CRISPR/Cas9技術編輯淀粉合成基因PUL
2019-10-18 03:20:58奉寶兵魏祥進焦桂愛胡時開鄔亞文謝黎虹圣忠華邵高能唐紹清
中國稻米 2019年5期
奉寶兵 魏祥進 焦桂愛 胡時開 鄔亞文 謝黎虹 圣忠華 邵高能 唐紹清
(中國水稻研究所/農業部水稻生物學與遺傳育種重點實驗室/水稻生物學國家重點實驗室,杭州310006;第一作者:tuopuyigou@163.com;*通訊作者:sqtang@126.com)
胚乳中淀粉的生物合成是通過一系列酶促反應轉變成淀粉的過程,因此ADP-葡萄糖焦磷酸化酶(AGPase,ADP glucose pyrophosphorylase)、淀粉合成酶(SS,starch synthase)、淀粉分支酶(SBE,starch branching enzyme)和淀粉去分支酶(DBE,debranching enzyme)被稱為淀粉合成過程中的4類關鍵酶[1-4]。
DBE家族分為異淀粉酶亞家族ISA1、ISA2、ISA3和PUL普魯蘭酶。DBEs在淀粉的合成與降解中都具有重要作用[5-7]。在支鏈淀粉合成過程中,可以移除不合理分子的短鏈,并且將其連接到新鏈上,保證淀粉形成穩定的晶體結構;在種子萌發的過程中,可以特異性切割支鏈淀粉的α-1,6糖苷鍵,保證淀粉降解,促進種子發芽[8-9]。
前人對DBEs的異淀粉酶(isoamylase)研究報道很多,但對OsPUL的研究較少。FUJITA等[10]認為,OsPUL可以部分恢復isa1癟粒突變體的表型,并報道不同突變位點與千粒重(脫殼后)既有正相關也有負相關。FUJITA等鑒定了3種不同pul等位突變體e10、i16和EM1003。對千粒重分析發現,EM1003(啟動子上游1 266 bp處C替換了T)千粒重為22.42±0.29 g,比野生型(千粒重為 20.37±0.03 g)增加 10.06%;而e10(第 10個外顯子內突變)和i16(第16個內含子內突變)千粒重分別為 20.12±0.02 g和 19.00±0.03 g,與野生型相比略有下降。LI等[11]報道OsPUL是胚乳特異性表達基因,在開花后15 d相對表達量最高,認為在OsPUL的啟動子區存在GCN4、AACA和E2F等多個順式作用元件。
近幾年基因組編輯(Genome editing)育種迅速發展[12-13],其主要優勢是不依賴隨機重組或雜交過程、可以精準誘變、易獲得純合突變。基因組編輯技術主要有三種人工核酸酶,分別是鋅指核酸酶(ZFNs)、TALE核酸酶(TALENs)以及CRISPR系統[14]。中科院遺傳所高彩霞課題用優化的Cas9蛋白對水稻OsBADH2、OsPDS和OsMPK2單個基因進行定點編輯,之后科研人員定點編輯了OsBadh2、OsGn1a、OsGT3、OsTGW6等重要的產量和品質相關基因[15-16]。
本研究用CRISPR/Cas9技術對調控稻米脫支酶基因OsPUL進行定點編輯,進一步探索OsPUL對淀粉合成和產量的關系,期望獲得具有重要育種價值的PUL等位變異,并為分子育種提供材料基礎和理論指導。
1 材料與方法
1.1 試驗材料
本研究所用水稻材料為粳稻品種中花11(Zhonghua 11,ZH11),以及以ZH11為轉基因受體的T0代植株、T0代植株自交后代T1植株等。所有轉基因材料單本移栽到轉基因安全圃,中耕除草,水肥管理和病蟲草害防治同大田管理。
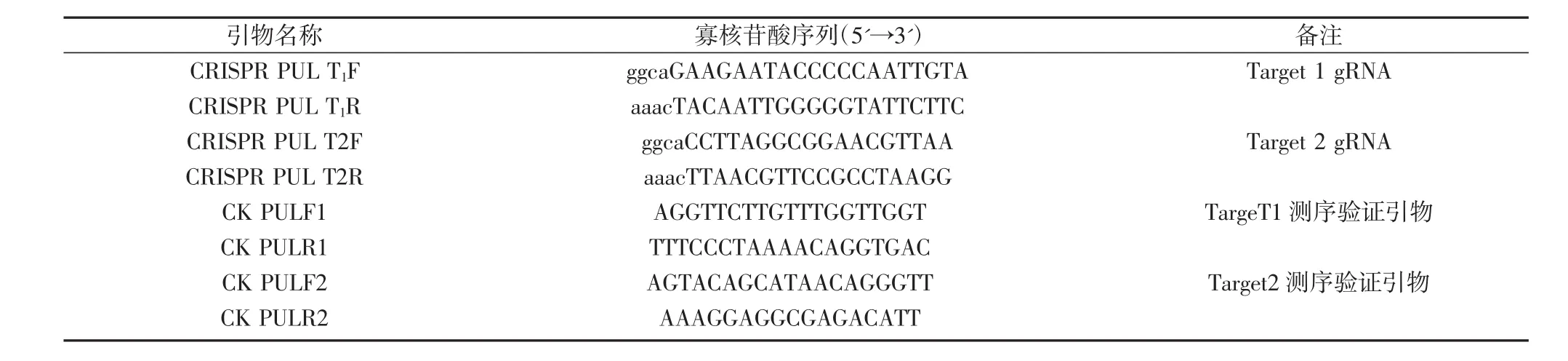
表1 CRISPR所用引物
1.2 PUL編輯載體構建
根據CRISPR/Cas9的引物設計原理[17-18],設計2個靶序列CRISPR-PUL-1和CRISPR-PUL-2(表1)。靶序列的引物退火成雙鏈后,與線性化的SK-gRNA/AarI(20~50 ng)連接,構建成中間載體gRNA::PUL1和gRNA::PUL2。然后,用一步法將 gRNA::PUL1、gRNA::PUL2和pC1300-Cas9等3個載體同時線性化,利用同尾酶具有相同的粘性末端的特性,將三者連接成pC1300-Cas9::PUL1-PUL2。CRISPR/Cas9相關載體SK-gRNA和pC1300-Cas9等由中國水稻研究所王克劍課題組饋贈。
1.3 轉基因陽性植株鑒定
將構建好的雙靶點pC1300-Cas9::PUL1-PUL2載體通過農桿菌EHA105介導轉化水稻品種ZH11成熟胚愈傷組織,T0代轉基因植株經過煉苗后單本移栽轉基因安全圃。分蘗期取葉片并編號,提取轉基因水稻基因組DNA。用潮霉素引物HptF/R(產物750 bp)和Cas9蛋白引物Cas9-Protein F/R(產物500bp)進行PCR擴增。
同時,根據靶位點序列特征設計CAPS標記,對PCR擴增產物用內切酶酶切鑒定(PCR-RE),能被切開成兩條帶,說明靶位點序列沒有發生突變,反之,則發生突變。此外,以ZH11和轉基因植株基因組DNA為模板,CK-PUL-F1/R1和CK-PUL-F2/R2為引物分別擴增PUL基因的2個靶位點,PCR產物測序結果用SnapGene 2.32軟件進行序列比對和峰圖分析,鑒定轉基因植株中PUL的基因型。
1.4 農藝性狀考察
在田間選擇同一小區里長勢一致,剔除邊行的植株,測定T1部分突變體的株高、穗長、劍葉長、劍葉寬,并計算這些植株的分蘗數、穗粒數、結實率。收種后烘干種子,去除癟粒,枝梗;隨機挑選飽滿的種子稱量千粒重,并用萬深考種儀考察粒長、粒寬、長寬比等粒形性狀。
1.5 稻米生理生化品質
對T0和T1兩代種子進行稻米品質以下生理指標檢測:總淀粉含量、直鏈淀粉含量、蛋白質含量、糊化溫度等,每個樣品測3次,取平均值進行統計分析。總淀粉含量測定采用分光光度計法,直鏈淀粉含量用Megazyme試劑盒(K-AMYL07/II)測定;蛋白質含量用近紅外透射光譜技術測定,實驗步驟參照孫成效的方法[3]。糊化溫度用差示掃描儀(DSC)測定,參照 Kweon M[19]的方法。
1.6 粒重和淀粉合成相關基因表達分析
取開花后12 d的種子,立即放入液氮中,然后轉入-80℃冰箱長期保存。選5粒冷凍種子進行RNA提取,參照Prescott和Marti的方法。用賽默飛公司的Nanodrop 2000檢測總RNA的純度和濃度,RNA質量合格后進行反轉錄。在Roche Light Cycle 480 Real-Time PCR儀上,以Ubiquitin作內參,用SYBR qPCR Mix配置反應體系,反應程序為:95℃ 30 s,95℃ 5 s/60℃ 10 s/72℃ 10 s(40 個循環),68℃ 10 min,10℃ 8 min。利用儀器自帶軟件進行分析,并用2-△△CT方法計算各基因的相對表達量和制作直方圖。
2 結果與分析
2.1 PUL基因編輯陽性植株鑒定
為了驗證PUL基因是否被成功敲除,我們首先利用CAPS引物對獲得的45株T0代轉基因植株進行PCR擴增及酶切檢測。從圖1可知,Psp1406I的酶切結果顯示,轉基因植株中靶位點2處有6株不能被限制酶切開,即發生突變,分別為 T0-3、T0-18、T0-21、T0-22、T0-32和T0-39。MunI酶切結果顯示,轉基因植株中靶位點1處有19株不能被限制酶切開,產生突變,分別為 T0-1、T0-3、T0-4、T0-7、T0-8、T0-11、T0-12、T0-15、T0-18、T0-21、T0-22、T0-25、T0-27、T0-32、T0-33、T0-36、T0-40、T0-43 和 T0-45(圖 2)。綜上所述,2 個靶位點處共有20株發生了基因編輯事件。

圖1 Psp1406I酶切靶位點2

圖2 MunI酶切驗證靶位點1

圖3 Sanger測序結果
PCR產物酶切結果可大致判斷靶位點處是否發生突變,卻不能準確判斷是純合突變還是雜合突變,以及突變類型。因此,我們對靶序列處的500 bp左右PCR產物進行測序鑒定突變類型。結果表明,T0代轉基因植株中靶位點1處有11株野生型、34株突變體,突變率為75.5%;靶位點2處有21株野生型、24株突變體,突變率為53.3%。我們對上述轉基因材料于成熟期單株收種,并對T0-1、T0-6、T0-11和T0-18共4個轉基因系進行遺傳分析。從圖3可知部分T1單株的突變類型主要以單堿基的插入和單堿基的缺失為主,并造成移碼突變導致氨基酸序列改變,從而影響蛋白質功能。
由考種數據可知T0-18的粒重極顯著增加(圖4 F),于是重點對T1-18的轉基因系進行遺傳分析,結果表明,敲除載體在T1-18后代中發生了分離(圖5)。此外,通過對T1-18的24個單株PUL 2個靶位點進行測序分析,發現目的基因的突變位點也發生了分離(表2)。并獲得了純合單株,2個靶位點都是純合突變用mu表示;2個靶位點存在1個雜合突變用H表示。因此,我們通過CRISPR/Cas9基因編輯技術對水稻淀粉脫支酶家族的普魯蘭基因進行編輯,獲得了pul的純合突變體,同時還獲得剔除載體骨架的遺傳材料2個,分別為T1-18-10和T1-18-21(圖5和表2)。
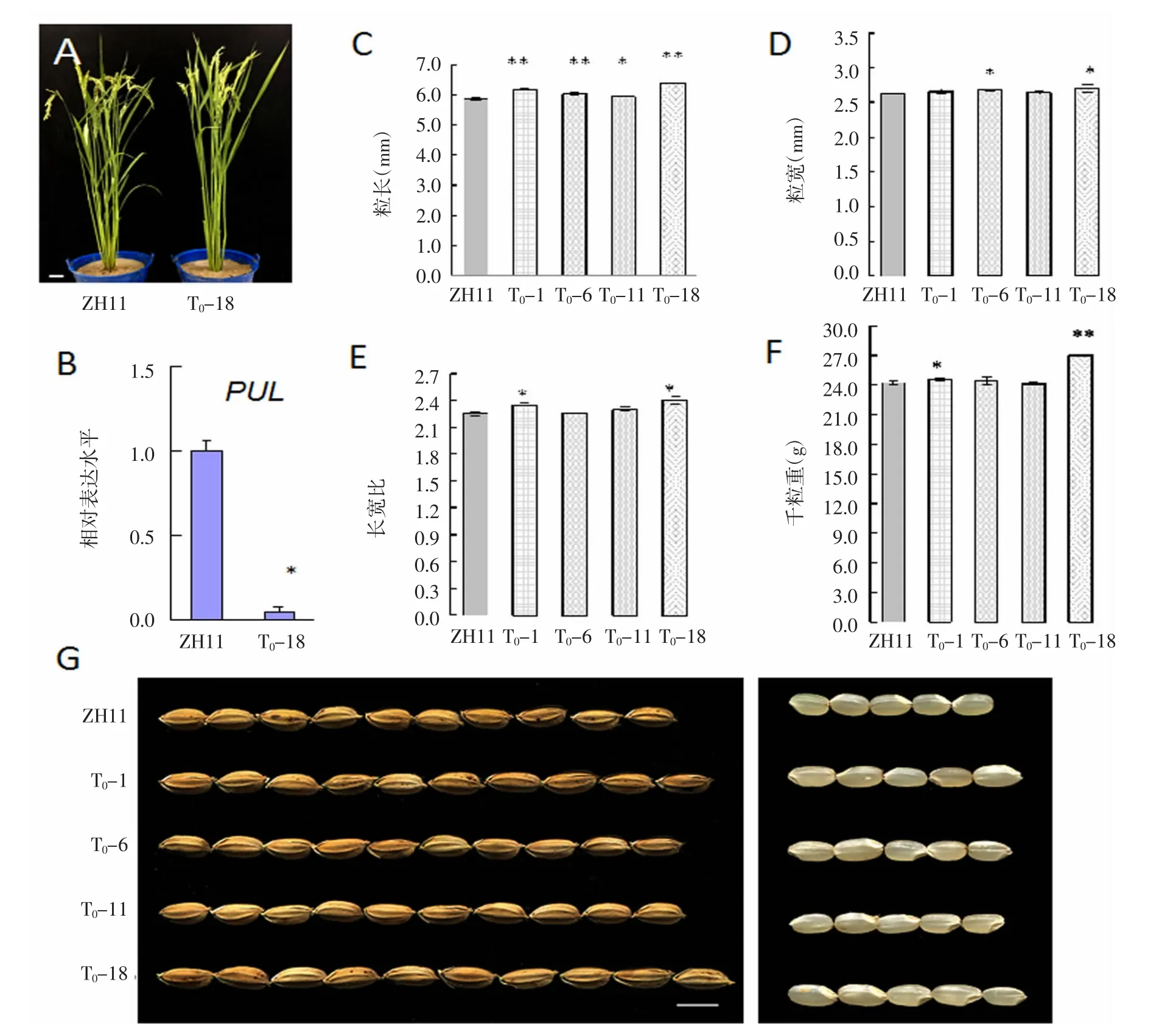
圖4 相關農藝性狀及PUL表達量

圖5 T1-18轉基因植株載體序列的分離

表2 T1-18轉基因植株PUL基因型的分離
2.2 產量相關性狀考察分析
為探明PUL基因突變后對產量相關性狀的影響,我們對T0代突變體和野生型ZH11進行產量性狀考察。
以轉基因系T0-18為代表進行詳細調查,在大田中可以看出T0-18的生育期比ZH11長,前者處于灌漿中期,后者處于灌前初期,生育期相差3~5 d,但是T0-18的株高和野生型沒有差異(圖4 A),RT-PCR結果表明,與野生型相比,T0-18植株中PUL基因的相對表達水平降低了17倍(圖4 B),從RNA水平上驗證了PUL基因被成功敲除。

圖6 稻米品質分析結果
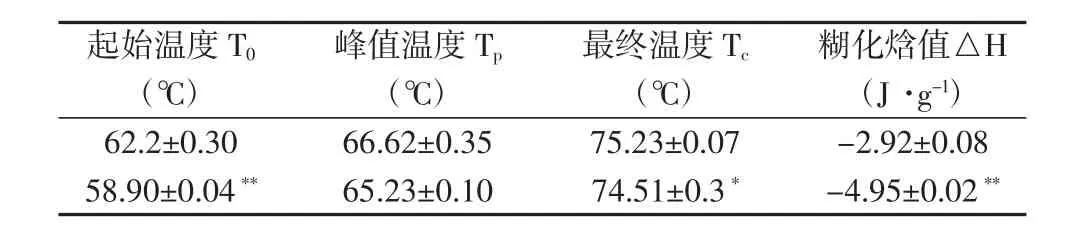
表3 野生型和T0-18胚乳的糊化特性

圖7 T0-18的相關基因表達
RT-PCR結果表明,T0-18的PUL基因的相對表達水平,與野生型相比降低了17倍(圖4 D);從RNA水平上驗證了PUL基因被成功敲除。轉基因系T0-18的千粒重與對照(ZH11)相比增加10.3%(圖4 F)。
粒形分析表明,T0-1、T0-6、T0-11和 T0-18株系的粒長都比ZH11增加,達到顯著差異,其中T0-18的粒長(6.39 mm)比 ZH11的粒長(5.88 mm)增加 8.6%(圖4 B、G);T0-6和T0-18株系的粒寬與ZH11相比顯著提高,而T0-1和T0-11株系的粒寬與ZH11無顯著差異。T0-1和T0-18的長寬比均比野生型ZH11顯著增加(圖4 E、F、G)。此外,轉基因系T0-18的千粒重與野生型ZH11相比增加10.3%(圖4 F)。
2.3 稻米品質分析
普魯蘭酶主要功能是去除極限糊精中的α-1,6糖苷鍵,雖然普魯蘭酶的功能在玉米、水稻、小麥、擬南芥等植物中都證實有上述功能。但是,普魯蘭酶在水稻種子淀粉合成的報道還比較少。為了探討OsPUL敲除后對稻米品質的影響,我們檢測了總淀粉、直鏈淀粉、支鏈淀粉、蛋白質的相對含量(圖6)和糊化溫度。
相比野生型。T0轉基因系的淀粉含量降低,蛋白質含量極顯著增加(圖6),糊化溫度略低0.7℃~2.0℃(表3)。T0-11和T0-18比野生型的直鏈淀粉含量極顯著降低,其他達到顯著水平。相對而言,T0-1、T0-6、T0-11、T0-18比野生型的蛋白質含量都極顯著增加。
2.4 品質及粒形相關基因表達
進一步解釋敲除突變體的稻米品質變化,及粒形、粒重與野生型的差異,我們檢測了品質及粒形相關基因的相對表達量。結果表明,淀粉合成相關基因和粒重相關基因在T0-18種子中的相對表達量有升有降(圖7)。表達量增加的有淀粉合成酶1(SSI)、淀粉分支酶2(BE2),以及正向調控粒形和千粒重的絲氨酸羧肽酶編碼基因GS5[11,20]和正向調控粒長的Gl7[21]。而粒重負調控基因 GS3[16,22]和 GW8[23]的表達量都較低。
3 討論與結論
本文通過CRISPR/Cas9基因編輯技術獲得了PUL基因的不同等位變異植株,且獲得了剔除轉基因成分的純合突變株系,為進一步利用該基因進行水稻遺傳改良提供了重要材料基礎。
FUJITA等[10]認為,OsPUL可以部分恢復isa1癟粒突變體的表型,該基因還影響千粒重,此外OsPUL影響淀粉的晶體結構和淀粉含量。我們研究發現,OsPUL不僅調控淀粉合成,還影響儲藏蛋白的含量。以ZH11作對照,選4個代表性的T0代植株,檢測其淀粉、蛋白質和糊化溫度等稻米營養指標發現,T0-18的直鏈淀粉含量降低了18.70%,其蛋白質含量增加了22.02%,而OsPUL敲除后對稻米外觀品質無顯著差異(圖4 G)。
根據前人報道,在淀粉合成酶基因中,SSI有利于促進支鏈淀粉合成,而SSIIIa則促進支鏈淀粉和超長支鏈淀粉合成[8]。SSI表達量增加,SSIIIa表達量下降(圖7),可以從側面推測參與直鏈淀粉合成的酶減少,同時參與支鏈淀粉合成的酶增加,這與T0代稻米中直鏈淀粉含量降低的結果相佐證(圖6)。PUL屬于淀粉脫支酶(DBEs)家族基因,當pul功能缺失時,并沒有像同家族的isa1(亦稱sug1)突變體一樣導致種子干癟,而是形成稍許腹白,可能淀粉脫支酶家族的基因有功能冗余,ISA1可以彌補pul突變對支鏈淀粉合成的影響。
PUL基因編輯材料可能促進粒長增加,具有增產潛力。已有研究表明,GW8是具有SBP結構域的轉錄因子,可以與GL7的啟動子結合,抑制其表達[21]。根據T0-18株系中粒形相關基因表達分析發現,GW8表達量下降,可導致對GL7的抑制作用減弱,從而使得GL7的表達量升高,導致粒長、粒重增加。T0-18純合突變體千粒重增加,可能是光合產物增加,也可能是粒形粒重相關基因的表達量變化,促使千粒重增加。此外,千粒重的正調控基因GS5和GL7表達量升高,而粒形和粒重的負調控基因GW8和GS3表達量降低[16,22,24],多個粒重相關基因形成復雜的調控網絡,共同作用促進了純合突變體的千粒重增加(圖7)。
