2016-2017 年廣東地區(qū)豬繁殖與呼吸綜合征病毒的分子檢測及NSP2、ORF5 基因變異分析
2019-08-20 07:06:06周吉培申翰欽劉相聰于林洋董建國張樂宜劉燕玲梁鵬帥宋長緒
中國獸醫(yī)雜志 2019年4期
徐 錚,周吉培,申翰欽,劉相聰,于林洋,董建國,2,張樂宜,劉燕玲,王 磊,梁鵬帥,宋長緒
(1.華南農(nóng)業(yè)大學動物科學學院 國家生豬種業(yè)工程技術研究中心,廣東 廣州510642;2.信陽農(nóng)林學院牧醫(yī)工程學院,河南 信陽464000)
豬繁殖與呼吸綜合征(PRRS)系由豬繁殖與呼吸綜合征病毒(PRRSV)引起的母豬繁殖障礙和各階段豬呼吸道癥狀的高度接觸性傳染性疾病,現(xiàn)已成為嚴重威脅全球養(yǎng)豬業(yè)的主要疫病之一。該病毒基因組為單股正鏈RNA,長約15 kb,包含10 個開放閱讀框(ORF),其中ORF5 和NSP2 基因變異最大,是作為分析PRRSV 遺傳變異的理想靶基因[1]。該病毒可分為2 個基因型:以Lelystad-virus(LV)株為代表株的歐洲型和以VR-2332 株為代表株的美洲型病毒株[2],分別是Wensvort 等[3]在1991 年和Benfield 等[4]于1992 年分離得到。郭寶清等[5]1996 年首次在國內分離到一株病毒,并命名為CH-1a,成為我國經(jīng)典毒株的代表。2001 年在美國分離到具有NSP2 蛋白的(111+1+19)共131 個氨基酸不連續(xù)缺失為特征的NADC30 株[6]。2006 年我國分離到PRRSV 的變異株—高致病性豬繁殖與呼吸綜合征病毒(HP-PRRSV)[7],其代表性毒株為JXA1和HUN4,此類毒株的NSP2 蛋白存在(1+29)共30個不連續(xù)氨基酸缺失的特征[8]。
本研究對2016-2017 年采自廣東省各地疑似PRRSV 感染病料進行檢測,并對其中的9 份陽性樣品的NSP2 和ORF5 基因進行測序和比對分析,以揭示廣東地區(qū)豬群PRRSV 最新流行動態(tài)和遺傳變異特點,希望為該地區(qū)PRRSV 的科學防控提供一定的有效參考依據(jù)。
1 材料與方法
1.1 病料樣品 采集和送檢PRRSV 的906 份病豬淋巴結、脾臟、肺臟等組織病料,樣品來自廣東省各地市的疑似發(fā)病豬場。
1.2 菌株、細胞及載體 大腸桿菌DH5α 感受態(tài)細胞、Marc-145 細胞均由華南農(nóng)業(yè)大學國家生豬種業(yè)工程技術研究中心保存;pMD-19T 克隆載體,購自寶生物工程(大連)有限公司。
1.3 主要試劑 病毒DNA/RNA 提取試劑盒,購自廣州美基(欣研)生物科技有限公司;PrimeScipt?One Step RT-PCR kit、DNA Marker DL-2 000,均購自寶日醫(yī)生物技術(北京)有限公司;膠回收試劑盒為Promega 公司產(chǎn)品;EB 替代染料,購自廣州華奇盛生物科技有限公司。
1.4 引物設計與合成 參照GenBank 中收錄的JXA1 和VR-2332 等毒株基因序列,應用Primer Premer5.0 分別設計2 對NSP2 和ORF5 基因的特異性引 物。 NSP2-F:5′-CCTCCGTGGTGCAACAAATCTTG-3′; NSP2-R: 5′-CGATGATGGCTTGAGCTGAGTAT-3′,擴增片段大小為1 064 bp。ORF5-F:5′-GTGTCAGGCATTGTGGCTGTG-3′;ORF5-R:5′-CATTATTGGCGTGTAGGTGATTAGAAAA-3′,擴增片段大小為821 bp。引物由蘇州金唯智生物科技有限公司合成。
1.5 病料總RNA 的提取及RT-PCR 擴增 取0.5 g左右無菌研磨好肺、脾等病料組織,置于1.5 mL 離心管內,以1 mL 無菌PBS 進行重懸,反復3 次凍融后,8 000 r/min 離心10 min,取上清液按照病毒RNA提取試劑盒說明書提取病料組織病毒RNA。以提取的RNA 為模板,利用PrimeScipt?One Step RTPCR kit 對目的片段進行擴增。擴增體系為50 μL,PrimeScript 1 step Enzyme Mix 2 μL,2×1 Step Buffer 25 μL,上游引物2 μL,下游引物2 μL,RNA 模板4 μL,RNase Free ddH20 15 μL。PCR 反應循環(huán)參數(shù)為:50 ℃30 min;94 ℃2 min;94 ℃30 s,60 ℃30 s,72 ℃90 s,35 個循環(huán);72 ℃延伸10 min。取5 μL 反應產(chǎn)物進行瓊脂糖凝膠電泳觀察。
1.6 NSP2 和ORF5 基因的克隆及測序 PCR 產(chǎn)物凝膠回收后克隆于pMD-19T 載體上,轉化DH5α 感受態(tài)細胞,陽性重組質粒送蘇州金維智生物科技有限公司測序。
1.7 NSP2 和ORF5 基因的遺傳變異分析 參考GenBank 收錄的國內外的PRRSV 參考毒株序列,利用Lasergene 軟件中MegAlign 程序對不同毒株的Nsp2 和ORF5 基因序列和推導氨基酸進行比對和同源性分析;運用MEGA5.0 軟件的Maximum Likelihood 方法繪制發(fā)育遺傳進化樹。
2 結果
2.1 RT-PCR 檢測PRRSV 結果 2016-2017 年共采集檢測來自廣東各地PRRSV 病料906 份,PRRSV檢測為陽性的樣本有399 份,陽性率為44%,其中2月份與9 月份PRRSV 核酸陽性率最低,約10%,8月份陽性率最高,達到了65%。提示PRRS 在廣東流行的情況比較嚴重,不同月份之間檢測差異應該與廣東的氣候和豬場免疫程序有很大關聯(lián)。從陽性樣品中挑選9 份,編號分別為:GDGZ、GDFS、GDSH、GDHY、GDHZ-1、GDHZ-2、GDMZ、GDYJ、GDSG進行NSP2、ORF5 基因變異分析。
2.2 NSP2 基因的變異特征
2.2.1 NSP2 基因核苷酸及其推導氨基酸序列比對分析 9 個NSP2 基因核苷酸序列之間同源性為88.9% ~100%,推導的氨基酸序列同源性為87.0%~100%;與歐洲型代表株LV 核苷酸同源性僅為40.1%~43.7%,推導氨基酸序列同源性為16.1%~18.0%;與經(jīng)典美洲株NADC30 核苷酸同源性為56.3%~60.0%,推導氨基酸序列同源性為49.5%~52.4%;與美洲型代表株VR-2332 的核苷酸同源性為74.4%~77.1%,推導氨基酸序列同源性為66.8%~68.4%;與近年一度在南方流行低致病性代表株GM2 核苷酸同源性為75.7%~79.5%,推導氨基酸序列同源性為67.0%~70.7%;與中國經(jīng)典株CH-1a 核苷酸同源性為85.9%~90.8%,推導氨基酸序列同源性為82.7%~88.2%;與中國高致病性代表株JXA1 和HUN4 核苷酸同源性分別為92.3%~98.3%和92.2%~98.2%,推導氨基酸序列同源性分別為91.5% ~98.1%和91.5% ~97.8%。結果表明,9 個NSP2 基因之間高度同源,與歐洲型之間同源性最低,與經(jīng)典美洲株NADC30、美洲型代表株VR-2332 和低致病性代表株GM2 之間同源性不高,與中國經(jīng)典株CH-1a 之間同源性較高,與中國高致病性代表株JXA1 和HUN4 同源性最高。
9 個NSP2 基因推導氨基酸序列與HP-PRRSV代表株高度相似,但與其他代表株相比,出現(xiàn)了多處多個氨基酸位點的突變(圖1)。其中GDFS、GDYJ、GDSH、GDSG、GDMZ、GDHZ-1、GDHZ-2 共7 個毒株與JXA1 和HUN4 相比,具有相同的氨基酸缺失特征,即有1+29 個氨基酸的缺失,如圖1 實線方框所示,也與文獻報道的HP-PRRVS的NSP2 氨基酸缺失特征一致。 GDHY 和GDGZ株在相應位置發(fā)現(xiàn)缺失8+1+0 的特征,與CH-1a 的缺失特征相似,如圖1 橢圓線框所示。從圖中我們也看到NADC30 株出現(xiàn)了與文獻報道一致的(111+1+19)131 個不連續(xù)氨基酸的缺失特征,但本次調查并沒有發(fā)現(xiàn)與之缺失情況相似的毒株,即目前國內外報道較多的NADC30 或NADC30-like 株。2.2.2 NSP2 基因核苷酸序列的遺傳進化分析 由9 個毒株與參考毒株的NSP2 基因序列構建的進化樹顯示,美洲型PRRSV 可分為5 個亞群(圖2),即以HP-PRRSV JXA1、TJ 和HUN4 等為代表的亞群1,以中國經(jīng)典株CH-1a 為代表的亞群2,以國內近年在華南和港臺地區(qū)流行的低致病性毒株GM2 為代表的亞群3,以VR-2332 為代表的亞群4,以及以經(jīng)典毒株NADC30 為代表的亞群5[9-10]。而9 株廣東地區(qū)PRRSV 毒株均屬于亞群1,表明近兩年廣東省流行的PRRSV 仍以高致病性毒株為主。
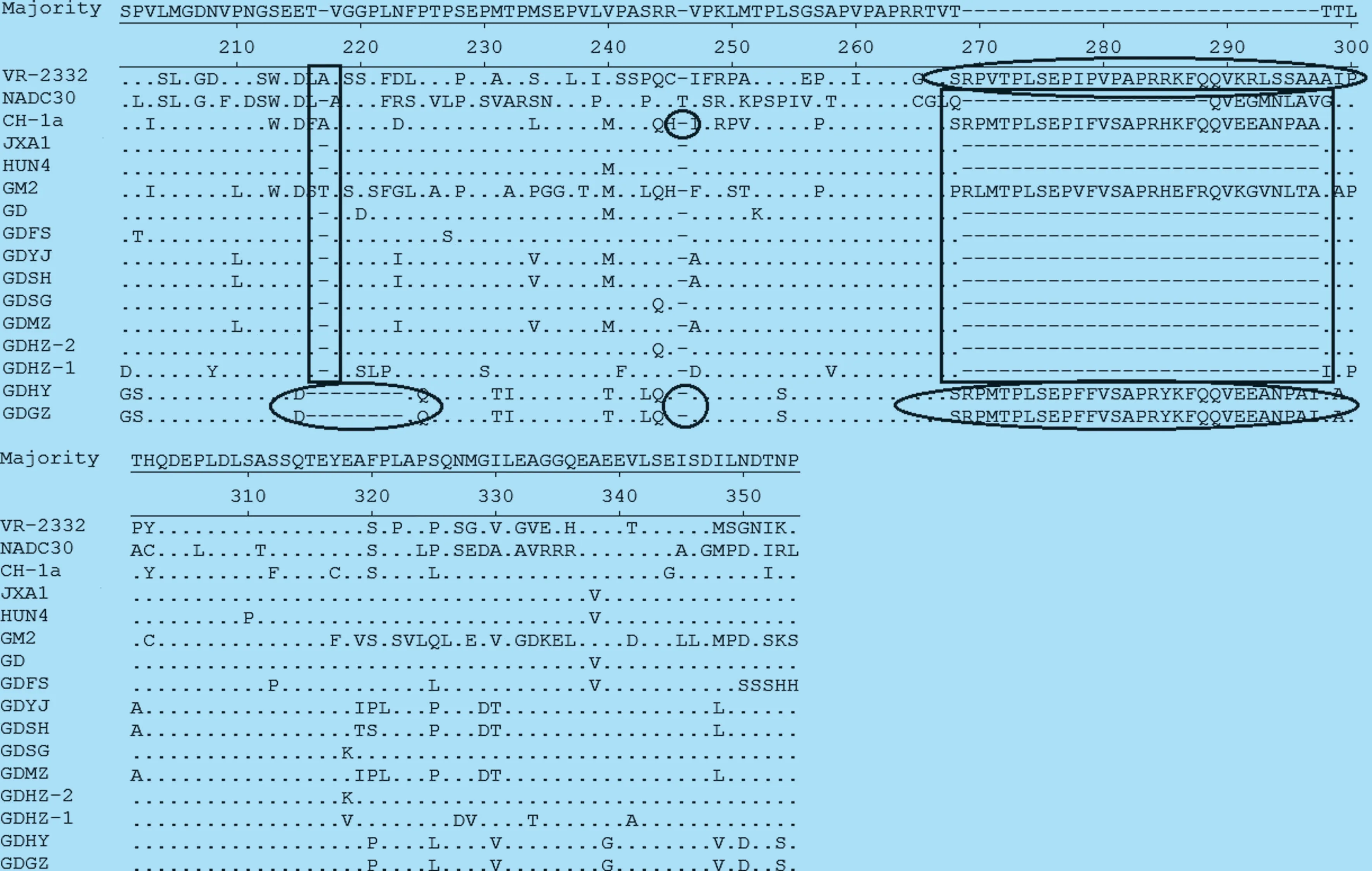
圖1 NSP2 蛋白推導氨基酸序列比對分析
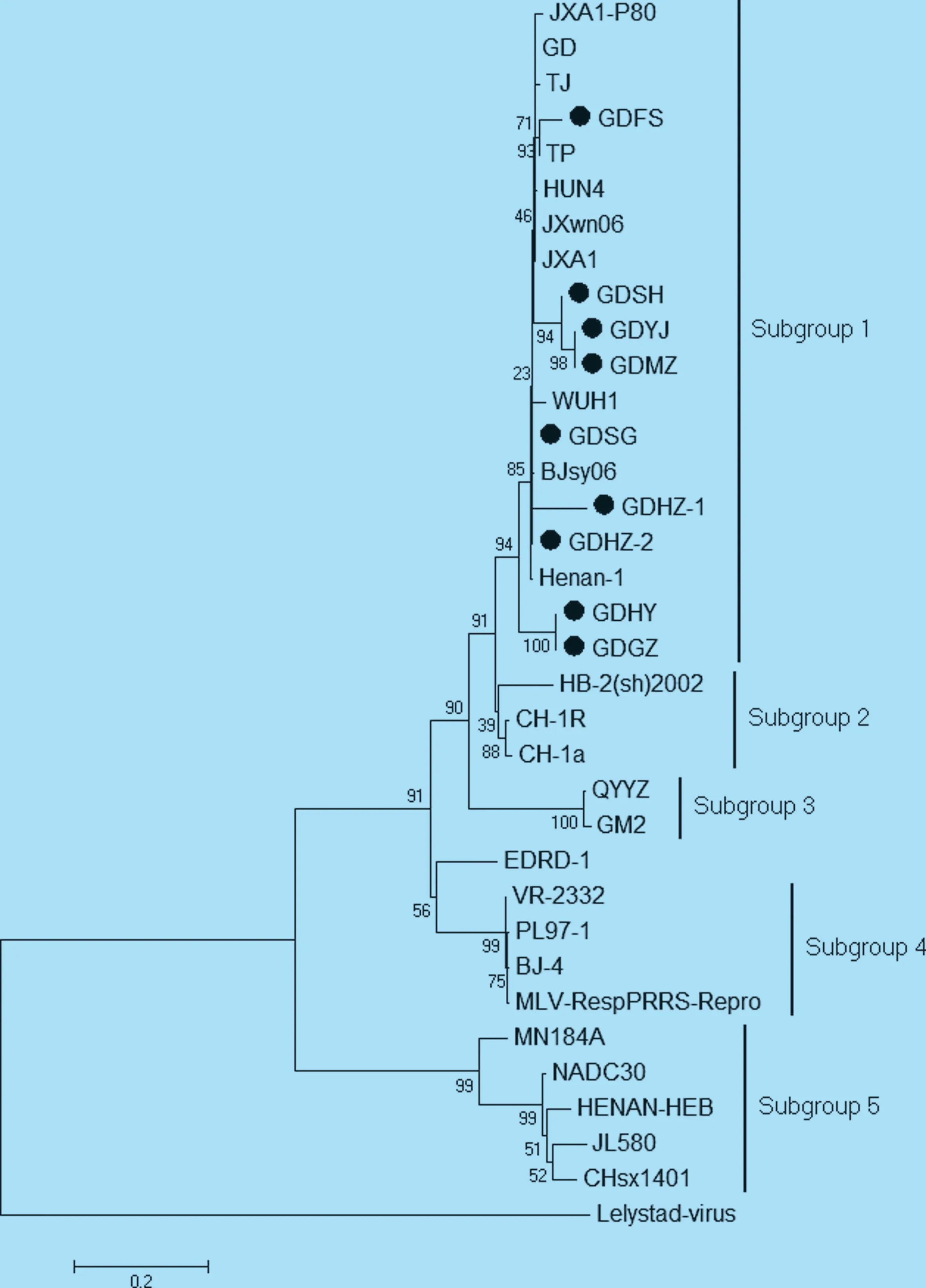
圖2 NSP2 基因核苷酸序列遺傳進化分析
2.3 ORF5 基因的變異特征
2.3.1 ORF5 基因核苷酸及其推導氨基酸序列比對分析 9 個ORF5 基因的核苷酸序列同源性為92.0% ~100%,其推導的氨基酸序列同源性為91.5%~100%;與歐洲型代表株LV 核苷酸同源性為62.6% ~64.3%,推導氨基酸序列同源性為56.6%~59.2%;與經(jīng)典美洲株NADC30 核苷酸同源性為84.2%~86.6%,推導氨基酸序列同源性為83.1%~87.6%;與美洲型代表株VR-2332 的核苷酸同源性為87.2%~90.0%,推導氨基酸序列同源性為84.6%~90.0%;與近年在南方流行的低致病性代表株GM2 核苷酸同源性為81.6%~84.2%,推導氨基酸序列同源性為79.1%~83.6%;與中國經(jīng)典株CH-1a 核苷酸同源性為91.7%~95.5%,推導氨基酸序列同源性為89.1%~93.5%;與中國高致病性代表株JXA1 和HUN4 核苷酸同源性分別為94.2%~99.7%和94.4%~99.8%,推導氨基酸序列同源性為95.0%~99.5%和95.0%~100%。結果表明,9 個NSP2 基因之間高度同源,與歐洲型之間同源性最低,與經(jīng)典美洲株NADC30、美洲型代表株VR-2332 和低致病性代表株GM2 之間同源性略高,與中國經(jīng)典株CH-1a 之間同源性較高,與中國高致病性代表株JXA1 和HUN4 同源性最高。
當前已確定美洲型的PRRSV GP5 蛋白有3 個表位,其中非中和表位2 個(aa27 ~aa30 和aa180 ~aa197),中和表位1 個(aa37 ~aa45),如圖3 黑色框所示。如圖3 所示,本試驗的9 個毒株與參考株比較,在非中和表位aa27 ~aa30,主要是第29 位氨基酸發(fā)生突變,與美洲型代表株VR-2332 相比,GDSG、GDHY、GDGZ、GDFS 與其一致,GDYJ、GDSH、GDMZ、GDHZ-1 和GDHZ-2 的A29V,與CH-1A、JXA1、HUN4N 和ADC30 一致。在aa180 ~aa197 中,在185 位和189 位,9 個毒株與JXA1 和HUN4 一致,但與VR-2332 相比發(fā)生V185A 和I189L;在196 位,GDSH、GDSG、GDMZ、GDHY、GDGZ、GDFS 株與VR-2332、CH-1a 和HUN4 一致,GDYJ、GDHZ-1、GDHZ-2三個毒株發(fā)生Q196R。在aa37 ~aa45,主要突變發(fā)生在第39 位氨基酸,9 個毒株基本與國內高致病毒株JXA1 和HUN4 一致,與VR-2332 和NADC30 相比發(fā)生L39I。總體來說,國內高致病性毒株(含本試驗9 株)與美洲型代表株VR-2332 相比,GP5 蛋白的2 個重要抗原相關區(qū)域的4 個氨基酸位點(29、39、185 和189)的變異較為明顯:A29V、L39I、V185和AI189L。提示這種變化或與兩者之間毒力差異相關,但要做出定論還需進一步研究。

圖3 GP5 蛋白推導氨基酸序列比對分析
2.3.2 ORF5 基因核苷酸序列的遺傳進化分析由9 個毒株與參考毒株的ORF5 基因序列構建的進化樹顯示,與NSP2 進化樹相似,美洲型PRRSV 也可分為5 個亞群(圖4)[9-10]。9 株廣東地區(qū)PRRSV毒株均屬于亞群1,表明近兩年廣東省流行的PRRSV 仍以高致病性毒株為主。
3 討論
HP-PRRSV 近年來在我國大部分地區(qū)流行甚至暴發(fā),使我國養(yǎng)豬業(yè)損失慘重,大量的政府主導強制免疫的HP-PRRSV 活疫苗涌入市場,使得弱毒疫苗有可能在選擇壓作用下發(fā)生毒力返強[11]。本試驗結果顯示,PRRSV 核酸陽性率達到44%,抽檢的9 個毒株均與高致病毒株高度同源且處于同一亞群,在如此高強度免疫的狀態(tài)下存在這種情況,提示可能為疫苗株返強。另外,弱毒疫苗株和野毒株同時在豬群中存在,也使得毒株之間發(fā)生重組的風險增加[12]。
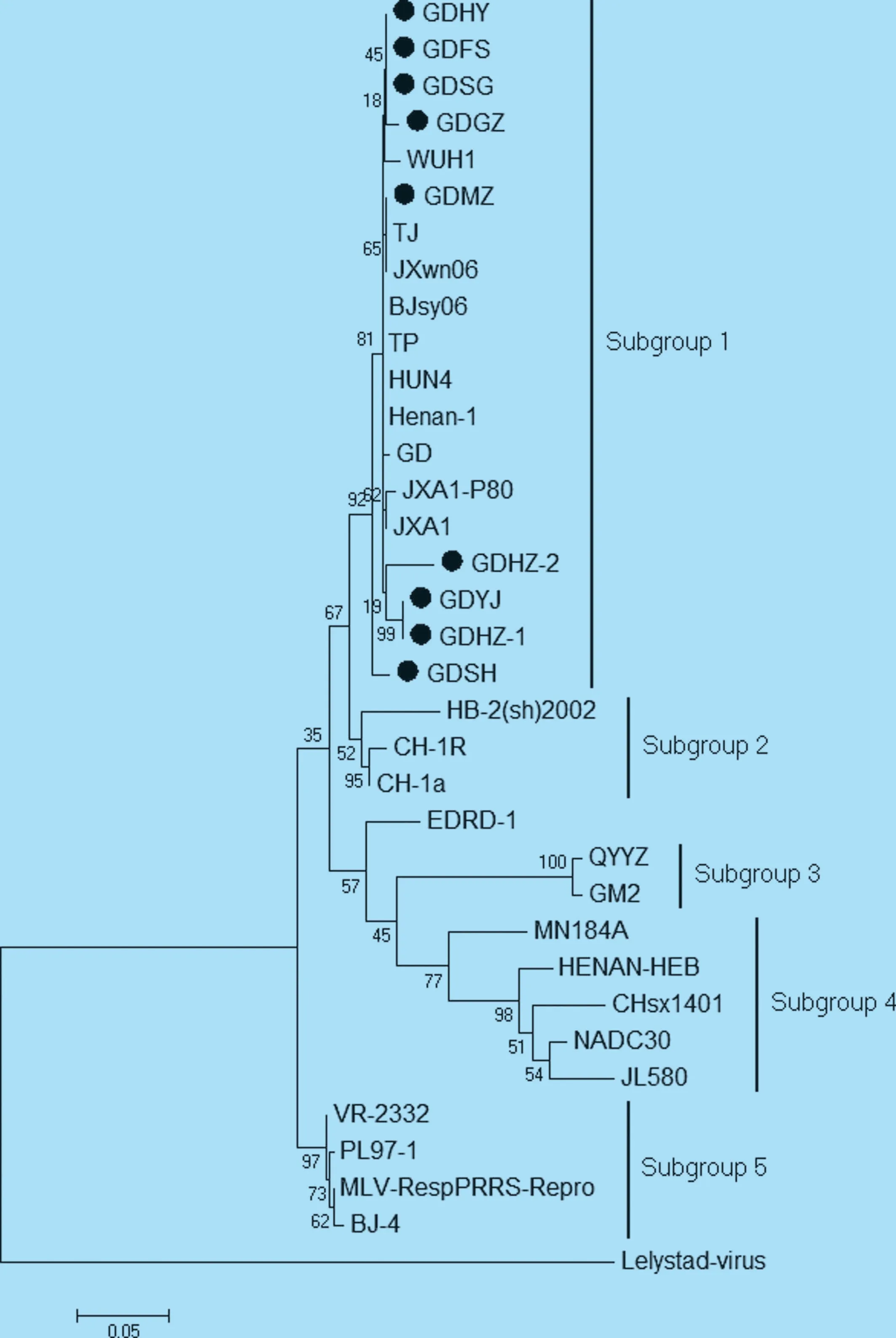
圖4 ORF5 基因核苷酸序列遺傳進化分析
本研究通過對2016-2017 年間采集自廣東各地的906 份疑似病料進行了RT-PCR 檢驗,測得陽性率為44%,為了進一步研究PRRSV 在廣東分子流行病學情況和毒株基因變異的發(fā)展,隨機挑選9個陽性樣本進行NSP2 和ORF5 基因序列及推導氨基酸比對,并對遺傳序列進行分析。結果顯示,9 個樣品均屬于HP-PRRSV,表明廣東流行的PRRSV 仍以高致病性毒株為主,這也與于林洋等[13]2016-2017 年在廣東、李海琴等[14]2013-2014 年在江西和李彬等[15]2013-2014 年在華中地區(qū)調查的結果一致,表明HP-PRRSV 仍是南方地區(qū)的優(yōu)勢毒株。但周峰等[16]、郭天準等[17]和董建國等[18]2012-2015 年間在河南調查的結果發(fā)現(xiàn)除HP-PRRSV 之外,還出現(xiàn)了與NADC30 高度同源的新變異株-NADC30-like 株的流行,該變異株與國內HP-PRRSV株有重組現(xiàn)象,且致病力有所增強[19],李真等[20]2014-2016 年在華北的地區(qū)的調查也發(fā)現(xiàn)了相同的情況,而且還發(fā)現(xiàn)新的變異毒株GM2-like 株的存在。以上結果似乎表明中國南北方流行的毒株有所差異,提示近區(qū)域引種或精液交易可能是造成這種差異的原因,但具體原因有待進一步研究。總之,本研究調查發(fā)現(xiàn)豬場在普遍使用了PRRSV 弱毒活疫苗后,檢出陽性率仍然較高,而且不斷有新的變異株出現(xiàn),說明目前市場的疫苗不能很好地控制該病的發(fā)生,一方面說明目前中國PRRSV 毒株呈多樣化趨勢,另一方面提示PRRSV 在不斷的發(fā)生變異,究竟是毒力返強還是不同毒株之間的重組,或是病毒抗原表位突變導致免疫逃逸,以及這些變異與其毒力和致病力之間的相關性還需進行深入研究。
