中國不同地區草地貪夜蛾種群生物型分子特征分析
2019-09-04 09:34:39張磊柳貝姜玉英劉杰吳孔明蕭玉濤
植物保護 2019年4期
張磊 柳貝 姜玉英 劉杰 吳孔明 蕭玉濤
摘要 原產于美洲的草地貪夜蛾已入侵我國,對我國糧食生產安全構成重大威脅。在美洲,該蟲的寄主類型、遷飛行為以及抗藥性等方面因群體分化而存在較大差異。本研究利用2個分子標記對中國13省(市、自治區)131個縣市的318份樣品進行群體遺傳特征比較,基于線粒體COⅠ基因分析結果顯示96%以上為水稻型,玉米型比例不到4%,且中國樣品COⅠ序列特征與美國佛羅里達州種群有很高的一致性,而基于核基因組Tpi基因分析結果表明所有樣品單倍型特點均表現為玉米型。分析認為,入侵我國的草地貪夜蛾群體很可能來自一個水稻型母本和玉米型父本雜交群體的后代,在長期的演化擴散過程中,玉米型的核基因組占據了主導地位,從而成為一種特殊的玉米型。厘清入侵中國的草地貪夜蛾具體的遺傳特征屬性,對于風險評估、精準監測和科學防控具有重要指導意義。
關鍵詞 草地貪夜蛾; 群體演化; 水稻型; 玉米型; 雜交
中圖分類號: S 435.132
文獻標識碼: A
DOI: 10.16688/j.zwbh.2019296
聯合國糧農組織全球預警的重大害蟲草地貪夜蛾自2019年1月入侵中國以來,5個月時間內已擴散至18個省(市、自治區),大有席卷全國之勢,給我國農業生產造成重大影響,國家糧食安全和生態安全受到嚴重威脅。
目前,草地貪夜蛾在我國主要為害玉米和甘蔗[1],對其他作物的潛在危害造成的損失尚未確定。考慮到草地貪夜蛾在美洲的為害特征,其寄主植物多達80余種,對玉米、水稻、高粱、大豆、小麥、棉花、牧草等均造成較大的經濟損失[23],其危害的潛在風險巨大。草地貪夜蛾還存在寄主種類嗜好性差異,主要由于存在兩種不同的亞型,通常稱為水稻型和玉米型[45],除此之外,遷飛行為[67]、農藥抗性[89]以及對轉基因作物的抗性[1011]也與不同地理群體相關。目前我國發生草地貪夜蛾的省份都是水稻種植區,水稻是否會受到為害引起高度關注。因此,對入侵我國的草地貪夜蛾進行群體遺傳特征鑒定顯得極為迫切。
分子標記是鑒定草地貪夜蛾不同亞型的主要手段,其中來源于線粒體的細胞色素C氧化酶亞基Ⅰ基因(cytochrome c oxidase subunit Ⅰ,COⅠ)以及位于Z染色體上的磷酸甘油醛異構酶基因(triose-phosphate isomerase,Tpi),是當前最常用也是最成熟的標記基因[1221]。不同亞型之間的雜交行為在實驗室以及田間均證實存在[2223],由于線粒體具有母系遺傳的特征,基于線粒體COⅠ基因的亞型鑒定具有一定的不確定性,而基于核基因組Z染色體的Tpi基因則表現出相對的穩定性[2425]。然而,關于草地貪夜蛾復雜的群體遺傳動態分析,這兩個標記依然是當前最合適的選擇。
本研究采集我國13個省(市、自治區)131個縣(市、區)的318份草地貪夜蛾樣品,利用COⅠ和Tpi兩個基因片段進行分子檢測,鑒定具體生物型,并分析種群遺傳特征。
1 材料與方法
1.1 試驗材料
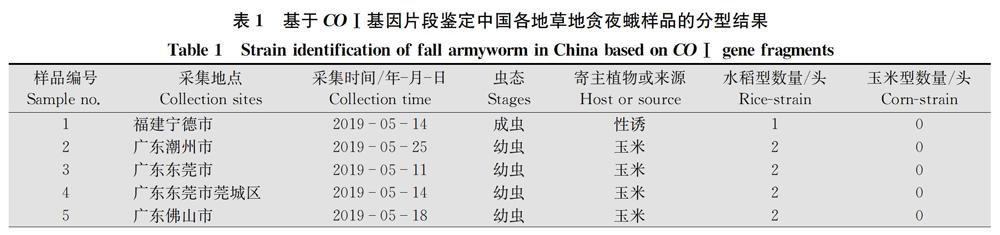
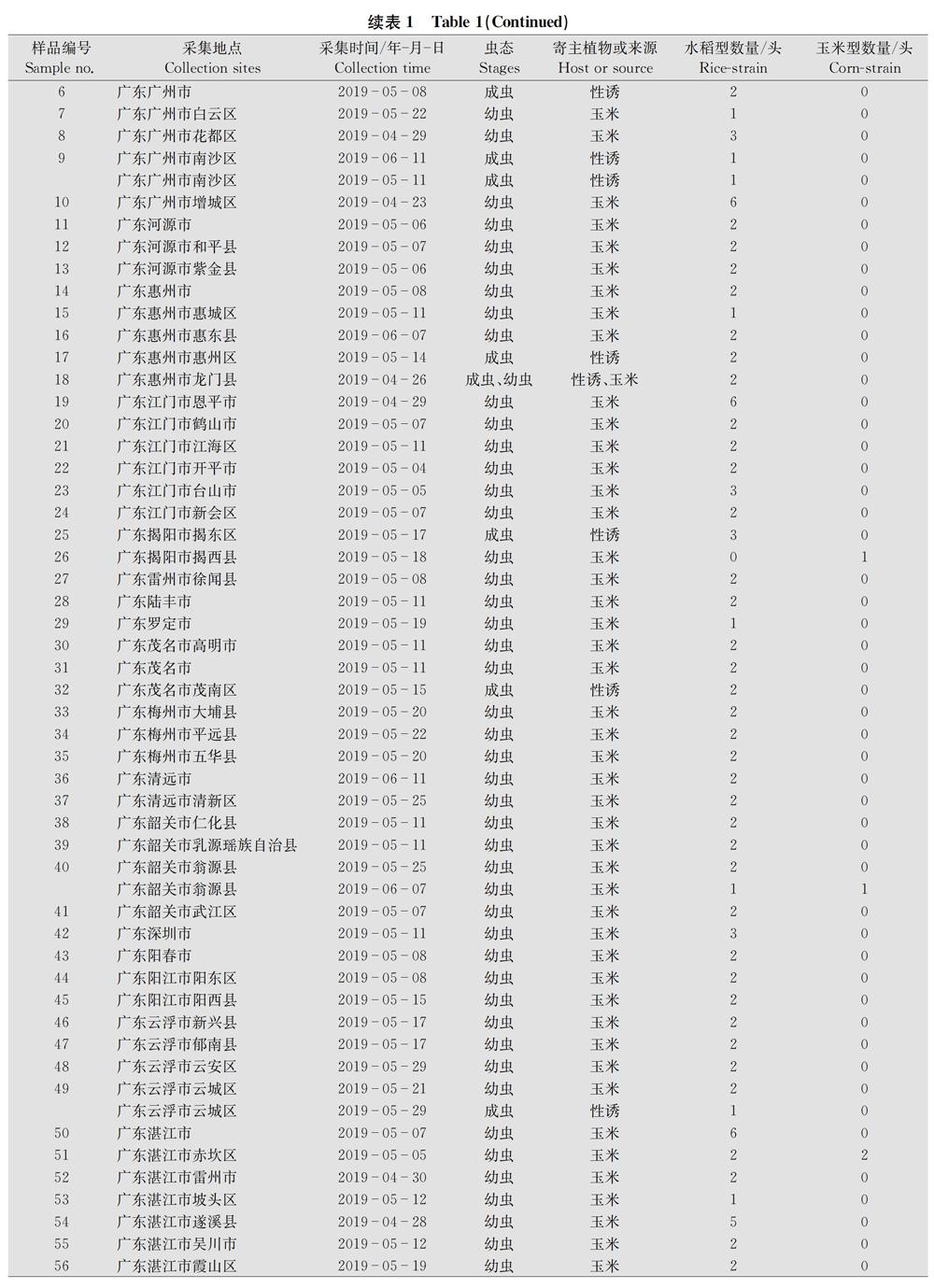
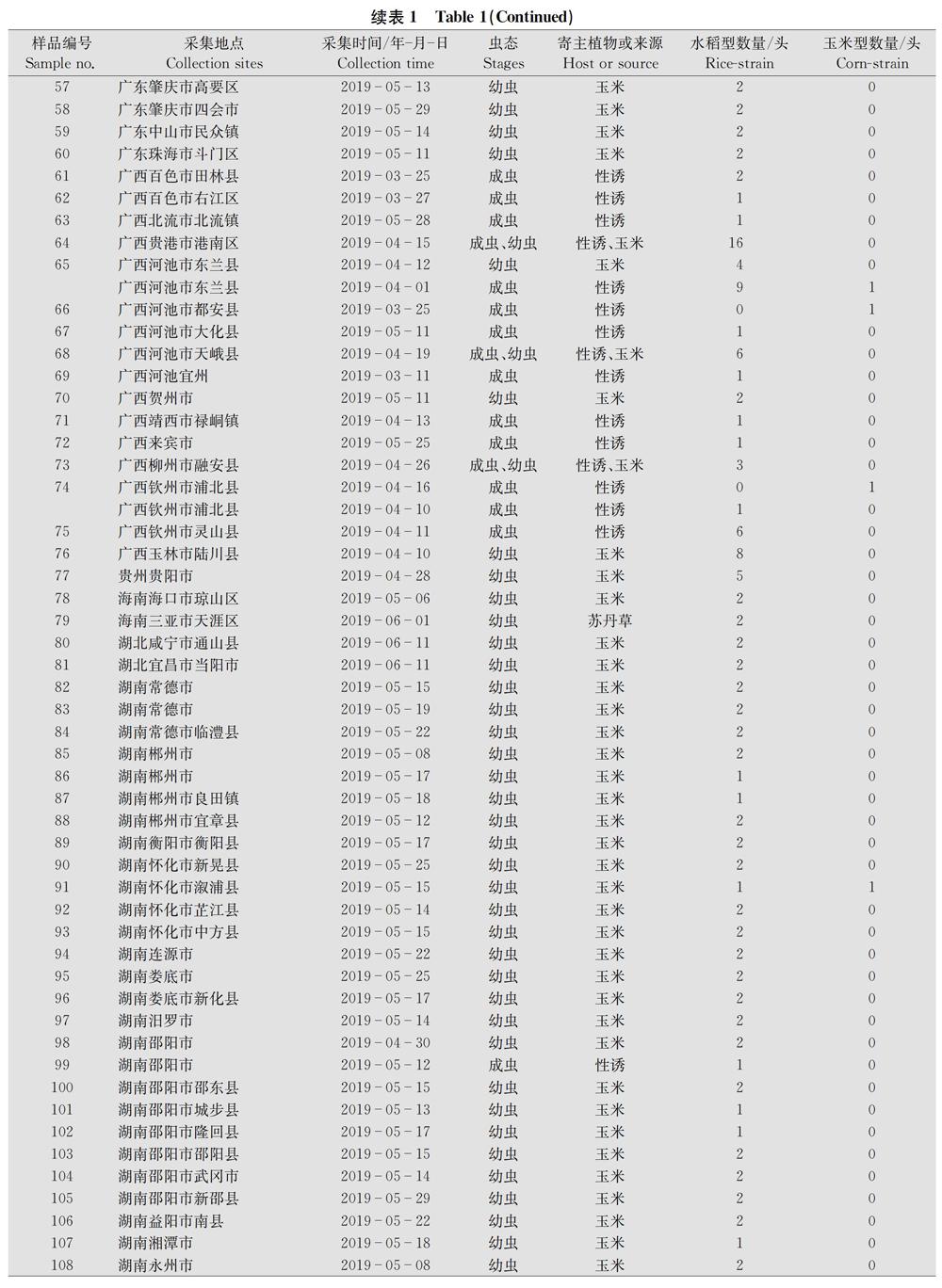
本次研究的樣品一共有318份,包含田間采集的幼蟲和性誘的成蟲。采集地點包括云南、廣西、廣東、四川、貴州、海南、湖南、福建、浙江、江西、上海、重慶、湖北13個省(市、自治區)131個縣(市、區),樣品采集時間為2019年3月11日至6月11日,寄主植物主要為玉米,少數樣品取于蘇丹草或水稻。幼蟲樣品收到后用75%乙醇浸泡30 min,隨后常溫運送至實驗室,成蟲樣品保持干燥,常溫運送至實驗室。各縣市具體樣品信息見表1。
1.2 PCR擴增及測序
成蟲或幼蟲單頭蟲體用液氮研磨成粉后,采用Multisource Genomic DNA Miniprep Kit試劑盒進行DNA提取,DNA經過1%瓊脂糖電泳檢測,使用NanoDrop ND-2000儀器測定濃度。根據已發表引物和PCR擴增方法[26],分別對COⅠ基因和Tpi基因片段進行擴增,使用1%瓊脂糖電泳檢測擴增結果,PCR產物送生工生物工程(上海)股份有限公司測序。
1.3 數據分析
利用BioEdit軟件查看測序序列峰圖,去除兩端低質量序列,區分雜合位點,獲得準確的基因片段序列。從GenBank數據庫下載全球各代表性地區草地貪夜蛾COⅠ基因核酸序列。所有序列采用MEGA 5.0軟件進行比對,刪除兩端不對齊序列,選取Kimura-2參數模型,以鄰接法(neighbor joining,NJ)構建系統進化樹,系統樹各分支的自舉檢驗值(bootstrap)由1 000次重復檢驗獲得。針對玉米型和水稻型Tpi基因序列單倍型差異特點[25],運用DNAMAN軟件進行特異位點比對分析。
2 結果與分析
通過對COⅠ基因序列進行比對分析,從318份樣品中鑒定出水稻型和玉米型兩種基因型,其中水稻型306份,占96.2%,玉米型不到4%(表1)。可見以COⅠ基因作為分子標記,水稻型在群體數量上占絕對主導。
將本研究測定的COⅠ基因序列與NCBI公布的北美美國、南美巴西、非洲南非、亞洲印度等的草地貪夜蛾的COⅠ序列進行比對,以棉鈴蟲Helicoverpa armigera COⅠ序列作為外群構建系統進化樹(圖1)。由于我國所有樣品中屬于同一基因型的樣品序列高度一致,我們僅各選取1條序列(strain 1和strain 2)進行比對分析,結果表明,中國樣品strain 2的COⅠ序列與印度、南非、美國佛羅里達種群的序列一致,與巴西以及美國的另一種群的序列有一定差別。雖然整個分支均為水稻型,但是從進化樹也可看出,美洲不同地域種群的序列之間存在一定程度變異。在玉米型分支中,中國樣品strain 1與南非以及美國佛羅里達種群的序列一致。與水稻型分支類似,來自美國不同地區的玉米型的COⅠ序列也表現出一定的差異。
我們比對所有樣品Tpi基因序列,針對水稻型和玉米型10個差異單倍型位點進行分析,除了部分樣品在2個位點顯示為雜合外(圖2),其余樣品均只顯示一種基因型,并且均與玉米型單倍型特征相符合,該結果表明基于Tpi基因的序列分析顯示所有樣品均為玉米型。
3 討論
草地貪夜蛾在原產地美洲由于取食寄主植物的不同分化出玉米型和水稻型兩種亞型,玉米型偏向取食玉米、甘蔗、高粱、谷子、棉花、大豆、花生等,而水稻型偏向取食水稻、各種牧草等。兩種亞型在形態學上完全無法區分[5, 27],通過COⅠ和Tpi等分子標記進行鑒定一直是主流方法,能夠根據不同亞型單倍型特點將絕大多數水稻型和玉米型有效區分[1214, 2425]。此外,不同亞型在長期進化過程中,形成了一些特定的群體,包括對農藥的抗性群體以及針對轉基因作物的抗性群體,這些群體的鑒定也多依賴于分子輔助檢測手段[9, 11]。
在前期工作中,我們利用COⅠ和Tpi兩種分子標記聯合檢測,發現入侵我國云南的82份草地貪夜蛾幼蟲樣品具有完全一致的基因型,并且通過Tpi基因單倍型位點比對分析鑒定為玉米型[26]。在本研究中,我們擴大了樣品采集地區,大大增加了樣本數量,不僅包含了各齡期幼蟲和成蟲,還覆蓋了不同類型的寄主植物,運用相同的方法對入侵我國的草地貪夜蛾群體遺傳特征進行了更進一步的研究。分析結果表明入侵我國的草地貪夜蛾具有兩種COⅠ基因型,而基于Tpi基因的結果顯示均為玉米型,僅僅部分樣品在2個位點表現出雜合現象,使用兩種分子標記鑒定出現了不一致的結果。由于線粒體屬于母系遺傳,后代僅能遺傳母本的線粒體DNA,且兩種亞型之間并不存在絕對的生殖隔離,雌性水稻型和雄性玉米型的交配行為在實驗室和田間均被證實存在[23],因此分析認為,本研究中的不一致是由于水稻型母本和玉米型父本雜交產生,導致后代獲得了水稻型線粒體的COⅠ基因序列,但是核基因組在長期的繁衍擴散過程中更多地保留了玉米型的背景基因組,從而演化成特殊的玉米型。本研究中占主導的96%的樣品為一個雜交型群體的后代,由水稻型母本和玉米型父本雜交后代演化而來。由于在我國所有樣品中均未檢測出完全符合水稻型Tpi基因單倍型,因此推測該雜交發生在入侵我國、甚至是入侵非洲之前。
通過構建基于COⅠ基因的系統進化樹,我們發現入侵我國的草地貪夜蛾與南非、印度以及美國佛羅里達的草地貪夜蛾聚為一支,序列比對也表明具有完全一樣的基因型。在美洲,根據COⅠ基因單倍型頻率可以將玉米型草地貪夜蛾分為佛羅里達(FL-type)和德克薩斯(TX-type)兩個地理型亞群,其中TX-type廣泛分布在南美、墨西哥和美國大部分地區,FL-type限于佛羅里達、加勒比海地區和美國東海岸[2830]。已有研究報道顯示,入侵非洲的草地貪夜蛾群體很可能是通過貿易往來被帶入非洲的一小部分群體繁衍擴散形成的[2]。由于在美洲已有少量關于雜交型群體后代的報道[22],因此極有可能是來自美國佛羅里達的雜交演化群體后代傳入非洲。從進化樹可以看出,中國草地貪夜蛾與非洲草地貪夜蛾COⅠ基因序列完全一致,且主導型比例相同[15,31],因此我們推斷草地貪夜蛾入侵非洲之后,不斷擴散,隨后進入西亞,持續往東擴散至東南亞地區,最后通過中緬邊境侵入我國,呈現擴散暴發態勢。
雖然目前有部分研究報道水稻型在非洲和亞洲也存在,但是僅僅是基于COⅠ單基因或多個線粒體來源的基因鑒定[15,17],對于Tpi基因的分析也是根據個別變異位點的判斷[16],基于PCR的一代測序也可能會由于擴增或測序誤差造成堿基差異。在本研究中,部分樣品Tpi基因也存在2個位點的雜合(圖2),由于草地貪夜蛾性別決定屬于ZW型,Tpi基因位于Z染色體,因此可能會因為雌性(ZW)或者雄性(ZZ)個體性別差異造成雜合。
通常情況下,背景單一的小種群長期的近交會經歷種群瓶頸效應,容易造成遺傳衰退[32]。而侵入我國的草地貪夜蛾的祖先具有雜交背景,后代理論上可能具有某種程度的雜交優勢,顯示出更強的適應性;也可能由于雜交而產生一定的適合度代價,導致自身抵抗能力下降。值得注意的是,侵入我國的草地貪夜蛾在取食行為以及為害特征上可能與傳統的玉米型有所差異,包括農藥抗性和對轉基因作物的抗性遺傳特征會相應改變,因此僅僅通過連鎖分子標記檢測可能造成一定偏差,還需要結合更加完整的生物測定進行判斷。對于這一新的玉米型草地貪夜蛾,在防控措施上可能要更加的謹慎。鑒于入侵群體祖先經歷過雜交,理論上存在進化出水稻型的可能性,還需要進一步基因組測序的數據來論證,因此需要對水稻作物進行更加嚴密的監控。
目前我國發現受害作物主要有玉米,部分省份有甘蔗、高粱和少量的谷子、花生,均屬于玉米型寄主植物。四川省合江縣植保站4月30日在水稻秧苗捕捉到幾頭高齡草地貪夜蛾幼蟲(COⅠ基因鑒定為草地貪夜蛾,但是Tpi基因無法擴增),湖南農業大學李有志教授5月13日在湖南省岳陽市平江縣稻田發現在3 m2的范圍內有17頭4齡期幼蟲,并且都在稻株上進行取食,后來當地植保部門普查未在稻田見有為害。推測為低齡幼蟲吐絲擴散至水稻植株上,或遷入成蟲在無法搜尋到最適寄主玉米的情況下,被迫選擇水稻產卵,因此應是環境脅迫造成的偶然事件。
總的來說,目前我國發生的草地貪夜蛾存在兩種線粒體COⅠ基因型,且比例差異懸殊。然而兩種基因型所代表的群體之間是否有雜交,在雌雄交配選擇上是否有偏好性,需要更進一步的研究。此外,本研究中有多份樣品僅能得到COⅠ擴增片段,而采用Tpi引物無法擴增出有效片段,可能是由于樣品DNA質量較差,也不排除具有更加復雜的基因組背景特征,需要采用全基因組分析手段,選取更多的位點,針對水稻型和玉米型兩種亞型基因組差異特征[33],進行具體的測序分析和位點比對,更加深入地挖掘入侵我國的草地貪夜蛾的遺傳特征,為其防控提供更加準確的信息。我國當前對于草地貪夜蛾的群體遺傳研究有限,亟待更多的數據支持。還應密切跟蹤非洲、亞洲等國家水稻型擴散動向,提高我國防范該類型草地貪夜蛾的主動性。
參考文獻
[1] 楊普云,朱曉明,郭井菲,等.我國草地貪夜蛾的防控對策與建議[J/OL].植物保護,2019,45.https:∥doi.org/10.16688/j.zwbh.2019260.
[2] NAGOSHI R N,GOERGEN G,PLESSIS H D,et al. Genetic comparisons of fall armyworm populations from 11 countries spanning sub-Saharan Africa provide insights into strain composition and migratory behaviors[J/OL]. Scientific Reports,2019,9:8311.
[3] STOKSTAD E. New crop pest takes Africa at lightning speed[J]. Science,2017,356 (6337):473474.
[4] PASHLEY D P,MARTIN J A. Reproductive incompatibility between host strains of the fall armyworm (Lepidoptera:Noctuidae)[J]. Annals of the Entomological Society of America,1987,80(6):731733.
[5] PASHLEY D P. Host-associated genetic differentiation in fall army worm (Lepidoptera:Noctuidae):a sibling species complex[J]. Annals of the Entomological Society of America,1986,79(6):898904.
[6] MITCHELL E R,MCNEIL J N,WESTBROOK J K,et al. Seasonal periodicity of fall armyworm,(Lepidoptera:Noctuidae) in the Caribbean basin and northward to Canada[J]. Journal of Entomological Science,1991,26(1):3950.
[7] WESTBROOK J K,NAGOSHI R N,MEAGHER R L,et al. Modeling seasonal migration of fall armyworm moths [J]. International Journal of Biometeorology,2016,60:255267.
[8] YU S J,NGUYEN S N,ABO-ELGHAR G E. Biochemical characteristics of insecticide resistance in the fall armyworm,Spodoptera frugiperda (J. E. Smith)[J]. Pesticide Biochemistry and Physiology,2003,77:111.
[9] CARVALHO R A,OMOTO C,FIELD L M,et al. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda[J/OL]. PLoS ONE,2013,8(4):e62268.
[10]STORER N P,BABCOCK J M,SCHLENZ M,et al. Discovery and characterization of field resistance to Bt maize:Spodoptera frugiperda (Lepidoptera:Noctuidae) in Puerto Rico [J]. Journal of Economic Entomology,2010,103(4):10311038.
[11]BANERJEE R,HASLER J,MEAGHER R,et al. Mechanism and DNA-based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda)[J/OL]. Scientific Reports,2017,7:10877.
[12]MEAGHER R L, CALLO-MEAGHER M. Identifying host of fall armyworm (Lepidoptera:Noctuidae) in Florida using mitochondrial markers[J]. Florida Entomologist, 2003, 86(4): 450455.
[13]NAGOSHI R N,MURUA M G,HAY-ROE M,et al. Genetic characterization of fall armyworm (Lepidoptera:Noctuidae) host strains in Argentina [J]. Journal of Economic Entomology,2012,105(2):418428.
[14]NAGOSHI R N. The fall armyworm triosephosphate isomerase (Tpi) gene as a marker of strain identity and interstrain mating [J]. Annals of the Entomological Society of America,2010,103(2):283292.
[15]JACOBS A,VUUREN A V,RONG I H. Characterisation of the fall armyworm (Spodoptera frugiperda J. E. Smith) (Lepidoptera:Noctuidae) from South Africa [J]. Africa Entomology,2018,26(1):4549.
[16]NAGOSHI R N,KOFFI D,AGBOKA K,et al. Comparative molecular analyses of invasive fall armyworm in Togo reveal strong similarities to populations from the eastern United States and the Greater Antilles [J/OL]. PLoS ONE,2017,12(7):e0181982.
[17]KALLESHWARASWAMY C M,ASOKAN R,MAHADEVASWAMY H M,et al. First record of invasive fall armyworm,Spodoptera frugiperda (J. E. Smith) (Lepidoptera:Noctuidae) on rice (Oryza sativa) from India[J]. Journal of Entomology and Zoology Studies,2019,7(3):332337.
[18]JUREZ M L,MURUA M G,GARCA M G,et al. Host association of Spodoptera frugiperda (Lepidoptera:Noctuidae) corn and rice strains in Argentina,Brazil and Paraguay[J]. Journal of Economic Entomology,2012,105(2):573582.
[19]OTIM M H,TEK T W,WALSH T K,et al. Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera:Noctuidae) from Uganda [J/OL]. PLoS ONE,2018,13(4):e0194571.
[20]COCK M J W,BESEH P K,BUDDIE A G,et al. Molecular methods to detect Spodoptera frugiperda in Ghana,and implications for monitoring the spread of invasive species in developing countries [J/OL]. Scientific Reports,2017,7:4103.
[21]NAGOSHI R N,GOERGEN G,TOUNOU K A,et al. Analysis of strain distribution,migratory potential,and invasion history of fall armyworm populations in northern Sub-Saharan Africa [J]. Scientific Reports,2018,8:3710.
[22]NAGOSHI R N,MEAGHER R L,NUSSELY G,et al. Effects of fall armyworm (Lepidoptera:Noctuidae) interstrain mating in wild populations [J]. Environmental Entomology,2006,35:561568.
[23]PASCALINE D,FABRICE L,CLAIRE L,et al. Spodoptera frugiperda (Lepidoptera:Noctuidae) host-plant variants:two host strains or two distinct species?[J]. Genetica,2015,143(3):305316.
[24]NAGOSHI R N,MEAGHER R L. Using intron sequence comparisons in the triose-phosphate isomerase gene to study the divergence of the fall armyworm host strains [J]. Insect Molecular Biology,2016,25(3):324337.
[25]NAGOSHI R N. Improvements in the identification of strains facilitate population studies of fall armyworm subgroups[J]. Annals of the Entomological Society of America,2012,105(2):351358.
[26]張磊,靳明輝,張丹丹,等. 入侵云南草地貪夜蛾的分子鑒定[J].植物保護,2019,45(2):1924.
[27]PASHLEY D P,HAMMOND A M,HARDY T N. Reproductive isolating mechanisms in fall armyworm host strains (Lepidoptera,Noctuidae)[J]. Annals of the Entomological Society of America,1992,85(4):400405.
[28]NAGOSHI R N,SILVIE P,MEAGHER R L. Comparison of haplotype frequencies differentiate fall armyworm (Lepidoptera:Noctuidae) corn-strain populations from Florida and Brazil [J]. Journal of Economic Entomology,2007,100(3):954961.
[29]NAGOSHI R N,MEAGHER R L,FLANDERS K,et al. Using haplotypes to monitor the migration of fall armyworm (Lepidoptera:Noctuidae) corn-strain populations from Texas and Florida [J]. Journal of Economic Entomology,2008,101(3):742749.
[30]NAGOSHI R N,ROSAS-GARCIA N M,MEAGHER R L,et al. Haplotype profile comparisons between Spodoptera frugiperda (Lepidoptera:Noctuidae) populations from Mexico with those from Puerto Rico,South America,and the United States and their implications to migratory behavior [J]. Journal of Economic Entomology,2015,108(1):135144.
[31]NAGOSHI R N. Evidence that a major subpopulation of fall armyworm found in the Western Hemisphere is rare or absent in Africa,which may limit the range of crops at risk of infestation [J/OL]. PLoS ONE,2019,14(4):e0208966.
[32]FACON B,HUFBAUER R,TAYEH A,et al. Inbreeding depression is purged in the invasive insect Harmonia axyridis[J]. Current Biology,2011,21(5):424427.
[33]GOUIN A,BRETAUDEAU A,NAM K. Two genomes of highly polyphagous lepidopteran pests (Spodoptera frugiperda,Noctuidae) with different host-plant ranges [J/OL]. Scientific Reports,2017,7:11816.
(責任編輯:楊明麗)
