澳洲堅果不定芽誘導及生根培養研究
2019-08-30 07:08:10鄒家通羅一然韓國偉何小帆和潤喜何承忠
西南林業大學學報 2019年5期
鄒家通 羅一然 韓國偉 何小帆 和潤喜 何承忠,3
( 1. 西南林業大學云南省高校林木遺傳改良與繁育重點實驗室,云南 昆明 650233;2. 西南林業大學西南地區生物多樣性保育國家林業局重點實驗室,云南 昆明 650233;3. 西南林業大學西南山地森林資源保育與利用教育部重點實驗室,云南 昆明 650233)
澳洲堅果(Macadamia ternifolia)又稱為澳洲胡桃、夏威夷果、昆士蘭堅果、巴布果,屬山龍眼科(Proteaceae)澳洲堅果屬(Macadamia)的常綠喬木。原產于澳大利亞東南部的昆士蘭洲沿海地區及新南威爾士洲北部的江河地區,因其具有豐富的營養和獨特的風味而備受消費者青睞,且價格昂貴,具有很大的市場潛力和很高的經濟價值、科研價值。我國自20世紀70年代引進澳洲堅果品種并開展了環境適應性試驗,至今已成為澳洲堅果發展最快、種植面積最大的國家[1]。目前,澳洲堅果苗木繁殖方法主要采用扦插育苗和嫁接育苗[2]。
組織培養快繁技術不僅能夠實現植物優良品種的工廠化育苗,在較短時間內達到規模化生產苗木,且組培再生體系的建立也是開展植物分子育種的前提和基礎[3]。Gitonga等[4]探討了澳洲堅果組織培養的基本條件,并在WPM培養基上以幼嫩莖段為外植體獲得了高達98%的萌芽率,但進一步嘗試生根誘導時,沒有獲得成功。Cha-um等[5]以蛭石為載體,向培養容器中充入高濃度CO2,從而獲得了根系發達的粗殼澳洲堅果品種"Keaau"的生根苗。國內有關澳洲堅果組培技術報道較少,肖再云[6]利用莖段培養愈傷組織和畸胚,再進一步培養分化獲得不定芽,剪下單芽誘導生根培養60 d,再將這些無根苗移入沙床中煉苗60 d,最終獲得了生根苗。郭凌飛等[7]在澳洲堅果成年樹上剪取帶芽莖段進行組織培養研究,獲得了萌芽率較好的培養基配方,但未能成功獲得生根苗。由此可見,目前國內澳洲堅果組織培養技術尚不成熟,限制了澳洲堅果遺傳改良、良種快繁及現代生物技術育種等工作[8]。澳洲堅果是多年生木本植物,其外植體存在脫毒難、易褐化、生根誘導困難等問題。本研究以澳洲堅果‘H2’品種幼苗的帶腋芽莖段為外植體材料,通過開展不同植物生長調節劑及其配比對腋芽啟動、增殖及生根的影響研究,從而建立其組織培養快繁技術體系,為澳洲堅果優良品種苗木規模化繁育、人工多倍體誘導育種及現代生物技術育種等研究提供參考。
1 材料與方法
1.1 材料來源
供試材料為德宏州林業局苗圃中心引種栽培的澳洲堅果‘H2’品種2年生實生苗,將苗木帶回昆明并栽種于西南林業大學格林溫室,待苗木抽生新梢后,取著生有側芽的幼嫩莖段為外植體。
1.2 試驗方法
1.2.1 外植體消毒
將剪取的幼嫩莖段切除葉片,用洗潔精溶液清洗其表面的泥土及灰塵,之后置于燒杯中流水沖洗1 h。在超凈工作臺上先用75%乙醇浸泡20 s,再用無菌水沖洗4次,5%次氯酸鈉浸泡3 min,0.1%升汞處理5 min,用無菌水沖洗5次,無菌濾紙吸干表面水分,備用。
1.2.2 腋芽啟動培養
將消毒處理后的外植體截取長度為2.5~3.0 cm的帶腋芽莖段,將其豎直插入腋芽啟動培養基中。腋芽誘導培養基設計的因素及水平為:6-BA 1.0、2.0、3.0 mg/L; IBA 0、0.5、1.0 mg/L;NAA 0、0.5、1.0 mg/L;活性炭(AC)0、200、400 mg/L。以WPM為基本培養基,添加20 g/L蔗糖和4 g/L瓊脂。采用L9(34)正交試驗探索不同濃度6-BA、IBA、NAA、AC對腋芽啟動培養的影響。試驗共9個處理,每個處理接種5瓶,每瓶接種3個外植體,3次重復。接種后定期觀察和記錄腋芽萌發情況,15 d后統計腋芽誘導系數。
1.2.3 腋芽增殖培養
待腋芽萌發生長至1~2 cm時,將其切下接種于不定芽分化的增殖培養基中。增殖培養基以MS為基本培養基,加入30 g/L蔗糖和4 g/L瓊脂,分別添加2.0 mg/L的TDZ、2.0 mg/L的6-BA、0.5 mg/L或 1.0 mg/L的 NAA以及 0.5 mg/L或1.0 mg/L的KT。共設計8個處理組合,每個處理組合接種5瓶,每瓶接入3個單芽,3次重復,培養45 d后統計不定芽增殖系數。
1.2.4 生根誘導培養
待不定芽生長高度達到2 cm左右時,剪取生長健壯的不定芽,接種于生根培養基誘導生根。生根培養以WPM、1/2MS為基本培養基,加入30 g/L蔗糖,4 g/L瓊脂,200 mg/ L的AC,植物生長調節劑6-BA 0.2 mg/L,NAA 1.0 mg/L,IBA(0.5、1.0 mg/L),ABA 0.5 mg/L,調節培養基酸度分別為pH=5.0和pH=6.0兩個水平。共設置8個處理組合,每個處理組合接種5瓶,每瓶接種3個不定芽,3次重復。培養40 d后統計生根率,并觀察組培苗生長狀況。
1.2.5 煉苗移栽
生根組培苗苗高達到4~5 cm時即可進行煉苗。在培養室內打開瓶蓋,放置3~4 d,然后移至室溫下放置7 d,取出生根苗洗凈粘附在根部的培養基,移栽到用多菌靈消毒后的基質中(珍珠巖、河沙、紅土、腐殖土4種基質按不同比例混合配制),移栽后定期采用霧狀化噴水保濕,于14 d后統計成活率。
1.2.6 培養條件
培養條件為晝溫(25±2)℃,夜溫(22±2)℃,光照強度2 000~2 500 lx,光照時間12 h/d。
1.2.7 數據統計與分析
采用Excel 2007進行數據整理,運用SPSS 22.0軟件對試驗數據進行統計分析,多重比較采用Duncan’s新復極差測驗法(P<0.01)進行顯著性分析。
主要指標計算公式如下:

2 結果與分析
2.1 植物生長調節劑配比對腋芽萌發的影響
將消毒外理后的帶腋芽幼嫩莖段接入含不同濃度植物生長調節劑配比的培養基中,15 d后腋芽萌發(圖1a)及生長狀況見表1。由表1可見,處理組合AG6為最優組合,萌發的腋芽生長健壯,葉片翠綠舒展,且腋芽誘導系數達到3.51,極顯著地高于其他處理組合,表明植物生長調節劑6-BA與IBA按一定濃度配比使用時,對澳洲堅果腋芽萌發具有顯著的促進作用,且有利于萌發后的腋芽生長。處理組合AG1的腋芽誘導系數最低,僅為0.42,說明在WPM基本培養中僅添加植物生長調節劑6-BA,對澳洲堅果腋芽萌發和腋芽生長的作用效果不明顯。然而,當6-BA使用濃度為3.0 mg/L時,無論與IBA、NAA共同配比使用,還是僅與IBA或NAA配比使用,雖然對腋芽萌發具有較好的促進作用,但萌發抽生的腋芽出現較嚴重的玻璃化和葉片畸形現象(處理組合AG7,AG8和AG9)。此外,在培養基中添加適量的活性炭(AC),腋芽生長更為健壯,葉色翠綠。因此,結合腋芽生長狀況和誘導系數,從節約成本方面考慮,較適宜澳洲堅果莖段腋芽萌發的培養基為WPM+ 6-BA 2.0 mg/L+IBA 1.0 mg/L+AC 200 mg/L。

圖 1 澳洲堅果不定芽誘導與生根培養Fig. 1 Adventitious bud induction and rooting culture of M. ternifolia
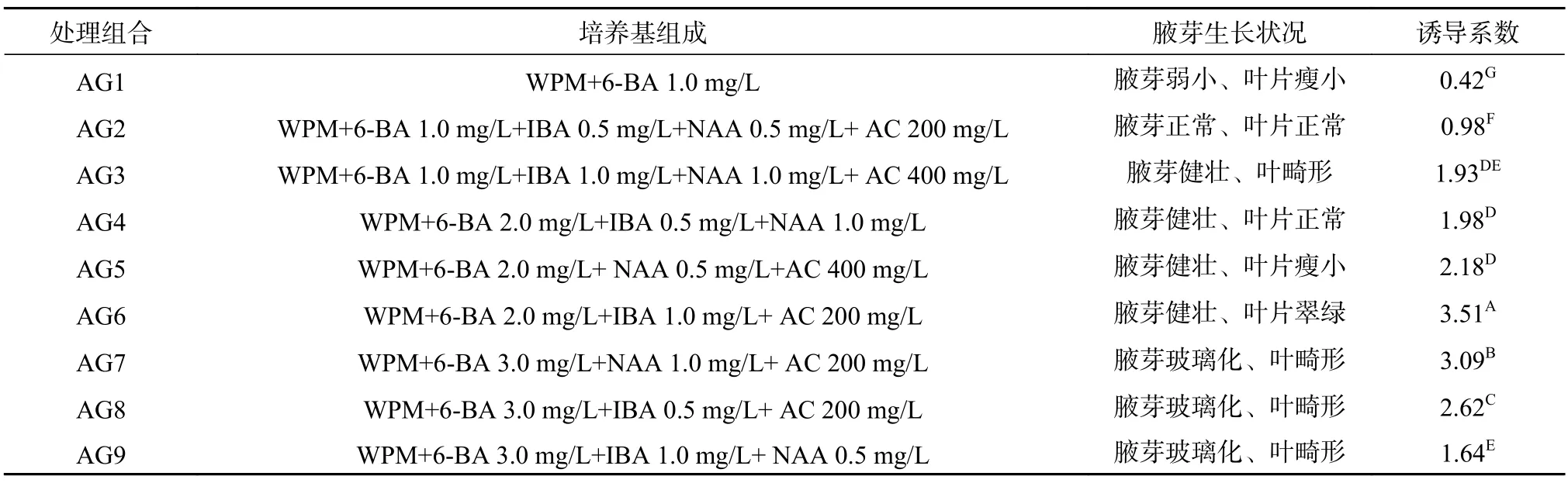
表 1 不同植物生長調節劑配比對腋芽萌發的影響Table 1 Effects of different exogenous hormones ratio on the axillary bud germination
2.2 植物生長調節劑配比對腋芽增殖的影響
將生長健壯,高度達到2.0 cm以上的腋芽從基部切下,豎直接種于增殖培養基,45 d后統計增殖系數及不定芽生長情況(圖1b和表2)。由表2可知,不同處理組合間的不定芽增殖系數呈極顯著差異(P<0.01)。其中處理組合AP7的增殖系數最高(3.49),其次是處理組合AP3,增殖系數為3.09,二者之間差異不顯著,且誘導發生的不定芽生長勢良好,而TDZ + KT/NAA組合(AP1,AP2,AP5和AP6)不僅增殖系數低,而且誘導發生的不定芽生長勢一般,表明6-BA +KT/NAA組合更適宜于澳洲堅果腋芽的增殖培養。此外,澳洲堅果不定芽對NAA比較敏感。當NAA濃度為0.5 mg/L時(處理組合AP1,AP3),不定芽生長良好;而NAA濃度增加為1.0 mg/L時(處理組合AP2,AP4),則引起不定芽的玻璃化。植物生長調節劑KT與TDZ組合對澳洲堅果不定芽也存在相同的作用效果,但KT與6-BA組合中(處理組合AP7,AP8),KT濃度為0.5 mg/L或1.0 mg/L時均沒有出現玻璃化現象。綜合上述分析認為,澳洲堅果腋芽增殖的適宜培養基為MS+6-BA 2.0 mg/L+KT 0.5 mg/L。
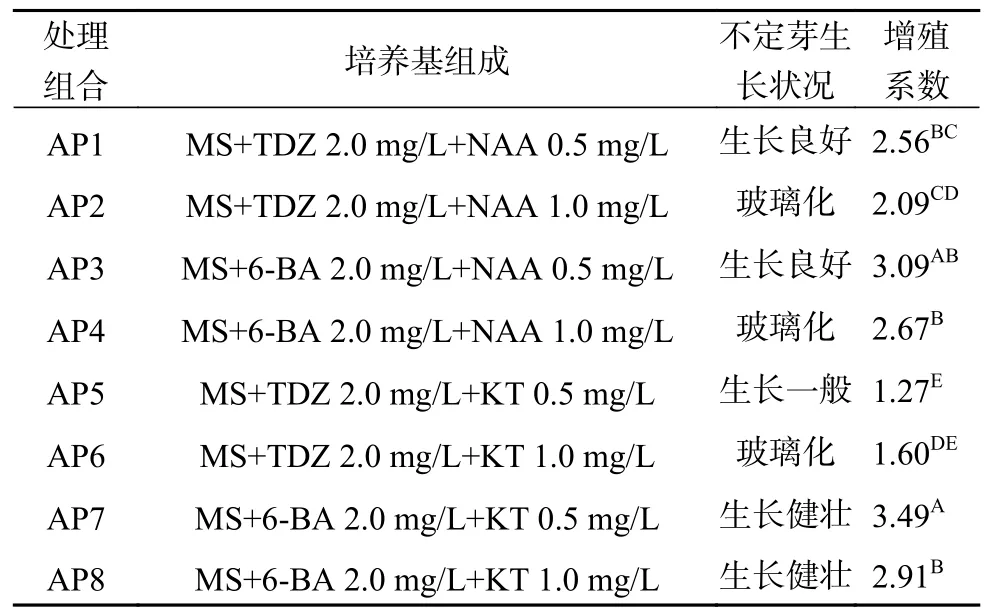
表 2 不同植物生長調節劑配比處理對腋芽增殖的影響Table 2 Effects of different exogenous hormones ratio on proliferation of axillary bud
2.3 植物生長調節劑配比對誘導生根的影響
剪取生長健壯的不定芽,接種于生根培養基誘導生根(圖1c),培養40 d后不定芽生根結果見表3。結果顯示,當培養基pH=6.0時,僅有3個處理組合(R2,R6,R8)誘導不定芽產生不定根,生根率最高值為16.3%;當培養基pH=5.0時,除R1處理組合外,其他7個處理組合均能夠誘導不定芽產生不定根,生根率最高值達到42.2%,表明生根培養基pH=5.0更適宜于澳洲堅果不定芽的生根培養。在植物生長調節劑配比相同的情況下,以1/2MS為基本培養基的處理組合生根率均顯著高于以WPM為基本培養基的處理組合生根率,說明1/2MS基本培養基較WPM基本培養基更適宜用于澳洲堅果不定芽的生根培養。進一步分析發現,接種于添加有ABA成分的培養基(處理組合R2和R6),不定芽生根率顯著高于其他培養基,表明植物生長調節劑ABA對澳洲堅果不定芽誘導生根具有促進作用。由此可知,可用于澳洲堅果不定芽誘導生根的培養基為1/2MS+6-BA 0.2 mg/L+IBA 0.5 mg/L+ABA 0.5 mg/L,pH=5.0。
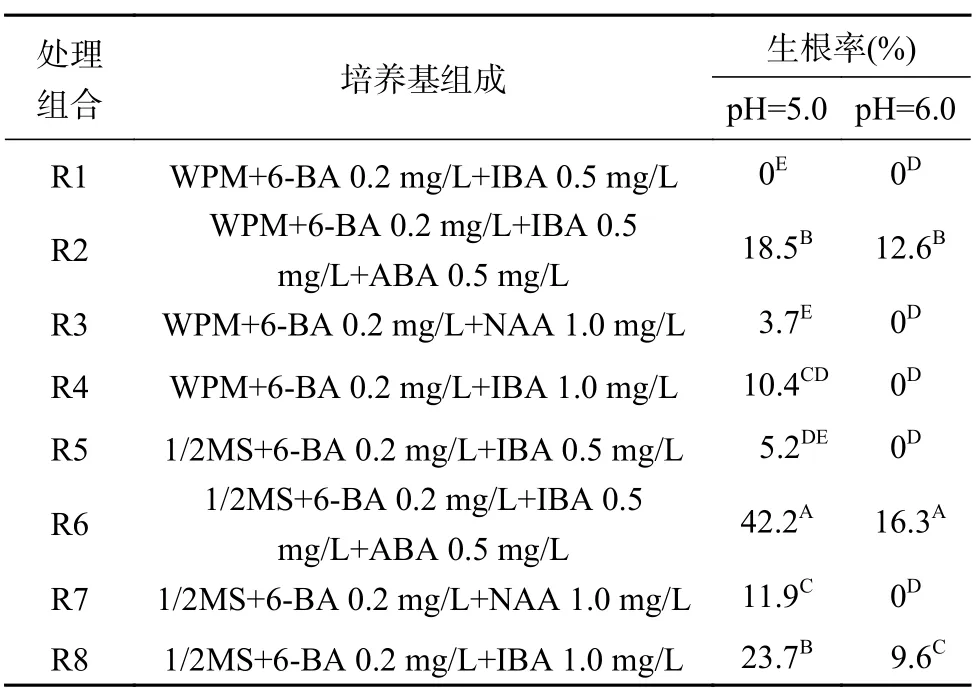
表 3 不同植物生長調節劑配比對生根誘導的影響Table 3 Effects of different exogenous hormones ratio on rooting induction
2.4 基質對組培苗移栽成活的影響
將澳洲堅果組培苗在培養室內開瓶煉苗3 d,移至實驗室內室溫下煉苗5 d后,移栽于不同基質中(圖1d),14 d后統計成活情況(表4)。由表4可見,組培苗移栽成活率最高者是配比組合S2,成活率達到66.7%,其次是配比組合S1,移栽成活率為66.3%,移栽成活率最低者是配比組合S3(53.3%),但各配比組合基質間的組培苗移栽成活率差異不顯著(P=0.397>0.05),而含有河沙的栽種基質有利于組培苗成活。由此,V(河沙)∶V(紅土)∶ V(腐殖土)=1∶ 1∶ 1的基質適用于澳洲堅果組培苗的移栽。

表 4 不同基質對組培苗成活的影響Table 4 Effects of different substrates on survival rate
3 結論與討論
植物細胞在理論上都存在全能性,任何植物的器官或組織都能作為組織培養的外植體[9]。但在組織培養實踐中,植物不同品種、組織或器官,其分化能力存在顯著差異,而且外植體的生理年齡、生長階段與取材部位等都直接影響著組織培養技術體系的建立,因此,外植體選擇是組織培養能否成功的首要因素[10]。在很多木本植物組織培養中,選擇以葉片、葉柄作為外植體,均能獲得較好的誘導效果。在預試驗時,采集‘H2’澳洲堅果的幼嫩葉片、葉柄和幼嫩莖段作為外植體開展誘導研究,但葉片和葉柄的分化能力差,均在30 d后逐漸褐化、變黃死亡,只有以幼嫩莖段作為外植體獲得了較好的誘導效果。
澳洲堅果組培苗誘導生根較困難。郭凌飛等[7]和Bhalla等[11]在研究澳洲堅果組織培養時,曾嘗試添加多種植物生長調節劑進行誘導生根,但均未獲得理想效果。Cha-um等[5]將澳洲堅果不定芽接入以蛭石為載體的MS液態培養基中,并向培養容器中注入高濃度CO2,成功誘導出了不定根。一般地,WPM培養基更適合于木本植物的組培苗生根培養,但本研究結果表明,‘H2’澳洲堅果組培苗在1/2MS培養基中的誘導生根效果優于WPM培養基。
ABA作為植物天然的生長抑制物質,對植物形態發生和生長具有負效應[12],并對植物生根發揮著關鍵作用[13-14]。赤霉素能夠抑制植株生根培養起始階段的細胞分裂,從而抑制根原基的分化和形成[15]。作為赤霉素的天然拮抗劑[16],較高濃度的ABA在不定根的分化階段發揮作用,能夠促進植物不定根的形成[14]。本研究中也發現,在不定芽誘導生根培養基中添加適量植物生長調節劑ABA,能夠顯著地提高澳洲堅果不定芽的生根率,與前人研究結果一致。
此外,培養基的酸堿度對澳洲堅果不定芽生根誘導有一定的影響,偏酸性(pH=5.0)的培養基有利于不定根的發生。總之,本研究僅初步建立了澳洲堅果的組織培養快繁技術體系,組培苗生根率較低,僅為42.2%,還需要進一步優化完善。
[ 參 考 文 獻 ]
[1]賀熙勇, 陶亮, 柳覲, 等. 我國澳洲堅果產業概況及發展趨勢 [J]. 熱帶農業科技, 2015, 38(3): 12-16.
[2]羅培四, 何新華, 韋巧云, 等. 澳洲堅果繁育技術研究進展 [J]. 中國南方果樹, 2017, 46(3): 179-183.
[3]Oliveira J P S, Hakimi O, Murgu M, et al. Tissue culture and metabolome investigation of a wild endangered medicinal plant using high definition mass spectrometry [J]. Plant Cell, Tissue and Organ Culture,2018, 134(1): 153-167.
[4]Gitonga L, Gichuki S, Ngamau K, et al. Effect of explant type, source and genotype on in vitro shoot regeneration in Macadamia (Macadamia spp.) [J]. Agricultural Biotechnology and Sustainable Development,2010, 2(7): 129-135.
[5]Cha-um S, Chanseetis C, Chintakovid W, et al. Promoting root induction and growth of in vitro macadamia(Macadamia tetraphylla "Keaau") plantlets using CO2-enriched photoautotrophic conditions [J]. Plant Cell, Tissue and Organ Culture, 2011, 106: 435-444.
[6]肖再云. 澳洲堅果的組織培養研究 [J]. 熱帶農業科學, 1999(4): 12-14.
[7]郭凌飛, 彭靖茹, 覃劍峰, 等. 澳洲堅果組織培養研究初報 [J]. 中國農學通報, 2010(26): 385-386.
[8]柳覲, 孔廣紅, 倪書邦, 等. 澳洲堅果多倍體的化學誘導及流式細胞術鑒定 [J]. 熱帶作物學報, 2013,34(11): 2138-2144.
[9]王小箐, 李玲. 植物生長調節劑在植物組織培養中的應用 [M]. 北京: 化學工業出版社, 2002: 36.
[10]Rout G R, Sahoo S. In vitro selection and plant regeneration of copper-tolerant plants from leaf explants of Nicotiana tabacum L. cv. 'Xanthi' [J]. Plant Breeding,2007, 126(4): 403-409.
[11]Bhalla P L, Mulwa R M S. Tissue culture and macadamia propagation [J]. Acta Horticulturae, 2003, 616:343-346.
[12]費學謙, 闕國寧, 周立紅. ZT、ABA等內源物質與杉樹齡及其組培復壯的關系 [J]. 林業科學研究, 1992,5(3): 317-320.
[13]郭敏敏, 王清連, 胡根海. 利用高效液相色譜法分離和測定棉花組織培養過程種4種內源激素 [J]. 生物技術通訊, 2009, 20(2): 213-216.
[14]鄭均寶, 劉玉軍, 裴保華, 等. 幾種木本植物插穗生根與內源IAA, ABA的關系 [J]. 植物生理學報, 1991,17(3): 313-316.
[15]孟慶偉, 高輝遠. 植物生物學 [M]. 2 版. 北京: 中國農業出版社, 2017: 199-240.
[16]周燕, 高述民, 李鳳蘭. 胡楊不定根原基發生的分生細胞結構特征及內源激素變化分析 [J]. 西北植物學報, 2009, 29(7): 1342-1350.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
