l菌株可通過競爭作用、重寄生作用和抗生作用等多重生防機制抑制麥根腐離蠕孢的生長。
關鍵詞:棘孢木霉;麥根腐離蠕孢;拮抗機制
Antagonistic Mechanism of Trichodermaasperellum JM - 1 against Bipolaris sorokinianaZhang Mei, Wu Bin, Xu Dekun, Wang Shengji, Xin Xiangqi, Jiang Shanshan
Abstract In order to study the antagonistic mechanism of Trichoderma spp. JM - I against Bipolaris so-rokiniana, the taxonomic status of JM - 1 was confirmed by morphological and molecular biological identifica-tion methods, and the antagonistic mechanism between JM -1 and CY6 isolates were analyzed by plate con-frontation culture and microscopic observation. The results showed that JM - 1 isolate was identified as Tri-choderma asperellum, and it restrained the growth of CY6 isolate through spatial and nutrient competition withand the inhibition rate as 77.22% . Microscopic observation found that JM - 1 isolate had the significant para-sitic effect on CY6 isolate, and produced some antibiotic substance, which led to deformation and digestion ofmycelia and conidia. In summary, T. asperelluim JM - 1 isolate could inhibit the growth of B. sorokinianathrough multiple biological control mechanisms such as competition, mycoparasitism and antibiosis.
Keywords Triclzoderma asperellum ; Bipolaris sorokiniana ; Antagonistic mechanism
木霉菌(Trichoderma spp.)對多種植物病原菌都具有較好的拮抗作用[1],是一類應用廣泛、適應力強、拮抗機制多樣的生防菌[2],在防治植物病害上發揮著重要作用。棘孢木霉(Trichoder-ma asperellum]是由Samuels等[3]于1999年命名的木霉新種,在我國由章初龍等[4]于2005年首次報道。目前研究表明,棘孢木霉是一種優良的生防真菌,對黃瓜枯萎病菌(Fusarium oxyspo-rum)[5]、玉米大斑病菌(Exserohilum turcicum)[6]、水稻紋枯病菌(Rhizoctonia solani)[7]等病原菌都有良好的抑制效果。
小麥根腐病是山東小麥產區的重要病害之一,該病危害嚴重并呈逐年加重趨勢,主要病原物為麥根腐離蠕孢(Bipolaris sorokiniana)。崔云龍等[8]研究表明,短小芽孢桿菌D82對小麥根腐病有較強的抗生作用,防效顯著。王立等[9]研究發現,叢枝菌根真菌摩西球囊霉( Glomus mosseae)能有效防治小麥根腐病,并對小麥的營養生長有促進作用。目前利用木霉菌防治小麥根腐病的研究報道較少。本研究采用形態學和分子生物學鑒定方法,確定木霉菌JM-1菌株的分類地位,通過平板對峙培養和顯微觀察,明確其對麥根腐離蠕孢的拮抗機制,為木霉菌JM-1菌株防治小麥根腐病提供理論依據。
1 材料與方法
1.1供試菌株
供試菌株:木霉菌JM -1菌株分離自小麥根際土壤;小麥根腐病麥根腐離蠕孢(Bipolaris soro-kiniana)CY6菌株為本實驗室保存。
1.2 生化試劑及其他
Easy Taq酶購自北京全式金生物技術有限公司,真菌DNA提取試劑盒購自Omega公司,DNA凝膠回收試劑盒購自上海Sangon公司。其他常規化學藥品均為分析純試劑。PCR引物合成由上海Sangon公司完成。DNA測序由鉑尚生物技術(上海)有限公司完成。
1.3 木霉菌的鑒定
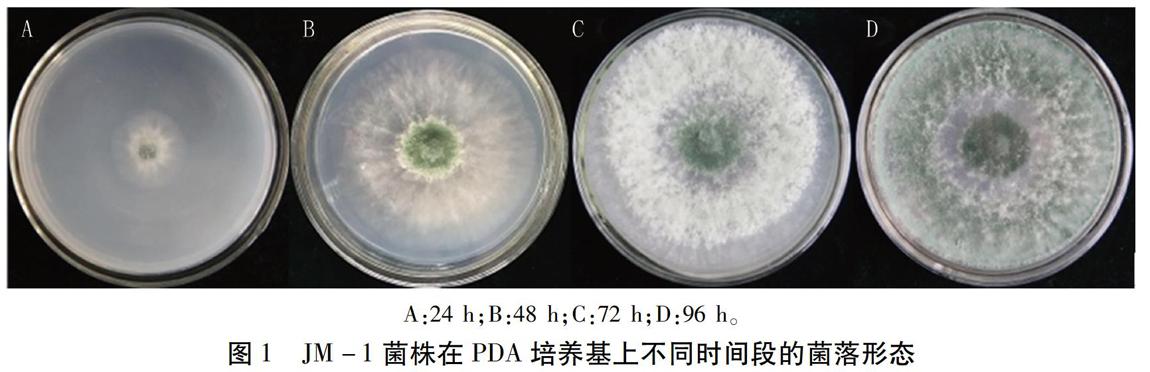
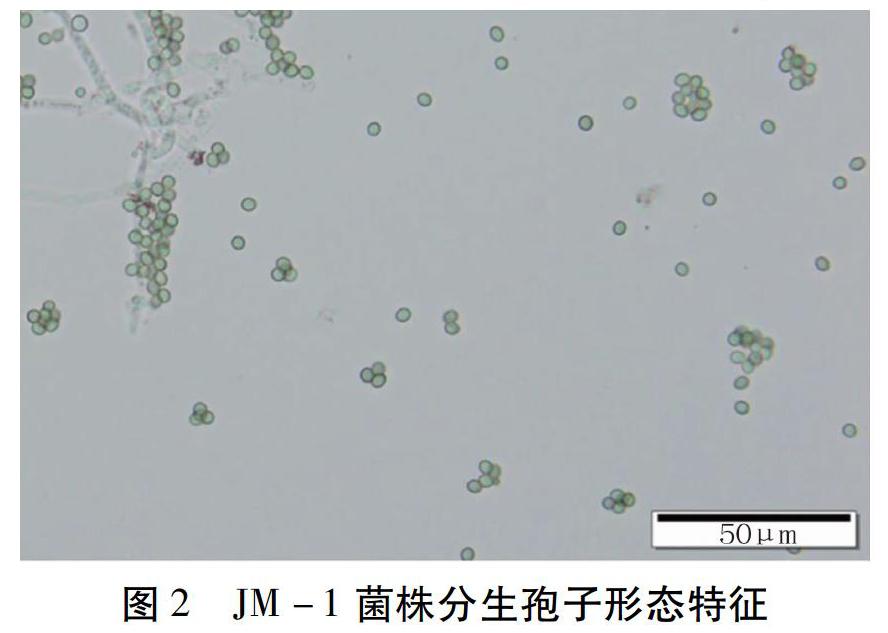
1.3.1 形態學鑒定 將純化的木霉菌JM -1菌株接種于PDA培養基平板,28C倒置培養,分別在接種24、48、72、96 h觀察菌落形態、顏色及培養性狀。挑取菌絲,顯微觀察分生孢子梗和分生孢子的形態特征。
1.3.2 分子生物學鑒定 收集JM -1菌株的菌絲體,用HP Fungal DNA Kit提取試劑盒提取菌株基因組,以DNA為模板,用通用引物ITSIF(5-TCCGTAGGTGAACCTGCGG -3)和ITS4R(5 -TCCTCCGCTTATTGATATGC -3)擴增rDNA -ITS序列,水作陰性對照。PCR反應體系為25μL:IO×Easy Taq Buffer(含Mg2+)2.5μL,2.5mmol/L dNTPs 2poL,5 U/pL Easy Taq 0.3μL,10ymol/L正反向引物各0.5μL,DNA模板1μL,ddH,0補足至25μL。PCR反應程序為:94℃ 4min;94℃ 30 s,56℃ 30 s,72℃ 35 s,35個循環;72℃ 10 min。切膠回收目的條帶進行測序。
在GenBank數據庫中下載合適的木霉菌株序列,通過Clustalx l.83軟件對ITS序列進行比對。利用MEGA 5.0分析軟件中的Neighbor-Joining分析法構建系統發育樹,基于Kimura 2-parameter距離模式計算遺傳距離。各分支置信度(Bootstrap)進行1000次重復分析。
1.4 木霉菌對病原菌的拮抗作用
采用平板對峙培養法:木霉菌JM-1菌株和麥根腐離蠕孢CY6菌株均置于28℃活化3d,然后用直徑7mm的打孔器分別在菌落邊緣打取菌餅,接種于PDA平板中線上相距7 cm的兩點處,28℃黑暗培養。以一端接種病原菌菌餅,一端接種空白PDA菌餅作對照,每個處理重復3次。每隔24h測量菌落半徑,計算抑菌率。抑菌率(%)=(對照病原菌菌落半徑一處理病原菌菌落半徑)/對照病原菌菌落半徑×l00。
1.5 拮抗作用的顯微觀察
在PDA平板兩端分別接種木霉菌JM-1菌株和麥根腐離蠕孢CY6菌株的菌餅,在菌餅之間平放無菌蓋玻片,28℃黑暗培養。待兩端菌絲長至蓋玻片上,菌絲相交后,進行顯微觀察。以蓋玻片上只長有病原菌作對照。
2 結果與分析
2.1 木霉菌JM-1菌株的形態學鑒定
在PDA培養基上培養24 h菌落圓形,呈輻射狀生長,菌絲白色;培養48 h菌落中央產生綠色孢子;72 h菌落即能長滿直徑9 cm的培養皿,并出現一圈稀薄一圈厚密的菌絲生長帶,綠色孢子由中央向四周擴展;96h厚密的菌絲生長帶上產生大量綠色孢子,菌落呈現白綠相間的同心圓狀(圖1)。
顯微鏡觀察發現,菌絲細長,有隔,分生孢子梗對生分支,頂端有瓶梗,瓶梗2個或多個為一組輪生,瓶梗中間膨大,呈安瓿瓶形。分生孢子大小為(2.95~4.07) μm×(3.11~4.69)卜Lm,青綠色,圓形或卵形(圖2)。該菌株的培養性狀和形態特征與棘孢木霉(Trichoderma asperellum)基本一致,因此初步鑒定為棘孢木霉。
2.2 木霉菌JM-1菌株的分子生物學鑒定
用通用引物ITSIF/ITS4R擴增JM-1菌株基因組的ITS序列,得到一條約600 bp的目的條帶(圖3)。將測定的JM-1菌株ITS序列與GeneBank數據庫中下載的木霉序列進行同源性比較并構建系統發育樹,從樹狀圖(圖4)可以看出,JM-1菌株聚類到Trichoderma asperellum進化枝上。綜合形態學和分子生物學鑒定結果,明確JM-1菌株為棘孢木霉(Trichoderma asperel-lum)。
2.3 棘孢木霉對麥根腐離蠕孢的拮抗作用
在PDA平板對峙培養過程中,棘孢木霉JM-1菌株對麥根腐離蠕孢CY6菌株有較強的抑制作用,抑菌率為77.22%。相比CY6菌株,JM-1菌株生長迅速,培養3d菌絲即與CY6菌絲相交,CY6菌絲的生長明顯受到抑制,菌落邊緣出現輕微塌陷。培養至第4d,JM-1菌絲將CY6菌落包圍,菌絲幾乎不能生長,而JM-1菌絲覆蓋在CY6菌落上繼續生長。對峙培養10d,JM -1菌絲已將CY6菌落完全覆蓋,并在CY6菌落上產生大量綠色孢子(圖5)。從以上結果可以看出,棘孢木霉JM -1菌株能夠通過對生長空間和營養物質的競爭有效抑制麥根腐離蠕孢CY6菌株的生長。
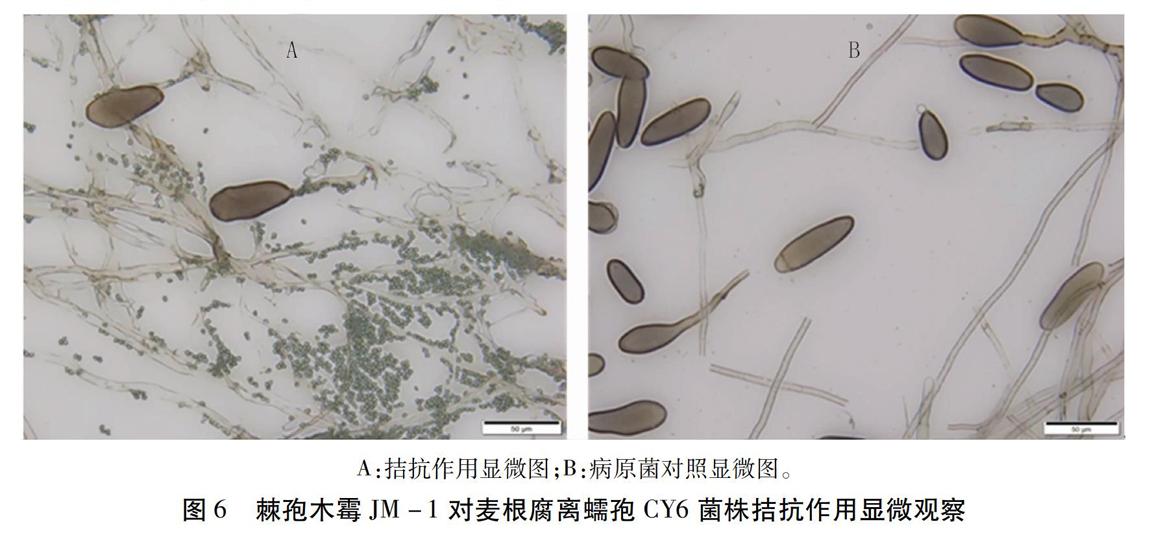
2.4 棘孢木霉對麥根腐離蠕孢拮抗作用的顯微觀察
在PDA培養基上,棘孢木霉JM -1菌株與麥根腐離蠕孢CY6菌株對峙培養3d菌絲相交,顯微觀察發現,棘孢木霉JM-1菌絲逐漸纏繞CY6菌絲,與對照相比,CY6菌絲發生變形、斷裂、消解,分生孢子量明顯減少,并出現變形、破裂現象(圖6)。由此可見,棘孢木霉JM-1菌株可通過纏繞的方式寄生在CY6菌絲上,具有重寄生作用,此外,JM-1菌株還可能產生了某些抗生物質,導致CY6菌絲和分生孢子變形、解體。
3 討論與結論
木霉屬真菌廣泛存在于土壤、植物殘體等生態環境中,是土壤中微生物種群的優勢真菌[10]。該屬真菌種類繁多,常見的木霉屬真菌有哈茨木霉(T.harzianum)、綠色木霉(T.viride)、棘孢木霉(T.asperelluim)、深綠木霉(T.atroviride)、長枝木霉(T. longibrachiatum)、康寧木霉(T.konin-gii)等。本研究從菌落形態、分生孢子梗及分生孢子特性等方面,初步確定木霉菌JM-1菌株為棘孢木霉。但由于木霉菌形態特征較復雜,同時缺乏穩定性[5],單從形態特征上不能準確確定分類地位。真菌rDNA - ITS序列因既具有保守性,又具有序列多態性,能夠有效鑒定微生物的種屬關系。本研究通過擴增JM-1菌株ITS序列并構建系統發育樹發現,木霉菌JM-1菌株聚類在了Trichoderma asperellu:m進化分支上。分子生物學鑒定結果與形態學鑒定結果一致,明確了木霉菌JM-1菌株為棘孢木霉(Trichoderma asperellum)。
木霉菌可通過競爭作用、重寄生作用、抗生作用、誘導抗性和協同拮抗作用等多重生防機制防治植物病害[ll]。李琳等[6]篩選獲得一株棘孢木霉T31菌株,該菌對玉米大斑病菌有較強的生長競爭優勢,同時具有明顯的重寄生作用。夏偉等[12]研究結果表明,棘孢木霉菌株I4能夠通過競爭和抗生雙重作用有效抑制立枯絲核菌的生長。本研究發現,棘孢木霉JM-1菌株能夠快速占領生長空間,且能覆蓋在麥根腐離蠕孢CY6菌落上,表明棘孢木霉JM -1菌株具有較強的空間和營養競爭優勢,可通過競爭作用抑制CY6菌絲的生長。同時,棘孢木霉JM-1菌株還可纏繞在CY6菌株的菌絲上,通過重寄生作用吸取CY6菌絲的營養物質,使其菌絲發生斷裂、解體。木霉菌發揮抗生作用而產生的抗菌物質主要有抗菌肽類、抗生素類以及揮發性物質[13],這些代謝產物能夠引起病原菌菌絲原生質凝聚、菌絲變形解體[12,14],抑制病原孢子萌發[15]等。本研究顯微觀察發現,棘孢木霉JM-1菌株與麥根腐離蠕孢CY6菌株相交后,CY6菌絲發生變形、斷裂、解體,分生孢子發生變形消解。由此可以推斷,棘孢木霉JM -1菌株可能產生了某些抗菌物質,從而對麥根腐離蠕孢CY6菌株起到抗生作用。綜上,棘孢木霉JM-1菌株可通過競爭作用、重寄生作用和抗生作用有效抑制麥根腐離蠕孢的生長。下一步將針對棘孢木霉JM-1菌株產生的抗生物質主要成分以及該菌株對小麥根腐病的生防作用做進一步研究。
參考文獻:
[1]郭潤芳,劉曉光,高克祥,等.拮抗木霉菌在生物防治中的應用與研究進展[J].中國生物防治,2002,18(4):180 -184.
[2] 吉海龍,伊洪偉,池玉杰.長枝木霉菌株T05抑菌活性與拮抗機制[J].東北林業大學學報,2016,44(1):114 -119.
[3] Samuels G J,Lieckfeldt E,NirenbergH I.Trichoderma a.s-perellum,a new species with warted conidia. and redesCriptionof Zviricle[J].Svdowia,1999,51(1):71 - 88.
[4] 章初龍,徐同.我國河北、浙江、云南及西藏木霉種記述[J].菌物學報,2005,24(2):184 -192.
[5] 夏偉,顏艷偉,張紅,等.木霉菌Trl0的鑒定及其對幾種病原菌的抑制作用[J].安徽農業科學,2010,38(5):2424 - 2426.
[6] 李琳,張雅梅,張祥輝,等.生防棘孢木霉T31菌株的分離篩選及其生物學特性[J].植物保護學報,2014,41(1):54 -60.
[7] 陳立華,金秋,牛明,等.棘孢木霉對水稻紋枯病病原菌立枯絲核菌生物防治的研究[J].江蘇農業科學,2015,43(5):115 -117.
[8] 崔云龍,姬金紅,衣海青.短小芽孢桿菌D82對小麥根腐病原菌拮抗的研究[J].中國生物防治,1995,11(3):114 - 118.
[9] 王立,王敏,馬放,等.叢枝菌真菌對小麥的促生長效應與根腐病抑制效應[J].哈爾濱工業大學學報,2015,47(4):15 -19.
[10]張廣志,楊合同,文成敬.木霉菌形態學分類檢索與分子生物學鑒定[J].山東農業大學學報(自然科學版),2011,42(2):309 -316.
[11] 王永陽,杜佳,高克祥.苦瓜枯萎病生防木霉的篩選鑒定及其定殖的qPCR檢測[J].山東農業科學,2018,50(8):110 - 115.
[12] 夏偉,張紅,顏艷偉,等.棘孢木霉L4對立枯絲核菌的拮抗機制[J].植物保護學報,2010,37(5):477 -478.
[13] 李沖偉,楊立賓,鄧勛,等.木霉菌株對金黃殼囊孢菌的抑菌效應及機制[J].林業科學,2012,48(9):88 -94.
[14] 朱天輝,邱德勛.Trichoderma harzzanum對Rhizoctonic solani的抗生現象[J].四川農業大學學報,1994,12(1):11 -15.
[15] 蔣妮,白丹宇,宋利沙,等.棘孢木霉F2菌株對三七灰霉病的生物防治作用[J].江蘇農業科學,2018,46(20):94 - 97.
