康菲溢油事故 艱難的法律維權(quán)路
2012-09-09 08:11:12李鈺
中國(guó)新時(shí)代 2012年1期
關(guān)鍵詞:法律
| 文· 本刊實(shí)習(xí)記者 李鈺
2012年新年伊始,雖然仍處寒冬,但大家都憧憬著美好的未來(lái),期待春暖花開(kāi)的好日子。然而,康菲溢油事故中的受害者卻絲毫感覺(jué)不到新年的喜氣,內(nèi)心猶如這冬天一樣冰冷。雖然各方面表明海洋污染訴訟的歷程將困難重重,但是仍然有很多人拿起法律利劍之器奔走在維權(quán)路上。一路遇到的坎坷,讓我們不禁深思,誰(shuí)能為這次事故的維權(quán)之路點(diǎn)上一盞照明燈呢?我們期待曙光的出現(xiàn)
康菲溢油何時(shí)止?
2011年6月4日,康菲公司在渤海灣蓬萊19-3油田B平臺(tái)附近的海床出現(xiàn)原油滲漏;6月17日,C平臺(tái)一口在鉆井發(fā)生小型井涌,導(dǎo)致溢油發(fā)生。蓬萊19-3油田是我國(guó)最大的海上油氣田,屬于康菲公司與中海油的合作項(xiàng)目,康菲公司為作業(yè)方。7月5日,即溢油發(fā)生一個(gè)多月之后,中國(guó)國(guó)家海洋局才發(fā)布事故公告,證實(shí)康菲溢油事故的發(fā)生,并介入調(diào)查。但是在這一個(gè)月期間,康菲公司并未向公眾透露半點(diǎn)關(guān)于海洋石油污染的信息,直到70多天后,才首次向公眾披露油基泥漿總量由原來(lái)預(yù)計(jì)的1,500桶增加到2,500桶,遠(yuǎn)超預(yù)期。
2011年8月,國(guó)家海洋局對(duì)康菲公司下達(dá)最后期限,要求在8月31日前徹底排查并切斷溢油源,徹底排查并消除再次發(fā)生溢油的風(fēng)險(xiǎn)。8月19日,康菲中國(guó)承諾做好渤海灣安全、環(huán)保和清理工作,首次向公眾道歉,但對(duì)賠償只字未提。8月24日,康菲公司召開(kāi)新聞發(fā)布會(huì),總裁司徒瑞介紹康菲公司已經(jīng)對(duì)溢油源進(jìn)行了永久性封堵,同時(shí)表示愿意就此事件承擔(dān)應(yīng)負(fù)的責(zé)任。而當(dāng)有人問(wèn)及英國(guó)BP公司在墨西哥灣溢油事件中設(shè)立200億美元賠償基金時(shí),康菲公司總裁的答案是:康菲不是BP。8月31日,在眾所關(guān)注的“大限”之日,康菲向國(guó)家海洋局提交了一份綜合報(bào)告,向海洋局及外界確認(rèn)“康菲已經(jīng)查明并且封堵了溢油源”,完成了“兩個(gè)徹底”。但是,蓬萊19—3油田溢油事故聯(lián)合調(diào)查組做出結(jié)論:康菲石油中國(guó)有限公司沒(méi)有完成“兩個(gè)徹底”,因?yàn)?月2日C平臺(tái)附近的海面上有大片油帶,個(gè)別地點(diǎn)還有油花冒出,海面溢油污染情況沒(méi)有明顯改善。B平臺(tái)即使沒(méi)有發(fā)現(xiàn)油帶和漂油,但是據(jù)中國(guó)海監(jiān)北海總隊(duì)副總隊(duì)長(zhǎng)林芳忠向媒體介紹,即使康菲公司安裝了集油罩,但是集油罩是一個(gè)臨時(shí)性措施,仍存在著溢油污染海洋的風(fēng)險(xiǎn)。
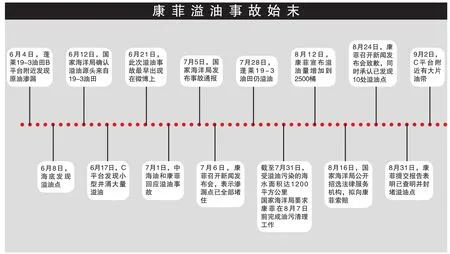
康菲石油中國(guó)在2011年9月6日宣布將設(shè)立基金,旨在根據(jù)中國(guó)相關(guān)法律承擔(dān)公司應(yīng)盡的責(zé)任并有益于渤海灣的整體環(huán)境。總部位于休斯頓的康菲公司又于9月19日宣布設(shè)立第二項(xiàng)基金。然而,自9月6日公布第一項(xiàng)渤海灣基金至今,上述兩項(xiàng)基金的資金規(guī)模、法律依據(jù),以及出資方、受益方等詳情,至今成謎。國(guó)家海洋局副局長(zhǎng)王飛在國(guó)家海洋局8月25日召開(kāi)的渤海溢油事故處置情況視頻通報(bào)會(huì)上介紹,渤海溢油事故已經(jīng)造成累計(jì)5,500平方公里海面遭受污染,其中“劣四類”水質(zhì)海面超過(guò)870平方公里,已有部分渤海周邊岸線受到溢油事故影響;2011年11月,事故調(diào)查組組長(zhǎng)告訴媒體,溢油面積已達(dá)到了6,200平方公里。本刊記者從國(guó)家海洋局網(wǎng)站獲悉,2011年12月5日,雖然B、C平臺(tái)附近海域未見(jiàn)油帶,但是C平臺(tái)附近有油花溢出,最少0個(gè)/分鐘,最多15個(gè)/分鐘,估算當(dāng)日溢油量約0.16升。曾有媒體將康菲溢油事故與墨西哥灣溢油事故做過(guò)比較:“雖然漏油總量?jī)H有其1/10左右,但渤海的面積只有墨西哥灣的1/20,而且封閉程度要比墨西哥灣大得多,計(jì)算出的油污染負(fù)荷量要比墨西哥灣漏油高出1倍多,所造成污染強(qiáng)度將遠(yuǎn)超過(guò)墨西哥灣漏油事故。”

2011年11月11日,由國(guó)家海洋局牽頭國(guó)土資源部、環(huán)境保護(hù)部、交通運(yùn)輸部、農(nóng)業(yè)部、安監(jiān)總局、國(guó)家能源局等部門組成的聯(lián)合調(diào)查組在康菲溢油事故責(zé)任認(rèn)定報(bào)告中指出,康菲石油中國(guó)有限公司在蓬萊19-3油田生產(chǎn)作業(yè)過(guò)程中沒(méi)有執(zhí)行相關(guān)方案,事故定性為“重大海洋溢油污染責(zé)任事故”。
海洋污染訴訟之回顧
2010年4月20日,英國(guó)石油公司(BP)在墨西哥灣的鉆井平臺(tái)爆炸,約2億桶原油泄漏入海,釀成美國(guó)史上最嚴(yán)重的漏油事故。事發(fā)后,受到影響的美國(guó)公民和企業(yè)陸續(xù)對(duì)BP提出了300多件民事訴訟,要求經(jīng)濟(jì)賠償。6月1日,美國(guó)司法部宣布對(duì)這家跨國(guó)石油公司展開(kāi)民事和刑事雙重調(diào)查。在多方壓力下,BP公司設(shè)立200億美元賠償基金,為漏油事故受害者提供賠償。據(jù)路透社消息,BP公司通過(guò)旗下設(shè)立的基金向墨西哥灣漏油事故中20.44萬(wàn)名受害者支付了超過(guò)50億美元的賠償金。但是,對(duì)于BP的追責(zé)與賠償沒(méi)有停止。經(jīng)過(guò)半年多的調(diào)查取證,美國(guó)司法部向美國(guó)聯(lián)邦法庭提出起訴,認(rèn)為BP等9家企業(yè)無(wú)視安全生產(chǎn)法律、違規(guī)操作最終釀成悲劇。根據(jù)美國(guó)《石油污染法》,9家被告企業(yè)應(yīng)為原油泄漏事故所造成的全部清理成本和損失承擔(dān)責(zé)任,其中包括給美國(guó)自然資源帶來(lái)的破壞,索賠金額不設(shè)上限;此外,依據(jù)美國(guó)《潔凈水資源法》,BP應(yīng)承擔(dān)每桶泄漏原油1,100美元的民事罰款,如被判嚴(yán)重違規(guī)操作,罰款金額將高達(dá)每桶4,300美元。而據(jù)美國(guó)政府8月發(fā)布的報(bào)告,在墨西哥灣漏油事故中泄漏原油的總量為490萬(wàn)桶。這意味著B(niǎo)P面臨著54億至210億美元的民事罰款。雖然至今尚未對(duì)墨西哥灣溢油索賠訴訟作出判決,但是污染者BP公司、美國(guó)司法部等主體在事發(fā)后的積極態(tài)度以及美國(guó)相關(guān)法律依據(jù)的完善值得中國(guó)借鑒和思考。
康菲溢油引起的海洋污染并非我國(guó)海洋污染案例的首例,早在之前,我國(guó)就已經(jīng)發(fā)生了幾起比較嚴(yán)重的海洋污染,就在人們漸漸淡忘生態(tài)損失維權(quán)之艱難的時(shí)候,海洋污染因這起溢油事故再次出現(xiàn)在公眾的視線中。2002年“塔斯曼海”溢油案、2006年渤海海域石油污染案、2010年“7·16”大連原油爆炸泄漏事故案等,早已將中國(guó)海洋生態(tài)損害的評(píng)估、鑒定、環(huán)境監(jiān)測(cè)等制度缺失問(wèn)題暴露于外,中國(guó)環(huán)境保護(hù)尤其是海洋污染法律適用的不明、訴訟取證的不易、國(guó)家對(duì)大型國(guó)企的“特殊關(guān)照”等種種跡象表明:受傷的總是普通百姓,放棄訴訟等于吞食污染者留下的慢性毒藥,而法律維權(quán)得來(lái)的卻是身心疲憊和對(duì)法律的失望與無(wú)奈。我國(guó)極力倡導(dǎo)依法治國(guó),然而,環(huán)境保護(hù)法治化何時(shí)能夠真正實(shí)現(xiàn)?這是所有關(guān)心生態(tài)環(huán)境保護(hù)、因環(huán)境污染受損的廣大人士共同關(guān)心的話題。
中國(guó)海洋污染損失的法律維權(quán)并未就此結(jié)束。2011年6月的康菲溢油事故啟動(dòng)了我國(guó)又一次海洋污染索賠歷程。即使已有了多次的維權(quán)經(jīng)歷,但中國(guó)環(huán)境保護(hù)法律法規(guī)不健全、監(jiān)管手段和技術(shù)落后、司法易受外界因素干擾等多重因素使得各方人士對(duì)康菲的索賠之路前景不太看好。
康菲溢油索賠之路怎么走?
據(jù)國(guó)家海洋局公告,北海分局將代表國(guó)家依據(jù)相關(guān)法律的規(guī)定,向溢油事故責(zé)任方提起海洋生態(tài)損害索賠訴訟。據(jù)悉,已向全社會(huì)公開(kāi)選聘了法律服務(wù)團(tuán)隊(duì)代理此案。國(guó)家海洋局北海分局有關(guān)人士在接受《每日經(jīng)濟(jì)新聞》采訪時(shí)透露,相關(guān)律師已完成80%以上的取證工作,希望能在2011國(guó)慶節(jié)之前正式向康菲石油中國(guó)提起海洋生態(tài)索賠訴訟。但是至今未聽(tīng)到國(guó)家海洋局正式起訴康菲石油中國(guó)的消息。據(jù)中國(guó)政法大學(xué)教授王燦發(fā)分析,至今遲遲不起訴可能是源于國(guó)家海洋局對(duì)此次訴訟的慎重態(tài)度,海洋污染訴訟具有復(fù)雜性,必須準(zhǔn)備充分的證據(jù)方可保證案件的勝訴。
王燦發(fā)介紹,在康菲溢油事故發(fā)生后,污染受害者法律幫助中心專門指派律師赴河北省唐山市樂(lè)亭縣調(diào)查水產(chǎn)養(yǎng)殖場(chǎng)受污染情況,為漁民提供索賠及固定證據(jù)等方面的專業(yè)指導(dǎo),積極關(guān)注康菲溢油事故的后續(xù)索賠訴訟并對(duì)法律援助工作持續(xù)跟進(jìn)。目前,法律援助中心與受損害的漁民正在聯(lián)系之中。經(jīng)了解,幫助中心在十多年間已經(jīng)幫助了220多起環(huán)境案件的受害者向法院提起訴訟,有三分之一的案件取得了勝訴,有的即使敗訴,但是相關(guān)的工廠也被叫停。法律幫助中心的無(wú)償法律援助為受污染者維權(quán)帶來(lái)了福音。
北京華城律師事務(wù)所律師賈方義走在康菲溢油事故訴訟維權(quán)的最前端。2011年8月初,賈律師以個(gè)人名義向青島海事法院、天津海事法院以及海南省高級(jí)人民法院提起訴訟,要求中海油和康菲石油設(shè)立100億元的賠償基金,進(jìn)行生態(tài)賠償和生態(tài)恢復(fù)。賈律師向本刊記者轉(zhuǎn)述了法院的答復(fù),“公益訴訟屬于新型案件,須上報(bào)請(qǐng)示”,因此這場(chǎng)被稱為“標(biāo)桿”性質(zhì)的公益訴訟在法院立案門檻前止步。賈律師表示,“渤海是中國(guó)13億人的財(cái)產(chǎn),海洋污染侵害了中國(guó)的每一個(gè)人”,因此他提起的訴訟是有依據(jù)的,但是法院并未表態(tài)。賈律師表示,他已于2011年11月30日向全國(guó)人大提交立法修訂意見(jiàn),希望將公民納入公益訴訟原告范圍中。他認(rèn)為“《民事訴訟法》修正案將公民排除在公益訴訟原告之中,不利于環(huán)境保護(hù)權(quán)益的維護(hù)。”
賈律師還向北京市第一中級(jí)人民法院提出訴訟,狀告國(guó)家海洋局行政不作為。然而,這一訴訟也未立案。賈律師說(shuō),一中院曾向他轉(zhuǎn)述了國(guó)家海洋局領(lǐng)導(dǎo)的意見(jiàn),即事故原因需要調(diào)查,因此沒(méi)有及時(shí)公布事故信息,希望“延緩訴訟”。賈律師不認(rèn)可這一答復(fù),因?yàn)椤把泳徳V訟并非法律術(shù)語(yǔ)”。
除外,賈律師表示,他已于2011年9月6日向山東省公安廳、山東省檢察院、國(guó)家海洋局遞交公開(kāi)信,舉報(bào)康菲石油中國(guó)有限公司有重大環(huán)境污染事故罪等嫌疑,要求公安機(jī)關(guān)依據(jù)中國(guó)相關(guān)法律對(duì)其予以刑事調(diào)查。
賈律師在公益訴訟和行政訴訟均遭遇碰壁之后,接受了山東牟平30位漁民的訴訟代理,毅然走上了民事?lián)p害賠償訴訟之路。對(duì)于這次訴訟,賈律師原本信心滿滿,認(rèn)為在實(shí)體法和程序法中均不存在困難。賈律師表示,環(huán)境污染損害賠償訴訟實(shí)行“舉證責(zé)任倒置”,只要原告一方舉證證明死亡的魚蝦、貝類表面有油污顆粒和損失的大小即可。因此在賈律師乘著小船、迎著風(fēng)浪完成取證后,于11月18日向青島海事法院遞交起訴書,請(qǐng)求判令兩被告連帶承擔(dān)溢油事故環(huán)境污染給原告造成經(jīng)濟(jì)損失的賠償責(zé)任合計(jì)約715萬(wàn)元以及判令兩被告承擔(dān)訴訟費(fèi)用。賈律師告訴記者,他曾于23日打電話給青島海事法院確認(rèn)起訴書已收到。賈律師和漁民開(kāi)始了焦急的等待。然而,《民事訴訟法》規(guī)定的7天立案期限結(jié)束,他們也沒(méi)有得到立案的消息。本刊記者在2011年11月30日17點(diǎn)30分電話采訪了賈律師,電話中傳來(lái)的是焦急、憤慨的聲音。賈律師表示,他在立案的最后一天打電話給青島海事法院詢問(wèn)立案結(jié)果,法院將是否立案的決定權(quán)推諉給法院研究室,讓其咨詢研究室,然而研究室電話一天均無(wú)人接聽(tīng),立案最后期限日遭遇法院集體失語(yǔ)。
路在何方?
有媒體稱賈律師的行為是中國(guó)海洋侵權(quán)訴訟的“標(biāo)桿”。此刻,賈律師表示他已經(jīng)茫然了,不知這支“標(biāo)桿”該插在哪里、該如何擺正。賈律師說(shuō),在遭遇法院集體失語(yǔ)后,他作為一個(gè)法律工作者再次感受到了中國(guó)法律的困惑,貌似聽(tīng)到了漁民集體的哭泣。當(dāng)記者問(wèn)到接下來(lái)有什么打算時(shí),賈律師表示,“已有美國(guó)環(huán)境訴訟律師與我聯(lián)系,表示若中國(guó)不受理的話可以去美國(guó)提起訴訟,那么下一站估計(jì)要在康菲總部所在地美國(guó)德克薩斯州起訴了。”賈律師于2011年11月30在博客中寫下了《新渤海漁歌》:七日立案法無(wú)影,何處療治民心傷?默默無(wú)語(yǔ)千行淚,耳邊陣陣漏油聲。
去美國(guó)起訴不失為索賠的一條出路,但是,涉外民事訴訟帶來(lái)的訴訟成本的增加由誰(shuí)來(lái)承擔(dān)呢?已經(jīng)損失慘重的漁民如何負(fù)擔(dān)得了這些?所以,在不放棄涉外訴訟這條路的同時(shí),最大的希望是中國(guó)環(huán)境保護(hù)法律的健全,保障維權(quán)者維權(quán)之路通暢,這才是保護(hù)污染受害者權(quán)益的最根本需求。
猜你喜歡
新少年(2023年9期)2023-10-14 15:57:47
法律方法(2022年1期)2022-07-21 09:17:10
法律方法(2021年3期)2021-03-16 05:57:02
法律方法(2019年4期)2019-11-16 01:07:16
法律方法(2019年3期)2019-09-11 06:27:06
法律方法(2019年1期)2019-05-21 01:03:26
法律方法(2018年2期)2018-07-13 03:21:38
學(xué)生天地(2016年23期)2016-05-17 05:47:10
山東青年(2016年1期)2016-02-28 14:25:30
中國(guó)衛(wèi)生(2015年1期)2015-11-16 01:05:56
