移栽定殖根際有益菌番茄苗的田間效應研究①
2019-10-19 02:11:48鄧旭輝王東升陶成圓王其傳沈其榮
土壤 2019年4期
張 娜,黃 炎,徐 谞,張 博,鄧旭輝,王東升,陶成圓,王其傳,李 榮*,沈其榮
移栽定殖根際有益菌番茄苗的田間效應研究①
張 娜1,黃 炎1,徐 谞1,張 博1,鄧旭輝1,王東升2,陶成圓1,王其傳3,李 榮1*,沈其榮1
(1 江蘇省固體有機廢棄物資源化高技術研究重點實驗室/江蘇省有機固體廢棄物資源化協同創新中心/國家有機肥類肥料工程技術研究中心/南京農業大學資源與環境科學學院,南京 210095;2 南京市蔬菜科學研究所,南京 210042;3 淮安柴米河農業科技發展有限公司,江蘇淮安 223001)
為評估移栽定殖根際有益菌(PGPR)番茄苗對果實產量和青枯病防控效果的影響,通過連續3季田間試驗,研究了基于生物有機肥的施用,移栽生物育苗基質(在普通育苗基質中添加分離自根際的解淀粉芽孢桿菌)所育種苗(BIONS),相比于移栽普通育苗基質所育種苗(BIO),對設施番茄產量、發病率、收獲期植株土體與根際微生物數量和土壤基本理化性質的影響。連續3季田間試驗結果表明:相比于BIO處理,BIONS處理第一、二、三季的增產幅度分別達38.86%、47.87%、34.60%,產量差異均達到顯著性水平;BIONS處理的發病率每季均極顯著低于BIO處理;BIONS處理增加了根際細菌數量,降低了根際真菌數量;土壤基本理化性質方面,BIONS處理的硝態氮含量和銨態氮含量每季均高于BIO處理,且硝態氮含量與產量呈顯著正相關,硝態氮含量和銨態氮含量與發病率呈顯著負相關。因此,以生物有機肥為底肥,移栽生物育苗基質所育種苗,能夠有效防控番茄青枯病的發生,進而提高產量。
番茄;產量;青枯病;生物育苗基質;根際有益菌
隨著設施蔬菜產業的發展,番茄種植面積日益擴大[1],而番茄青枯病是嚴重制約設施番茄可持續生產的主要土傳病害之一[2]。番茄青枯病是由茄科勞爾氏菌()引起的一種細菌病害,其侵染范圍廣,分布具全球性,難以防治且一旦爆發對作物經濟產生毀滅性影響[3-4]。國內外研究者發現,通過土壤化學熏蒸,向土壤中添加有機物質、施用微量元素肥料、生物防控方法等均能降低青枯病發病率[5-8],但防控效果大多不具穩定性[9-11]。
生物有機肥是一種包含特定功能微生物菌種的特殊有機肥料[12],功能微生物與有機物質相結合能有效提高功能微生物的活性和病害防控能力[13]。國內外研究均表明,施用生物有機肥可促進植株產量的增加和品質的提升[14],同時能一定程度上保護作物不受病原菌侵染,從而有效抑制土傳病害的發生[15-17]。已有大量研究證實,生物有機肥能夠有效控制青枯病的發生[18-19];同時研究證實,生物有機肥發揮作用的關鍵在于有益微生物在植物根際的定殖[20-21]。因此,如能夠將生物有機肥中的有益菌在苗期提前定殖于植物根際,預計能夠有效促進作物的產量。已有研究表明,利用芽孢桿菌制成生物基質育苗,能夠促使功能菌苗期定殖于根際,從而提高西瓜的田間生物量和產量[22];孫躍躍等[23]研究結果也表明將生防放線菌等菌劑加入育苗基質育苗,可顯著提高甜瓜的生物量及抗逆性。然而,將生物育苗基質和生物有機肥聯合田間施用,對防控作物土傳病害的研究還少有報道,尤其是連續應用的效果,還缺少研究。
本研究采用連續3季田間試驗,以生物有機肥為底肥,評估了在育苗基質中添加有益菌T-5促使其在移栽大田前定殖于番茄根際,對番茄青枯病防控及產量增加的影響,以期為設施番茄高產高效栽培提供實際指導和理論支撐。
1 材料與方法
1.1 供試材料
供試番茄品種為“世紀紅冠”。供試菌株解淀粉芽孢桿菌T-5,由江蘇省固體有機廢棄物資源化高技術研究重點實驗室提供,該菌株能夠定殖于番茄根際,從而有效防控青枯病的發生[24-25]。
供試生物有機肥由本實驗室自行研制[26],產品中菌株T-5含量為6 × 108cfu/g(以干物質量計)。
供試普通育苗基質由南京市蔬菜科學研究所提供。
1.2 試驗設計
田間試驗于2014年3月至2015年6月(共3季)在南京市蔬菜科學研究所大棚內進行。供試土壤為黃壤土,基本理化性質為:pH 6.33、有機質43.56 g/kg、全氮1.89 g/kg、有效磷138.66 mg/kg、速效鉀464.81 mg/kg。
田間試驗共設置2個處理:①施用生物有機肥并移栽普通育苗基質所育種苗(BIO)處理,生物有機肥的施肥量為5 997 kg/hm2;②施用生物有機肥并移栽生物育苗基質所育種苗(BIONS)處理,生物有機肥的施入量同BIO處理。番茄全生育期所需其余養分用化肥補齊。每個處理設置3個重復,每個重復小區面積為10 m2,種植密度為每個小區32棵番茄,每季番茄種植時間為3個月。
本試驗生物育苗基質的制備及育苗步驟為:將解淀粉芽孢桿菌T-5在LB培養基中發酵以制備菌液[27];將菌液按5% (/,為干物質量) 比例與普通基質混合均勻制成生物育苗基質[22]。將番茄種子進行表面消毒后浸種催芽,待種子露白后分別埋于普通育苗基質和生物育苗基質;待番茄幼苗長出第二片真葉時移栽至田間。普通育苗基質所育種苗移栽于BIO處理,生物育苗基質所育種苗移栽于BIONS處理。
1.3 樣品采集及測定
1)產量和發病率統計。在番茄進入收獲期后,采摘番茄全部果實記產量,并統計番茄發病率,發病率表示為具青枯病癥狀植株占總植株數的百分比。
2)土壤樣品的采集。于番茄收獲期,各小區隨機選擇3株健康植株,連根帶土采集完后送至實驗室進行土體和根際土壤樣品的分離[28],各小區內土體和根際樣品分別進行合并為該小區代表性樣品,保存于超低溫冰箱中備用。
3)DNA的提取及測定。稱取0.25 g土壤樣品,用土壤DNA提取試劑盒 (Mo Bio Laboratories, Inc., Carlsbad, CA, USA),按操作說明提取土壤DNA。采用實時熒光定量PCR方法測定土壤樣品中病原菌、總細菌、總真菌數量。病原菌擴增引物選用flic F (5’- GAACGCCAACGGTGCGAACT-3’) 和flic R (5’-GG-CGGCCTTCAGGGAGGTC-3’)[29];細菌擴增引物選用338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和518R (5’-ATTACCGCGGCTG CTGG-3’)[30];真菌擴增引物選用ITS1f(5’-TCCGTAGGTGAACCTGCGG-3’)和5.8S(5’-CGCTGCGTTCTTCATCG-3’)[30]。定量使用儀器為ABI PRISM? 7500 Real-time PCR system,反應體系見表1。反應程序為:95 ℃預變性30 s,95 ℃變性5s,53 ℃退火和延伸34 s (病原菌的退火和延伸溫度為56 ℃),循環30次。每個樣品設置3次重復,用無菌超純水替代DNA模板設置陰性對照。根據樣品閾值 (Ct) 計算每克土中的拷貝數,結果以拷貝數取對數后表示(log (Copies/g,以干物質量計))。

表1 定量PCR擴增體系
4)土壤基本理化性質的測定。土壤理化性質的測定參照《土壤農化分析》[31]進行。
1.4 數據統計與分析
使用Microsoft Excel 2007對數據進行處理;用SPSS軟件 (20.0, SPSS Inc. Chicago, IL) 進行獨立樣本檢驗來分析兩處理之間的差異;用R(version 3.2.0)軟件進行土壤微生物數量、基本理化性質與番茄發病率和產量間的Spearman相關性分析,顯著水平設定為<0.05。
2 結果與分析
2.1 連續3季移栽生物育苗基質所育種苗對設施番茄產量的影響
由表2可知,3季田間試驗,施用生物有機肥后,移栽根際定殖功能菌的種苗處理(BIONS)單株產量均高于單施生物有機肥處理(BIO),但兩處理間均無顯著差異。每公頃產量BIONS處理均顯著高于BIO處理。第一季BIONS處理比BIO處理顯著增產18 695.44 kg/hm2,增產幅度達38.86%;第二季顯著增產19 578.39 kg/hm2,增產幅度達47.87%;第三季顯著增產14 510.63 kg/hm2,增產幅度達34.60%。
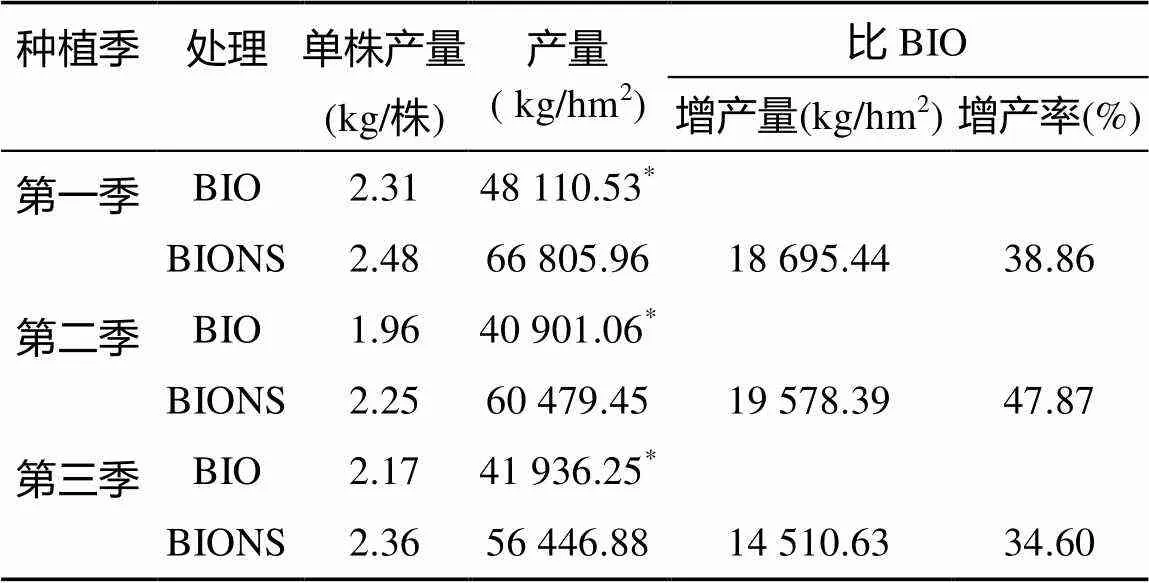
表2 不同處理對番茄產量的影響
注:*表示兩處理在<0.05水平差異顯著,表3同。
2.2 連續3季移栽生物育苗基質所育種苗對設施番茄發病率的影響
由圖1可知,單施生物有機肥處理(BIO)3季田間番茄青枯病發病率分別為34.38%、34.46% 和40.00%,移栽生物育苗基質所育種苗處理(BIONS)的發病率分別為15.63%、16.11% 和25.00%,與BIO處理相比,BIONS處理顯著降低了3季番茄青枯病的發病率,分別達到了54.55%、53.25% 和37.50% 的田間防控效果。
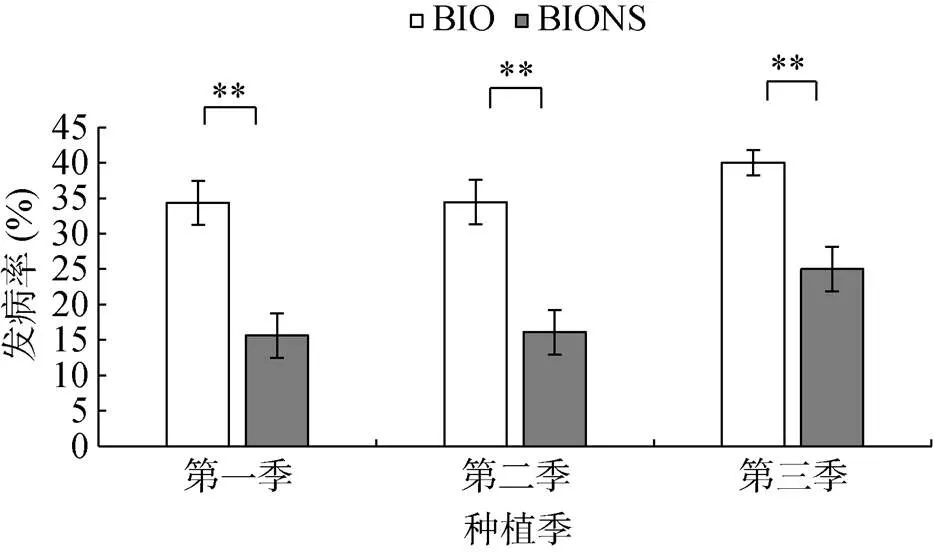
(**表示兩處理在P<0.01水平差異顯著)
2.3 連續3季移栽生物育苗基質所育種苗對土壤病原菌、總細菌和總真菌數量的影響
由圖2A和2B可知,在第一季和第三季田間試驗中,移栽生物育苗基質所育種苗處理(BIONS)和單施生物有機肥處理(BIO)的土體土壤病原菌數量無顯著性差異,第二季BIONS處理土體土壤中病原菌數量顯著低于BIO處理。第一季和第二季BIONS處理番茄根際土壤中病原菌數量高于BIO處理但無顯著性差異,第三季BIONS處理根際病原菌數量極顯著低于BIO處理。
由圖2C和2D可知,第一季BIONS處理土體總細菌數量高于BIO處理但無顯著性差異,第二季和第三季均低于BIO處理,且在第二季兩處理間具有顯著性差異。BIONS處理根際總細菌數量在3季田間試驗中均高于BIO處理,第二季兩處理間具有顯著性差異。
由圖2E和2F可知,3季BIONS處理土體總真菌數量均低于BIO處理,但不具有顯著性差異。第一季和第三季BIONS處理的根際總真菌數量極顯著低于BIO處理。
2.4 生物育苗基質聯合生物有機肥連續施用對土壤基本理化性質的影響
由表3可知,在第一季田間試驗中,生物有機肥處理(BIO)的pH顯著高于移栽生物育苗基質所育種苗處理(BIONS),BIO處理的銨態氮含量顯著低于BIONS處理,兩處理土壤在其余基本理化性質上無顯著性差異。第二季,BIO處理的速效鉀含量和硝態氮含量顯著低于BIONS處理。第三季,土壤基本理化性質在兩處理間無顯著差異。從總體上看,連續三季田間試驗BIO處理的硝態氮含量和銨態氮含量均低于BIONS處理,其余土壤理化性質無變化規律。
2.5 土壤微生物及理化指標與番茄產量的相關性
如表4所示,番茄發病率與產量呈顯著負相關(rho= –0.46,=0.000 10),細菌數量與產量呈顯著正相關(rho=0.40,=0.002 6),真菌數量與產量呈顯著負相關(rho= –0.32,=0.019)。土壤基本理化性質中,pH、速效鉀含量、有效磷含量、硝態氮含量和銨態氮含量均與產量呈正相關,其中pH、有效磷含量和硝態氮含量與產量呈顯著正相關(rho=0.32,=0.020;rho=0.33,=0.014;rho=0.28,=0.039,電導率與產量呈負相關,但相關性不顯著。
2.6 土壤微生物及理化指標與番茄青枯病發病率的相關性
如表5所示,3季田間試驗,病原菌數量與番茄青枯病發病率呈顯著正相關(rho=0.32,=0.019),其余無顯著相關性。土壤理化性質中,pH、電導率、有效磷含量、硝態氮含量和銨態氮含量均與番茄青枯病發病率呈負相關,但只有硝態氮和銨態氮含量與青枯病發病率呈顯著負相關(rho= –0.38,=0.004 9;rho= –0.51,=0.000 10),速效鉀含量與青枯病發病率呈正相關,但相關性不顯著。
3 討論
3季田間試驗BIONS處理的產量均顯著高于BIO處理,但單株產量兩處理間無顯著差異。已有研究表明生物育苗基質可培育出優質辣椒和番茄優質種苗,在移栽至大田后能有效提高辣椒和番茄的生物量和產量[32-33],但本研究中單株產量雖有增加,但兩處理間差異無顯著性,且Spearman相關性分析表明番茄產量與田間青枯病發病率和土壤真菌數量呈顯著負相關,與土壤細菌數量和硝態氮含量呈顯著正相關,因此,推測增產主要在于番茄青枯病的防控。
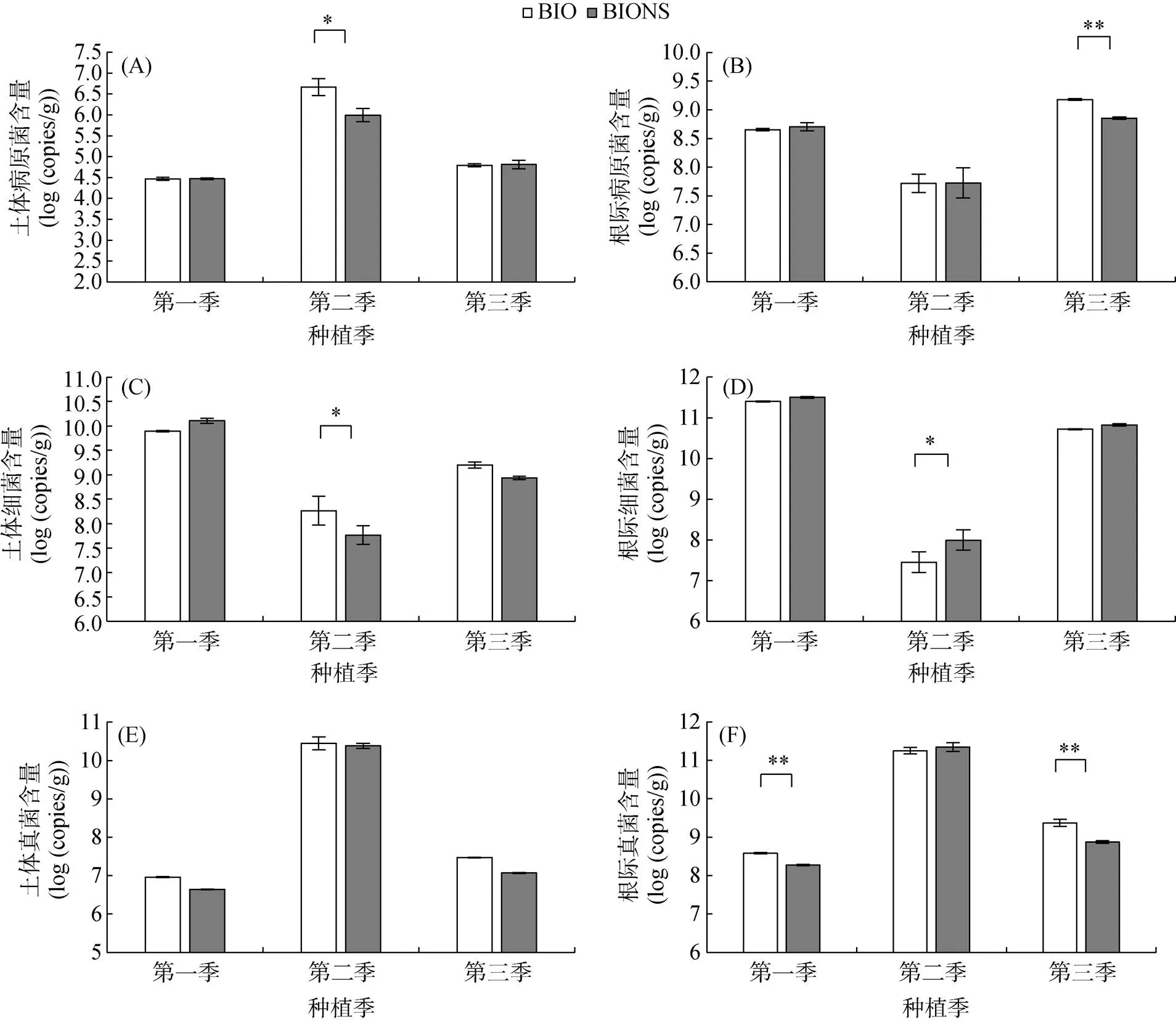
(圖A、C、E為土體土指標,B、D、F為根際土指標;*、**分別表示兩處理在P<0.05、P<0.01水平差異顯著)
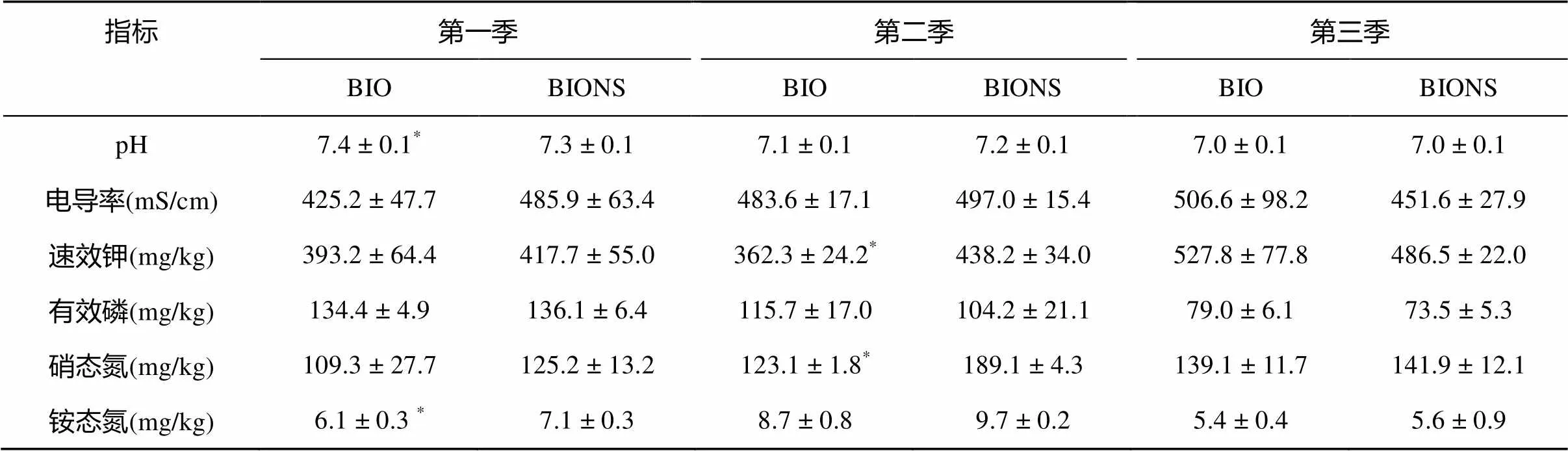
表3 不同處理對土壤理化性質的影響

表4 土壤微生物數量及基本理化性質與番茄產量的Spearman相關性
注:番茄產量與土壤微生物數量及基本理化性質的相異矩陣基于Bray-Curtis距離。

表5 土壤微生物數量及基本理化性質與番茄青枯病發病率的Spearman相關性
注:番茄青枯病發病率與微生物數量和土壤基本理化性質的相異矩陣基于Bray-Curtis距離。
相比BIO處理,BIONS處理顯著降低了番茄青枯病的發病率,Spearman相關性分析表明青枯病發病率與病原菌數量呈顯著負相關,且連續種植3季后,BIONS處理根際病原菌數量顯著低于BIO處理。與本研究結果類似,張鈺等[34]研究表明利用微生物菌劑制備的育苗基質育苗,能降低黃瓜根際枯萎病病原菌數量,進而顯著降低黃瓜枯萎病的發病率;本實驗室前期研究同樣表明,施用生物有機肥能夠有效防控番茄病害[35]。本研究基于此進一步證實了,在施用生物有機肥基礎上,進一步移栽利用生物育苗基質所育根際定殖功能菌的番茄苗能夠進一步降低根際病原菌數量,進而有效防控青枯病的發生,增加作物產量。
土壤微生物與植物病害息息相關[36],通過調控土壤微生物區系來增強土壤抑病能力,能有效防控土傳病害的發生[37]。在本研究中,根際細菌數量BIONS處理3季均高于BIO處理,而根際真菌數量第一季和第三季顯著低于BIO處理。Ling等[38]研究發現,連續施用生物有機肥有能效防控西瓜枯萎病的發生,與根際細菌群落出現顯著變化有關。何欣等[39]在香蕉幼苗期育苗和移栽時均施用結合SQR21和T37復合拮抗菌劑堆制生物有機肥,有效防控了香蕉枯萎病的發生,同時發現根際微生物組成發生了變化,其顯著增加了根際細菌數量同時真菌數量顯著降低。Shen等[28]研究表明施用生物有機肥能有效防控香蕉枯萎病的發生,同時能優化其土壤微生物區系,增加其中細菌和放線菌等的數量。鐘書堂等[40]的研究也得到了相同的結果。本研究與以上結果一致,番茄根際預定殖有益微生物同時聯用生物有機肥優化了根際微生物群落。
從總體上看,連續3季田間試驗,BIO處理的土壤硝態氮和銨態氮含量均低于BIONS處理,且其含量與番茄青枯病發病率呈顯著負相關,同時硝態氮含量與番茄產量呈顯著正相關。氮素對植物的生長至關重要,影響著植物病害和抗性[41-42],不同氮素形態對病害的防治作用也不同[43-44]。有研究表明銨態氮能有效降低尖孢鐮刀菌番茄專化型病原菌的致病力[45],Elmer等[46]的研究也同樣表明銨態氮能一定程度上降低草莓黑腐病的發病率。董鮮等[47]研究表明硝態氮能通過誘導香蕉幼苗木質素的形成進而提高其抗病性,有效防控香蕉枯萎病的發生。本研究結果顯示,連續3季BIONS處理中硝態氮和銨態氮含量均高于BIO處理,這可能也是根際定殖功能菌T-5降低番茄青枯病發生的原因。
綜上,本研究表明生物有機肥與生物育苗基質聯用能顯著提高番茄產量,且有效防控番茄青枯病的發生,研究結果能夠為生物育苗基質的研發及番茄青枯病的防御提供新思路。
4 結論
相比于單施生物有機肥處理(BIO),育苗基質和生物有機肥聯合處理(BIONS)能顯著降低番茄青枯病發病率,提高番茄產量;BIONS處理番茄青枯病發病率顯著降低,與番茄根際病原菌數量降低和土壤硝態氮含量和銨態氮含量較高等顯著相關;番茄青枯病的有效防控導致BIONS處理番茄產量的有效增加。
[1] 葛曉穎, 孫志剛, 李濤, 等. 設施番茄連作障礙與土壤芽孢桿菌和假單胞菌及微生物群落的關系分析[J]. 農業環境科學學報, 2016, 35(2): 514–523
[2] 蔡祖聰, 黃新琦. 土壤學不應忽視對作物土傳病原微生物的研究[J]. 土壤學報, 2016, 53(2): 305–310
[3] Caruso P, Palomo J L, Bertolini E, et al. Seasonal variation ofbiovar 2 populations in a Spanish river: Recovery of stressed cells at low temperatures[J]. Applied and Environment Microbiology, 2005, 71(1): 140–148
[4] Allen C, Prior P, Hayward A C, et al. Bacterial wilt disease and thespecies complex[J]. Bacterial Wilt Disease & the Species Complex, 2005
[5] Hsu S T. Ecology and control ofin Taiwan[J]. Plant Protection Bulletin Taiwan, 1991, 33: 72–79
[6] Bailey K L, Lazarovits G. Suppressing soil-borne diseases with residue management and organic amendments[J]. Soil and Tillage Research, 2003, 72: 169–180
[7] Kiirika L M, Stahl F, Wydra K. Phenotypic and molecular characterization of resistance induction by single and combined application of chitosan and silicon in tomato against[J]. Physiological and Molecular Plant Pathology, 2013, 81: 1–12
[8] Ma Y, Gentry T, Hu P, et al. Impact of brassicaceous seed meals on the composition of the soil fungal community and the incidence of Fusarium wilt on chili pepper[J]. Applied Soil Ecology, 2015, 90: 41–48
[9] Gamliel A, Austerweil M, Kritzman G. Non-chemical approach to soilborne pest management-organic amendments[J]. Crop Protection, 2000, 19: 847–853
[10] Bailey K L, Lazarovits G. Suppressing soil-borne diseases with residue management and organic amendments[J]. Soil and Tillage Research, 2003, 72: 169–180
[11] 陳志龍, 陳杰, 許建平, 等. 番茄青枯病生物防治研究進展[J]. 江蘇農業科學, 2013, 41(8): 131–134
[12] 楊興明, 徐陽春, 黃啟為, 等. 有機(類)肥料與農業可持續發展和生態環境保護[J]. 土壤學報, 2008, 45(5): 925– 932
[13] Kavino M, Harish S, Kumar N, et al. Effect of chitinolytic PGPR on growth, yield and physiological attributes of banana (, spp.) under field conditions[J]. Applied Soil Ecology, 2010, 45(2): 71–77
[14] 韋巧婕, 鄭新艷, 鄧開英, 等. 黃瓜枯萎病拮抗菌的篩選鑒定及其生物防效[J]. 南京農業大學學報, 2013, 36(1): 40–46
[15] Qiu M H, Zhang R F, Xue C, et al. Application of bio-organic fertilizer can controlof cucumber plants by regulating microbial community of rhizosphere soil[J]. Biology & Fertility of Soils, 2012, 48(7): 807–816
[16] Shen Z Z, Wang B B, Lv N N, et al. Effect of the combination of bio-organic fertilizer withNJN-6 on the control of banana fusarium wilt disease, crop production and banana rhizosphere culturable microflora[J]. Biocontrol Science and Technology, 2015, 25(6): 716–731
[17] Saravanan T, Muthusamy M, Marimuthu T. Development of integrated approach to manage the fusarial wilt of banana[J]. Crop Protection. 2003, 22: 1117–1123
[18] 丁傳雨, 喬煥英, 沈其榮, 等. 生物有機肥對茄子青枯病的防治及其機理探討[J]. 中國農業科學, 2012, 45(2): 239–245
[19] 袁英英, 李敏清, 胡偉, 等. 生物有機肥對番茄青枯病的防效及對土壤微生物的影響[J]. 農業環境科學學報, 2011, 30(7): 1344–1350
[20] 年洪娟, 陳麗梅. 土壤有益細菌在植物根際競爭定殖的影響因素[J]. 生態學雜志, 2010, 29(6): 1235–1239
[21] 李文英, 彭智平, 楊少海, 等. 植物根際促生菌對香蕉幼苗生長及抗枯萎病效應研究[J]. 園藝學報, 2012, 39(2): 234–242
[22] 張楊, 王甜甜, 孫玉涵, 等. 西瓜根際促生菌篩選及生物育苗基質研制[J]. 土壤學報, 2017, 54(3): 703–712
[23] 孫躍躍, 馬軍妮, 李玉龍, 等. 功能性育苗基質中生防菌及腐植酸鉀對甜瓜穴盤苗的促生作用[J]. 西北農林科技大學學報(自然科學版), 2016, 44(9): 174–180
[24] Tan S, Yang C, Mei X, et al. The effect of organic acids from tomato root exudates on rhizosphere colonization ofT-5[J]. Applied Soil Ecology, 2013, 64(1): 15–22
[25] Tan S, Jiang Y, Song S, et al. Twostrains isolated using the competitive tomato root enrichment method and their effects on suppressingand promoting tomato plant growth[J]. Crop Protection, 2013, 43: 134–140
[26] Huang Y, Sun L, Zhao J S, et al. Utilization of different waste proteins to create a novel PGPR-containing bio-organic fertilizer[J]. Scientific Reports, 2015, 5(7766): 1–11
[27] Tan S, Gu Y, Yang C, et al.T-5 may preventinfection through competitive exclusion[J]. Biology & Fertility of Soils, 2016, 52(3): 341–351
[28] Shen Z Z, Zhong S T, Wang Y G, et al. Induced soil microbial suppression of banana fusarium wilt disease using compost and biofertilizers to improve yield and quality[J]. European Journal of Soil Biology, 2013, 57(4): 1–8
[29] Sch?nfeld J, Heuer H, Van E J, et al. Specific and sensitive detection ofin soil on the basis of PCR amplification offragments[J]. Applied and Environmental Microbiology, 2003, 69(12): 7248–7256
[30] Fierer N, Jackson J A, Vilgalys R, et al. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays[J]. Applied and Environmental Microbiology, 2005, 71(7): 4117–4120
[31] 鮑士旦. 土壤農化分析[M]. 北京: 中國農業出版社, 2000
[32] 文春燕, 高琦, 張楊, 等. 含PGPR菌株生物育苗基質的研制與促生效應研究[J]. 土壤, 2016, 48(2): 414–417
[33] 張楊, 文春燕, 趙買瓊, 等. 辣椒根際促生菌的分離篩選及生物育苗基質研制[J]. 南京農業大學學報, 2015, 38(6): 950–957
[34] 張鈺, 孫錦, 郭世榮. 基質中添加微生物制劑對黃瓜幼苗生長和枯萎病抗性的影響[J]. 西北植物學報, 2013, 33(4):780–786
[35] Liu H, Xiong W, Zhang R, et al. Continuous application of different organic additives can suppress tomato disease by inducing the healthy rhizospheric microbiota through alterations to the bulk soil microflora[J]. Plant and Soil, 2018, 423: 229–240
[36] Elsas J D V, Garbeva P, Salles J. Effects of agronomical measures on the microbial diversity of soils as related to the suppression of soil-borne plant pathogens[J]. Biodegradation, 2002, 13(1): 29–40
[37] 劉金光, 李孝剛, 王興祥. 連續施用有機肥對連作花生根際微生物種群和酶活性的影響[J]. 土壤, 2018, 50(2): 305–311
[38] Ling N, Deng K, Song Y, et al. Variation of rhizosphere bacterial community in watermelon continuous mono- cropping soil by long-term application of a novel bioorganic fertilizer[J]. Microbiological Research, 2014, 169(7): 570–578
[39] 何欣, 郝文雅, 楊興明, 等. 生物有機肥對香蕉植株生長和香蕉枯萎病防治的研究[J]. 植物營養與肥料學報, 2010, 16(4): 978–985
[40] 鐘書堂, 沈宗專, 孫逸飛, 等. 生物有機肥對連作蕉園香蕉生產和土壤可培養微生物區系的影響[J]. 應用生態學報, 2015, 26(2): 481–489
[41] Marschner H. Book Reviews: Mineral nutrition in higher plants[J]. Plant, cell & Environment, 1988, 11: 147–148
[42] Dordas C. Role of nutrients in controlling plant diseases in sustainable agriculture: A review[J]. Agronomy for Sustainable Development, 2008, 28(1): 33–46
[43] Huber D M, Haneklaus S. Managing nutrition to control plant disease[J]. Landbauforschung Volkenrode, 2007, 4(4): 313–322
[44] Borrero C, Trillas M I, Delgado A, et al. Effect of ammonium/nitrate ratio in nutrient solution on control of Fusarium wiltof tomato by, T34[J]. Plant Pathology, 2012, 61(1): 132–139
[45] Lópezberges M S, Rispail N, Pradosrosales R C, et al. A nitrogen response pathway regulates virulence functions invia the Protein Kinase TOR and the b ZIP Protein MeaB[J]. Plant Cell, 2010, 22(7): 2459–2475
[46] Elmer W H, LaMondia J A. Influence of ammonium sulfate and rotation crops on strawberry black root rot[J]. Plant Disease, 1999, 83(83): 119–123
[47] 董鮮, 鄭青松, 王敏, 等. 銨態氮和硝態氮對香蕉枯萎病發生的比較研究[J]. 植物病理學報, 2015, 45(1): 73–79
Effects of Seedlings Colonized PGPR Stains on Bacterial Wilt Disease Suppression and Yield of Tomato
ZHANG Na1, HUANG Yan1, XU Xu1, ZHANG Bo1, DENG Xuhui1, WANG Dongsheng2, TAO Chengyuan1, WANG Qichuan3, LI Rong1*, SHEN Qirong1
(1 Jiangsu Key Laboratory of Solid Organic Waste Utilization, Jiangsu Collaborative Innovation Center for Solid Organic Waste Resource Utilization,National Engineering Research Center for Organic-based Fertilizers, College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095, China; 2 Nanjing Institute of Vegetable Science, Nanjing 210042, China; 3 Huaian Chaimihe Agricultural Technology Development Co. Ltd., Huaian, Jiangsu 223001, China)
The effects of seedlings colonized PGPR stains on tomato bacterial wilt disease suppression and yield were investigated via continuously 3 season field experiments in this study for providing a new strategy to maintain the tomato sustainable production. Compared with the control (BIO) transplanted with common seedlings cultivated in ordinary nursery substrate, seedlings colonized PGPR stain produced by bio-nursery substrate were transplanted in the field amended with bio-organic fertilizer (BIONS) to evaluate the tomato yield enhancement and bacterial wilt disease suppression ability as well as its influence on microbial biomass and soil properties in a plastic greenhouse. Results showed that compared to BIO, BIONS significantly increased tomato yield by 38.86%, 47.87% and 34.60% in the first, second and third seasons, respectively. Meanwhile, tomato bacterial wilt disease incidence of BIONS was prominently lower than BIO. Moreover, BIONS application significantly increased rhizosphere bacteria but decreased fungi populations. During the three seasons, the contents of ammonium nitrogen (NO– 3-N) and nitrate nitrogen (NH4+-N) were higher in BIONS than in BIO; ammonium nitrogen amount had a positive correlation with tomato yield; and negative correlation was observed between disease incidence and the contents of ammonium nitrogen and nitrate nitrogen. The above results prove that continuous application of the bio-nursery substrates in greenhouse can effectively control tomato bacterial wilt disease, thus can increase tomato yields.
Tomato; Yield; Bacterial wilt; Bio-nursery substrate; Rhizosphere beneficial bacteria
江蘇省科技計劃項目(BY2016077-05)、國家重點研發計劃項目(2016YFE0101100和2016YFD0800605)、江蘇省青年自然科學基金項目(BK20160710)、江蘇省高校品牌專業建設工程資助項目(PPZY2015A061)和校大學生創新訓練項目(1513A09)資助。
(lirong@njau.edu.cn)
張娜(1991—),女,新疆阿克蘇人,碩士研究生,主要研究領域為植物營養學。E-mail:2018203032@njau.edu.cn
S365
A
10.13758/j.cnki.tr.2019.04.005
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
科學大眾(2021年9期)2021-07-16 07:02:54
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
軍事文摘(2020年20期)2020-11-28 11:42:50
今日農業(2020年20期)2020-11-26 06:09:10
航空世界(2020年10期)2020-01-19 14:36:20
