河川沙塘鱧GHR和IGF-2基因克隆及m RNA的時序表達
2019-09-03 07:57:34朱文旭張宏葉張紅燕尹紹武陳樹橋周國勤
海洋漁業 2019年4期
關鍵詞:生長
朱文旭,張宏葉,王 濤,王 丹,張紅燕,尹紹武,陳樹橋,周國勤
(1.南京師范大學生命科學學院,江蘇省生物多樣性與生物技術重點實驗室,南京 210023;2.南京市水產科學研究所,南京 210036)
魚類既是自然界重要的生物資源,也是人類主要食物蛋白來源之一。魚類的生長是一個相當復雜而又系統的過程,受到多種激素的調節,其中生長激素和類胰島素生長因子軸(GHR/IGF軸)作為控制機體生長、發育、代謝的重要組件發揮著不可替代的作用。GHR/IGF軸由生長激素(GH)、生長激素結合蛋白(GHBP)、生長激素受體(GHR)、胰島素樣生長因子(IGF)、胰島素樣生長因子受體(IGFR)以及胰島素樣生長因子結合蛋白(IGFBP)等基因組成[1]。
GH是一種由腦垂體釋放的激素,能夠促進魚體生長,參與多種合成與代謝過程,其在組織和細胞水平上發揮作用的前提是與靶細胞膜上的GHR結合以介導下游的一系列反應。組織中GHR數量的多少、功能的正常與否將直接影響到GH在組織中效應的發揮[2]。LEE等[3]首次在金魚(Carassius auratus)中克隆出硬骨魚類的GHR基因,迄今為止已有數十種魚類的 GHR cDNA序列被克隆,包括大菱鲆(Scophthalmus maximus)[4]、虹鱒(Oncorhynchusmykiss)[5]、尼羅羅非魚(Oreochromis niloticus)[6]等。這些序列與哺乳動物的GHR序列具有一定的相似性,說明GHR基因的信號轉導機制具有一定的保守性。VINCENT等[7]對 GHR基因的結構分析表明,GHR兩個保守的BOX盒序列中任何一個氨基酸突變都會導致GH的效應喪失,表明它們在GHR介導的GH生理信號的過程中發揮了重要作用。ZHONG等[8]在鯽(Carassius auratus)的研究中表明,禁食處理后,三倍體鯽GHR/IGF軸基因表達量仍顯著高于二倍體鯽,表明三倍體中GH/IGF軸基因的表達升高在三倍體魚生長速度快中起關鍵作用。
胰島素樣生長因子(insulin-like growth factors,IGFs)是生長激素(growth hormone,GH)發揮作用的介質[9],主要在肝中合成,具有促進有絲分裂、誘導細胞分化、調節生長等作用[10]。IGF家族包括 IGF配體、IGF受體(insulin-like growth factor receptor,IGFR)和6種 IGF結合蛋白(insulin-like growth factor binding protein,IGFBPs)[11]。1978年 RINDERKNECHT等[12]首次在人類血液中分離出IGF-2,此后,IGF-2基因相繼在哺乳類、鳥類、兩棲類等動物中被克隆。在 魚 類 中 已 在 虹 鱒[13-14]、大 麻 哈 魚(Oncorhynchus keta)、銀大麻哈魚(O.kisutch)、斑馬魚(Brachydanio rerio)等[12]中克隆得到 IGF-2的cDNA序列。其轉錄產物在不同魚中的存在形式也不一致。有研究顯示,從羅非魚及馬蘇大麻哈魚(Oncorhynchusmasou)分別檢測到3種IGF-2轉錄產物和1種主要的IGF-2轉錄產物[15-16],對這些不同轉錄產物的結構分析表明,它們很可能是由于不同的剪接方式以及具有多個多聚腺苷酸位點引起的。此外,CHEN等[17]通過對尼羅羅非魚注射IGF-2,結果發現其體質量增加了72%,說明IGF-2能夠促進其生長。
動物的生長軸基因是調節出生后生長的主要內分泌軸線,其表達存在雌雄二態性,這可能是導致動物生長和形態變化雌雄二態性的主要原因。河川沙塘鱧(Odontobutis potamophila)屬鱸形目(Perciformes),沙塘鱧科(Odontobutidae),沙塘鱧屬,在生長上表現出顯著的性別二態性,雄性河川沙塘鱧比同期雌性生長速度快30%以上。目前,關于河川沙塘鱧生長調控分子的研究非常有限,這在很大程度上阻礙了河川沙塘鱧的產業化發展。本研究首次克隆得到河川沙塘鱧GHR/IGF軸基因GHR和IGF-2的全長,采用qRT-PCR技術對其組織分布及胚胎發育不同階段的表達進行了分析,并研究了河川沙塘鱧在不同生長發育階段和不同組織雌、雄魚的生長差異及GHR、IGF-2基因表達差異,旨在從分子水平探討河川沙塘鱧生長相關基因對其生長發育及雌雄生長差異的調控機理,為該魚的高效養殖積累基礎資料。
1 材料與方法
1.1 實驗材料
實驗用魚河川沙塘鱧取自江蘇省南京市水產科學研究所周崗基地,為同一繁殖批次。于玻璃循環缸中暫養7 d,確定健康無病后開始實驗。采用鏡檢觀察的方法確定胚胎發育時期并采集8個發育階段(受精卵期、桑椹胚期、原腸胚期、神經胚期、體節期、口裂期、出膜后1 d、3 d)。另取1齡河川沙塘鱧[體質量(30.0±1.5)g]的9個組織(腦、肝、鰓、肌肉、腎、性腺、脾、心、腸)的材料用于組織表達的研究。雌、雄魚基因表達差異研究則同一繁育批次成魚分別取90、150、210、270、330 d雌、雄河川沙塘鱧,雌、雄魚每時間段各隨機取3尾個體的肝、肌肉、腦組織分裝于1.5 mL離心管中(RNase-free)。所取的全部材料速凍于液氮10 min后于-80℃冰箱保存備用。
1.2 總RNA的提取及cDNA合成
將-80℃凍存的組織取出,用高純總RNA快速抽提試劑盒(離心柱型)(Bioteke,Beijing,China)根據說明書提取河川沙塘鱧組織總RNA,用RNA濃度測定儀測定每個樣品的RNA濃度及純度,用1%瓊脂糖凝膠電泳檢測其完整性。以總RNA為模板,根據反轉錄試劑盒HiScriptTM1st Strand cDNA Synthesis Kit(Vazyme,Shanghai,China)的說明進行反轉錄,獲得cDNA第一條鏈。
1.3 河川沙塘鱧GHR和IGF-2基因全長的克隆
從河川沙塘鱧轉錄組測序的cDNA文庫分別篩選得到654、1 950 bp的IGF-2和GHR基因的cDNA中間 EST序列,5′RACE System for Rapid Amplification of cDNA Ends Version 2.0試劑盒獲得基因的 5′端序列。采用 Clontech公司的SMARTerTMRACE cDNA Amplification Kit試劑盒獲得目的基因的3′端序列。本實驗克隆基因所需引物見表1。
1.4 GHR和IGF-2基因的生物信息學分析
使用DNAman軟件分析基因的生物信息。核苷酸和氨基酸序列的同源性使用BLAST程序(http://blast.ncbi.nlm.nih.gov/Blast.cgi)進行檢索。采用蛋白質分析專家系統(expert protein analysis system,ExPASy)(http://www.expasy.org/)確定其開放閱讀框進而預測氨基酸序列,理論分子量(MW)和等電點(pI)用pI/MW在線工具(http://www.expasy.org/tools/pi_tool.html)進行預測。使用在線軟件 SMART(http://smart.embl-heidelberg.de/)對預測到的氨基酸序列進行結構和功能的分析,SignalP 3.0(http://genome.cbs.dtu.dk/services/SignalP)進行信號肽的預測,使用在線軟件ClustalW2(http://www.ebi.ac.uk/Tools/msa/clustalo/)進行多序列比對。用MEGA 5.0軟件(http://www.megasoftware.net/),重復1000次構建 N-J系統發育樹[18]。
1.5 河川沙塘鱧GHR和IGF-2基因的表達分析
根據已獲得的GHR、IGF-2基因cDNA全長序列分別設計特異性引物,按照qRT-PCR試劑盒SYBR Green Master kit(Roche, Basel,Switzerland)說明書以β-actin作為內參基因分析GHR、IGF-2基因的mRNA表達。反應體系為20 uL,Faststart Universal SYBR GreenMaster 10μL,正反向引物各3μL,cDNA模板4μL。反應程序為95℃ 10 min;95℃ 10 s,55℃ 30 s,40個循環,4℃保存。其中,每條引物配置成濃度為2μmol·L-1的 工 作 液,cDNA 模 板 濃 度 為5 ng·L-1。

表1 克隆GHR和IGF2基因及實時熒光定量PCR所用引物Tab.1 Primers used for charactering IGF-2 and GHR gene of Odontobutis potamophila
1.6 數據分析
實驗結果數據采用2-ΔΔCt法來計算目的基因的相對表達量,應用SPSS 19軟件對實驗數據進行統計分析、差異顯著性檢驗分析。用t檢驗計算P值,當P<0.05時為差異顯著,P<0.01為差異極顯著。數值均采用平均值±標準差(Means±SD)表示。使用Origin 8.6軟件繪制柱狀圖。
2 結果與分析
2.1 GHR和IGF-2基因的序列分析
利用RACE技術克隆得到河川沙塘鱧GHR基因的cDNA全長序列為2 719 bp(GenBank登錄號:MG820031)(圖1),包括編碼609個氨基酸的開放閱讀框1 830 bp、251 bp的5′UTR區和638 bp的3′UTR區,其中3′UTR區域具有28 bp的PolyA尾巴,表明獲得的該cDNA全長序列3′末端序列具有完整性。ExPASy軟件預測的氨基酸分子量為98 589.94,理論等電點為4.8,GHR多肽序列包含一個21個氨基酸的信號肽、227個氨基酸的胞外結構域、22個氨基酸的單個跨膜結構域和339個氨基酸的胞內結構域,在胞外結構中包含有保守的半胱氨酸殘基和FGDFSmotif,胞內有兩個保守的box(圖1-A)。
IGF-2基因的cDNA全長序列為1 586 bp(GenBank登錄號:MG820032)(圖 2),包括 642 bp的開放閱讀框、48 bp的5′UTR區和896 bp的3′UTR區,其中3′UTR區域具有16 bp的 PolyA尾巴,表明獲得的該cDNA全長序列3′末端序列也具有完整性。共編碼213個氨基酸,ExPASy軟件預測的氨基酸分子量為24 448.46,理論等電點為10.0。IGF-2編碼區經SignalP軟件分析發現,其包含一個52個氨基酸的信號肽及A、B、C、D和E 5個保守的結構域,且6個半胱氨酸殘基參與形成3對二硫鍵(圖1-B)。
2.2 GHR和IGF-2基因的進化樹及氨基酸同源性分析
采用MEGA 5.0以N-J法構建基于GHR和IGF-2氨基酸序列的分子系統進化樹(圖2)。進化樹聚類結果顯示,所有魚類聚為一支,兩棲類(及雞)和哺乳類動物聚類為另外的分支,而河川沙塘鱧IGF-2、GHR基因與其親緣關系較近的大黃 魚 (Larimichthys crocea)、大 彈 涂 魚(Bolephthalmus pectinirostris)等魚類聚為一支,這與傳統的分類系統是一致的(圖2)。同源性序列分析表明河川沙塘鱧GHR基因與大彈涂魚、石斑魚(Epinephelus coioides)、半滑舌鰨(Cynoglossus semilaevis)、大黃魚的同源性為68.95%~80.79%(圖 3)。IGF-2基因與大黃魚、尖吻鱸(Lates niloticus)、鱖 (Siniperca chualsi)、卵 形 鯧 鲹(Trachinotus ovatus)的同源性較高,為86.98%~88.84%(圖4)。
2.3 河川沙塘鱧GHR和IGF-2基因的表達特征分析
利用qRT-PCR的方法分析了河川沙塘鱧GHR、IGF-2基因在9種組織表達情況,由圖5可知,GHR和IGF-2在9種組織中均有表達且在肝中的表達量最高(P<0.05);除肝臟之外,GHR在腦、肌肉、性腺和心臟中的表達量均顯著高于其他4種組織;而IGF-2在肌肉中的表達量相對較高,在性腺和腦中的表達量其次,在其他組織的表達量相對較低(圖5)。
分析河川沙塘鱧GHR、IGF-2基因在胚胎到仔魚不同發育時期的表達模式,由圖6可知,GHR基因在胚胎發育的各個時期均有表達,表達量呈先上升后下降的趨勢,在原腸胚時期達到最大值,極顯著的高于其他發育階段(P<0.01)。仔魚階段表達量迅速升高,其表達量是受精卵階段的4倍。相較于GHR基因,IGF-2在胚胎及仔魚時期表達量一直呈升高趨勢,與GHR基因相似,IGF-2基因在胚胎時期同樣在原腸胚時期表達量顯著高于其他時期(P<0.05)。
2.4 GHR、IGF-2基因的雌雄表達差異
如圖7所示,雄魚GHR和IGF-2基因在肝中的表達量顯著高于雌魚,GHR基因表達呈上升趨勢,IGF-2則先下降再上升。而在腦和肌肉中僅IGF-2基因在210 d的腦和150、210 d的肌肉中具有顯著性雌雄差異,其他時期無顯著性差異。
3 討論
魚類的生長與哺乳動物相類似,受到GHR/IGF軸的調控。GH主要在垂體產生,一方面可與肝臟等靶器官GHR作用,產生IGFs,通過IGFs調節動物生長發育;另一方面可直接與肌肉等靶器官上GHR結合,通過JAKSTAT途徑啟動信號轉導直接影響細胞營養物質分泌及代謝[3]。本研究首次克隆了河川沙塘鱧IGF-2和GHR基因的全長序列,序列分析表明河川沙塘鱧IGF-2和GHR基因的氨基酸序列與其他脊椎動物的相應基因表現出高度的同一性和相似性,表明這些基因在魚類和哺乳動物中可能具有相似的功能。

圖1 河川沙塘鱧GHR(A)和IGF-2(B)cDNA序列及推導的氨基酸序列Fig.1 Nucleotide and deduced am ino acid sequences of O.potamophila GHR(A)and IGF-2(B)cDNA

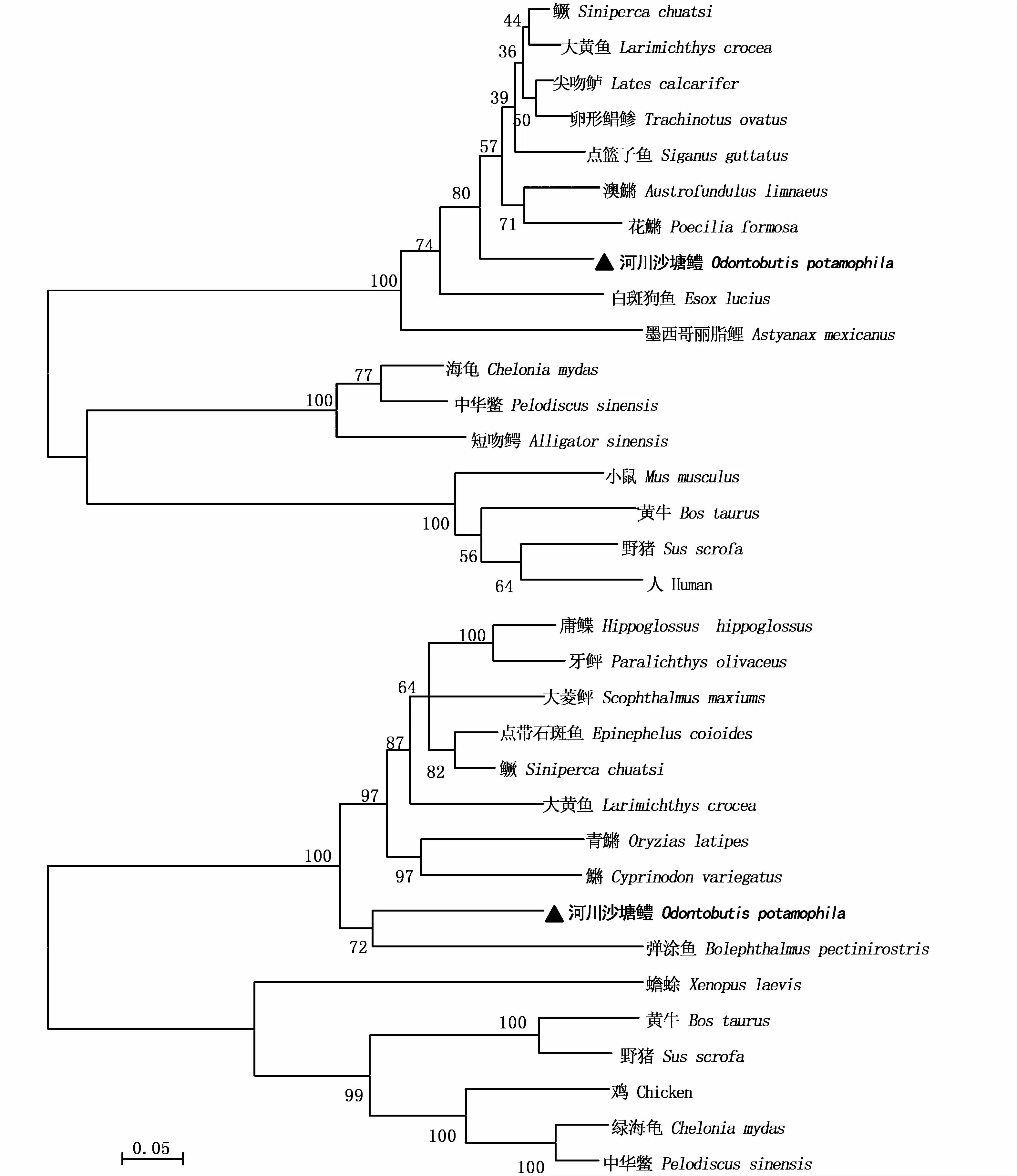
圖2 河川沙塘鱧與其他物種的GHR和IGF-2基因系統進化樹Fig.2 Phylogenetic tree of GHR and IGF-2 gene of O.potamophila and other vertebrates

圖3 河川沙塘鱧GHR氨基酸序列與其他物種同源序列對比Fig.3 M ultiple alignment of O.potamophila GHR polypeptide sequences w ith other known orthologs

圖4 河川沙塘鱧IGF-2氨基酸序列與其他物種同源序列對比Fig.4 Multiple alignment of O.potamophila IGF-2 polypeptide sequences w ith other known orthologs
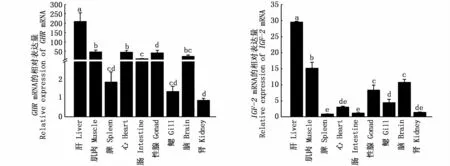
圖5 GHR和IGF-2在河川沙塘鱧不同組織中的表達Fig.5 GHR and IGF-2 gene expressions in various tissues of O.potamophila
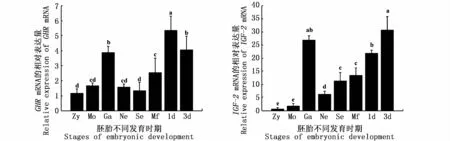
圖6 GHR和IGF-2在河川沙塘鱧胚胎和仔魚發育過程中的表達Fig.6 GHR and IGF-2 gene expressions during the embryonic and larval development of O.potamophila
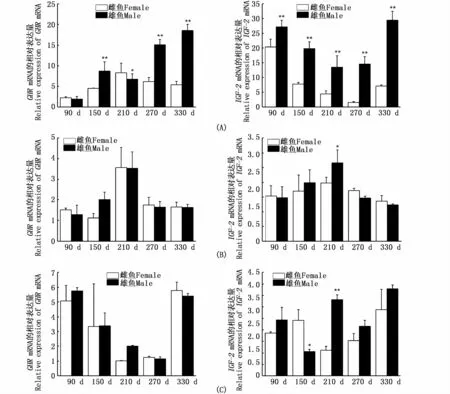
圖7 河川沙塘鱧不同生長階段GHR和IGF-2在肝(A)、腦(B)、肌肉(C)中表達量的雌雄對比Fig.7 Expression differences of GHR and IGF-2 in O.potamophila liver(A),brain(B)and muscle(C)between themale and fem ale
和大多數硬骨魚的GHR一樣,河川沙塘鱧GHR的胞外區有7個保守的半胱氨酸殘基和1個FGEFSmotif,其中半胱氨酸殘基可形成3個內部二硫鍵,未配對的半胱氨酸可能在GH誘導的GH受體二聚化過程中形成二硫鍵[19]。此外,FGDFSmotif靠近跨膜區,其在GH與GHR結合過程中起穩定空間構象的作用,能保證配體與受體的正常結合以及進一步的信號轉導過程,并參與GHR的二聚化[3],這是已知魚類GHR的共性之一。GHR胞內含有特征結構高度保守的BOX1、BOX2基序,BOX1區域是JAK2結合的位點,而BOX2區域參與受體的增殖反應,推測這兩個區域在河川沙塘鱧GH所介導的信號轉導過程中發揮重要作用。與哺乳動物相比BOX1中的保守序列PPVPVP在河川沙塘鱧的GHR基因中為PPIPAP,這種微小的改變也在其他魚中被發現[20-21],這種改變可能是GHR基因適應不同物種信號轉導機制的結果。自從PALAMARCHU等[15]在大麻哈魚中首次克隆出魚類的IGF-2基因,許多學者開始了對其生物學的研究。IGF-2前體蛋白由N端至 C端的信號肽、B、C、A、D、E區構成,切除信號肽和 E區導致轉變為成熟肽[22]。在本實驗中,河川沙塘鱧IGF-2基因全長1 586 bp,編碼212個氨基酸,成熟肽為71個氨基酸。系統進化樹分析表明,河川沙塘鱧GHR先與大彈涂魚(Boleophthalmus pectinirostris)聚為一支,再與青鳉(Oryzias latipes)、雜色鳉(Cyprinodon variegatus)聚為一支。而IGF-2則自己聚為一支,后與亞馬遜花鳉(Poecilia formosa)等聚為一支。聚類分析產生的進化樹基本上與各參考物種的分類進化地位相吻合,由此可以在一定程度上推測河川沙塘鱧的系統進化地位。
許多硬骨魚類GHR和IGF-2基因都具有廣泛表達的特性,如在暗紋東方鲀(Takifugu obscurus)、半滑舌鰨(Cynoglossus semilaevis)、金頭雕 (Sparus aurata)、多 耙 牙 鲆 (Paralichthys adspersus)中[20,23-24],在本 研 究 中 河 川 沙塘 鱧GHR基因在9個組織中均有表達,這與之前的研究是相一致的。GHR在多種組織中都有組織表達的模式支持了魚類GHR具有多種生理功能的觀點。TSE等[26]對鯽的研究表明IGF-2廣泛分布于腦、鰓、心臟、頭腎、肝臟、脾臟、腸,其中在肝臟中的表達量最高,這與已研究物種莫桑比克羅非魚(Oreochromismossambicus)的IGF-2在肝中的表達量最高相一致[16]。在本實驗中,也發現河川沙塘鱧GHR、IGF-2基因在檢測的9個組織中肝臟的表達量最高,表明GHR、IGF-2主要在肝中產生;其次,肝臟是GHR與GH結合和信號轉導最重要的組織,也是魚類和其他哺乳類IGFs轉錄的主要位點,這也可能是GHR、IGF-2基因在肝臟中高豐度表達的原因。魚類未受精卵中存在來自于母源保存和傳遞的 IGF-1和IGF-2 mRNA,對于進行體外受精和發育的硬骨魚類來說,這些母源傳遞的mRNA對于胚胎發育具有重要作用[27]。在很多硬骨魚 類中如 虹鱒[28]、斑馬魚[29]、金頭鯛[27]、半滑舌鰨[23]中 IGF-2和 GHR基因在胚胎發育過程中均持續表達。EIVERS等[30]在斑馬魚的研究中發現 IGF-2和 IGF-1 mRNA在胚胎發育的原腸期存在表達,表明IGF-2對早期胚體發育模式的建立具有重要作用。WHITE[31]在斑馬魚的研究中發現GHR在原腸胚期表達量最高,作為介導GH通路的重要組件,在胚胎發育中發揮重要作用。然而在IGF-2基因敲除實驗中,IGF-2 mRNA在囊胚期及原腸期高表達,而敲除IGF-2基因后并未對原腸期胚胎發育產生影響,提示IGF-2 mRNA盡管在原腸期高表達,但其翻譯成多肽實際發揮作用的時間可能要在原腸期之后[30]。河川沙塘鱧IGF-2和GHR基因在胚胎發育過程中均有表達,且在囊胚到原腸胚期表達量相對較高,這表明IGF-2和GHR基因在胚胎發育過程中扮演重要角色。
動物的生長軸基因在控制機體生長、發育、代謝的過程中發揮著不可替代的作用,其生長軸基因的表達存在雌雄二態性,是導致雌雄生長速度具有顯著差異性的原因之一,在魚類中這種兩性生長差異更加明顯,例如黃顙魚(Pseudobagrus vachelli)雄性個體生長明顯快于雌性個體,而鱖(Siniperca chuatsi)雌魚生長速度快于其雄魚[32-33]。河川沙塘鱧在生長方面也具有明顯的性別二態性,雄性河川沙塘鱧比雌性同期生長速度快30%以上。針對這一問題本研究檢測了河川沙塘鱧生長相關基因GHR和IGF-2在雌、雄魚不同生長階段的相關組織腦、肝、肌肉中的表達情況,結果表明在河川沙塘鱧快速生長期雄魚肝中GHR、IGF-2基因表達量均顯著高于雌魚,這與黃顙魚的研究結果一致[32],說明河川沙塘鱧的生長與GHR、IGF-2的調控作用是密切相關的。國內外大量研究表明,GHR可以通過GH/IGF生長內分泌軸促進動物組織細胞的分化與生長,例如MA等[23]對比了半滑舌鰨不同發育時期雌、雄魚肝臟中GHR基因的表達變化,發現不同發育階段在雄魚中的表達量均高于雌魚。大麻哈魚[24]、斑點叉尾鮰[34]、尼羅羅非魚[17]禁食后肝 GHR mRNA表達量均下降,降低了GH對魚的促生長作用,使魚類的生長變慢。在鯉(Cyprinus carpio)中注射GH后,腦、胰臟和肝中IGFs的表達量均顯著上升[26]。CHEN等[17]對尼羅羅非魚的研究表明,每周注射IGF1和IGF2,到第5周時尼羅羅非魚體長和體質量都明顯升高。在河川沙塘鱧中GHR基因在不同發育時期肝組織中的表達量有很大的差異,IGF-2在肝中的表達量也存在明顯的雌、雄二態性,說明IGFs在肝臟中的高表達與肝臟在產生IGFs和發揮IGFs生理功能方面起關鍵作用是一致的。GHR和IGF-2基因在河川沙塘鱧腦和肌肉中的大多數發育階段沒有顯著性差異,因此筆者推測肝中GHR和IGF-2基因表達的雌、雄差異可能是導致河川沙塘鱧生長性別二態性的主要原因。本文為研究河川沙塘鱧生長提供了分子生物學依據,也為進一步研究魚類生長基因的調控機制奠定了一定的基礎。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
