模擬氮沉降對貝加爾針茅草原土壤氮轉化微生物的影響
2019-10-23 06:01:06劉紅梅張海芳張艷軍楊殿林
農業環境科學學報 2019年10期
關鍵詞:研究
劉紅梅,張海芳,秦 潔,王 慧,張艷軍,楊殿林
(農業農村部環境保護科研監測所,農業農村部產地環境污染防控重點實驗室/天津市農業環境與農產品安全重點實驗室,天津300191)
大氣氮沉降增加呈現全球化趨勢,我國是繼歐洲、北美之后的第3大氮沉降區,大氣氮沉降遠高于全球平均水平[1]。大氣氮沉降借助其對土壤碳固定[2]、植物氮素利用等的直接或間接作用[3],極大地干預了生態系統碳蓄積和氮素重新分配過程[4-5]。高氮沉降導致土壤氮轉化過程發生變化[6]、土壤可利用性氮含量增加[7]、氮循環微生物數量及組成發生變化[8],進而對生態系統產生影響。
土壤氮轉化主要過程包括生物固氮作用、氨化作用、硝化作用和反硝化作用,這些過程都由土壤微生物所驅動。氮沉降進入土壤后的一系列轉化離不開與氮素轉化的相關微生物參與。近年來的研究表明,施氮肥顯著影響固氮菌、硝化菌和反硝化菌群落結構和豐度[8]。適量氮肥添加提高固氮菌豐度,促進土壤微生物的固氮功能[9];高濃度無機氮添加抑制固氮菌生長[10],從而抑制固氮微生物生長。Ning等[11]研究發現氮相關功能基因對氮添加速率表現出不同的敏感性,氨氧化細菌AOB-amoA基因比氨氧化古菌AOA-amoA基因對氮添加更加敏感。土壤氨氧化細菌豐度隨氮添加量增加而增加,而氨氧化古菌豐度則無明顯變化[12]。氮肥種類和氮肥施用量會影響土壤氨氧化微生物組成和豐度[13]。受氮素限制的黃土高原鹽堿草地,施氮后顯著地提高了微生物生物量氮轉化速率[14]。在亞熱帶森林、溫帶森林中研究發現,N2O排放、CH4吸收與AOA、AOB群落豐度分別呈現正、負相關關系,氨氧化菌群落動態能夠解釋土壤CH4吸收和N2O排放之間的消長作用[15]。
目前,在氮沉降增加的背景下,針對干旱和半干旱草原土壤氮轉化功能基因豐度對氮沉降增加的響應特征還不夠明確。為此,本研究通過施氮模擬氮沉降增加,研究不同氮添加水平對涉及土壤氮循環中的固氮nifH基因、氨氧化細菌AOB-amoA基因、氨氧化古菌AOA-amoA基因和反硝化基因nirK豐度的影響,通過分析氮轉化功能基因豐度與土壤理化因子之間的相關性關系,研究氮添加對微生物介導的固氮、氨氧化和反硝化過程的影響及其反饋,為深入認識草地生態系統氮循環及其對氮沉降增加的響應機制提供參考。
1 材料與方法
1.1 研究區域概況與樣地設置
研究區域概況、試驗樣地設置及試驗前植被類型和基本理化性質詳見參考文獻[16]。試驗樣地位于內蒙古貝加爾針茅草原(48°27′~48°35′N,119°35′~119°41′E)。于2010年開始進行圍欄試驗,氮添加水平設置參考國際上同類研究試驗[17-18]。氮添加處理設8個水平,依次為:0、15、30、50、100、150、200、300 kg N·hm-2·a-1,分別記為:對照N0,低氮(N15、N30和N50),高氮(N100、N150、N200和N300)。
氮素為NH4NO3,每年6月中旬和7月中旬進行氮添加。共設置8個處理水平,4次重復。小區面積8 m×8 m,小區間設置2 m隔離帶,重復間設置5 m隔離帶。
1.2 樣品采集與處理
土壤樣品采集于2015年8月中旬,用土鉆在各小區采集0~10 cm土壤樣品,每小區采集10個點,混合均勻。去除土壤入侵物和根系,將土壤樣品分為兩份,一份保存于冰盒中帶回實驗室,存于-20℃冰柜中;一份土壤樣品于室內自然風干。
1.3 測定方法
土壤理化性質測定:土壤pH采用玻璃電極法(土水比1∶2.5),土壤總有機碳測定采用重鉻酸鉀外加熱法,土壤全氮用凱氏定氮法,土壤銨態氮和硝態氮含量采用氯化鉀溶液提取-流動分析儀(QC8000)測定[19]。土壤微生物生物量氮采用氯仿熏蒸-K2SO4提取-TOC儀(Multi N/C 3000)測定[20]。土壤可溶性氮以測定土壤微生物生物量氮時未熏蒸土壤的全氮含量表示,土壤可溶性有機氮以土壤可溶性氮減去銨態氮和硝態氮的差值表示[21]。
土壤DNA提取:稱取0.25 g在冰柜中保存的土壤樣品,用PowerSoil?DNA Isolationkit試劑盒,按照說明書提取土壤總DNA。用Nanodrop2000超微量分光光度計和1%瓊脂糖電泳檢測DNA質量。將通過質量檢測的DNA樣品于-20℃冰箱保存待用。
標準品配制:用目的基因引物進行PCR擴增。PCR反應體系為50 μL,包括上下游引物各1 μL,模板1 μL,2×Taq MasterMix 25 μL,滅菌超純水22 μL。擴增條件為94℃5 min預變性后;94℃30 s,55℃30 s,72℃ 30 s,共30個循環;最后1輪循環完成后再72℃延伸10 min。反應完畢后,用1%瓊脂糖凝膠電泳檢查擴增結果,目的片段進行瓊脂糖凝膠回收。將回收PCR產物連接至pMD18-T載體,轉化至大腸桿菌DH5α感受態細胞中,經Amp+、IPTG和X-gal的LB平板篩選陽性克隆,送去生工生物有限公司進行序列測定分析。
標準曲線制作:通過做預實驗,將4種提取測序正確的質粒標準品從101~105倍進行10倍稀釋,每個梯度取2 μL做模板建立標準曲線。
定量PCR檢測:土壤固氮基因nifH、氨氧化細菌AOB、氨氧化古菌AOA和反硝化細菌定量擴增引物見表1。擴增反應體系總體積為20 μL,包括2×GoT-aq?qPCR Master Mix 10 μL,10 μmol·L-1的上下游引物各0.5 μL,DNA模板2 μL(1~10 ng),滅菌超純水7 μL。將加好樣的96-PCR板置于熒光定量PCR儀進行擴增反應,每個樣品3個重復。擴增反應條件為95℃預變性30 s;95℃ 變性5 s,60℃退火40 s,72℃延伸30 s,40個循環。
1.4 數據處理
應用SPSS 17.0統計軟件進行單因素方差分析(One-way ANOVA)、Pearson相關性分析。
2 結果與分析
2.1 氮沉降對土壤化學性質的影響
連續6年不同氮添加處理下,土壤化學性質變化見表2。高氮添加(N100、N150、N200和N300)土壤有機碳、硝態氮和微生物生物量氮含量高于或顯著高于低氮添加(N15、N30和N50)和對照N0。7個氮添加處理的土壤pH均低于或顯著低于對照N0,且高氮添加pH顯著低于低氮添加。土壤全氮含量在不同氮添加處理間無顯著差異。
2.2 氮沉降對土壤氮轉化功能基因豐度的影響
固氮微生物功能基因nifH豐度在各氮添加處理中的變化范圍在5.26×105~1.43×106copies·g-1soil之間(圖1A)。隨著氮添加水平的升高,nifH豐度表現為先升高后降低的趨勢,其中N50處理是對照N0的2.39倍。N15、N30、N50、N100、N150和 N200處理nifH豐度高于或顯著高于對照N0。N300處理nifH豐度低于對照N0,但無顯著差異(P>0.05)。

表1 定量PCR擴增引物Table 1 Amplification primer of quantitative PCR

表2 不同氮添加處理下土壤化學性質Table 2 Soil chemical properties in different nitrogen addition treatments
氨氧化細菌AOB-amoA基因豐度在各氮添加處理中的變化范圍為1.33×107~2.48×107copies·g-1soil之間(圖1B)。N15、N30和N50處理AOB-amoA基因豐度與對照相比無顯著差異(P>0.05),高氮添加(N100、N150、N200和N300)AOB-amoA基因豐度均顯著高于對照N0(P<0.05)。表明氮沉降增加使AOB-amoA基因豐度提高。氨氧化古菌AOA-amoA基因豐度在各氮添加處理中的變化范圍為2.56×106~3.97×106copies·g-1soil之間(圖1C)。N30處理AOA-amoA基因豐度最高,但與對照相比無顯著差異,其他6個氮添加處理均低于對照N0。土壤中AOB的基因拷貝數遠高于AOA基因拷貝數,且高氮添加顯著提高了AOB/AOA比值(圖1E)。
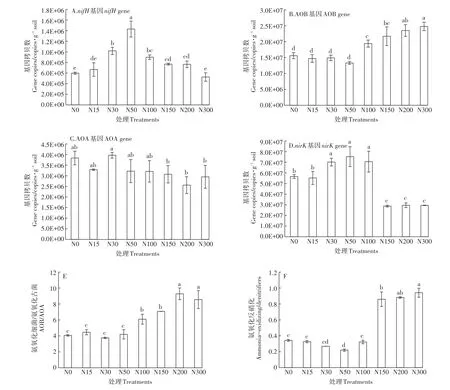
圖1 不同氮處理的功能基因豐度Figure 1 Functional genes in different nitrogen treatments
土壤反硝化功能基因nirK豐度在不同氮添加處理中的變化范圍為 2.88×107~7.53×107copies·g-1soil之間(圖1D)。N15處理nirK基因豐度低于對照N0,但無顯著差異(P>0.05);N30、N50和N100處理nirK基因豐度均顯著高于對照N0(P<0.05);高氮添加處理(N150、N200和N300)nirK基因豐度顯著低于對照N0(P<0.05),顯著提高了氨氧化/反硝化豐度比值(圖1F)。

表3 功能基因拷貝數與化學性質之間的相關性分析Table 3 Correlation analysis between functional gene copies and soil chemical properties
2.3 土壤氮素轉化功能基因豐度與土壤因子的相關性
土壤氮轉化功能基因拷貝數與土壤化學性質相關性分析見表3。土壤nifH基因拷貝數與總有機碳、硝態氮和微生物生物量氮含量呈顯著負相關(P<0.05),與土壤銨態氮含量呈極顯著正相關(P<0.01)。土壤AOB-amoA基因拷貝數與有機碳、硝態氮、可溶性有機氮和微生物生物量氮含量呈極顯著正相關(P<0.01),與土壤pH呈極顯著負相關(P<0.01)。土壤AOA-amoA基因拷貝數與土壤pH呈極顯著正相關(P<0.01),與土壤有機碳、硝態氮和微生物生物量氮含量呈顯著負相關(P<0.05)。土壤nirK基因拷貝數與土壤pH和銨態氮含量呈極顯著正相關關系(P<0.01),與土壤有機碳、硝態氮、可溶性有機氮和微生物生物量氮含量呈顯著負相關關系(P<0.05)。AOB/AOA與土壤有機碳、硝態氮、可溶性有機氮和微生物生物量氮呈極顯著正相關(P<0.01),與pH呈極顯著負相關(P<0.01)。氨氧化/反硝化與土壤有機碳、硝態氮、可溶性有機氮和微生物生物量氮呈極顯著正相關(P<0.01),與pH和銨態氮含量呈顯著負相關(P<0.05)。
3 討論
3.1 土壤固氮功能基因對氮沉降響應的綜合分析
土壤固氮微生物是一類重要的氮轉化功能微生物,其能夠將空氣中的氮還原成氨供給植物吸收利用。固氮微生物數量與輸入土壤的外源氮素密切相關,施氮量過高會降低固氮菌的豐度并抑制固氮酶的活性,并對固氮菌群落組成產生顯著影響[26]。本研究中,隨著氮添加水平的升高,nifH豐度表現為先升高后降低趨勢,N300處理降低了nifH基因相對豐度,其他6個氮添加處理均高于或顯著高于對照N0。即低于200 kg N·hm-2·a-1時更有利于固氮菌的生長,而高于300 kg N·hm-2·a-1時反而抑制其生長。這與劉彩霞等[27]得出的氮沉降(銨態氮)高于60 kg N·hm-2·a-1抑制杉木林固氮功能微生物生長結論一致。Zhang等[7]研究也表明,高濃度無機氮添加抑制固氮菌生長。前人研究認為,氮肥輸入降低固氮菌的豐度是由于土壤酸化或高氮含量造成的[28]。
土壤固氮菌豐度與土壤有機碳含量、pH、有效磷含量等其他化學因子密切相關。本研究相關分析表明,nifH基因豐度與土壤有機碳含量呈顯著負相關性。由于生物固氮過程需要消耗大量能量,而該過程在很大程度上依賴于土壤中的有機碳含量。高氮添加雖然提高了土壤有機碳含量,但高氮添加導致高的速效氮含量反而會抑制土壤的生物固氮過程。本研究表明,nifH基因豐度與銨態氮含量呈極顯著正相關,與土壤硝態氮含量呈顯著負相關。表明氮添加量增加引起土壤可利用性氮含量變化是引起固氮功能基因豐度發生變化的重要驅動因素。Wang等[29]研究表明,長期施肥引起小麥生長季土壤銨態氮含量改變,顯著影響固氮菌豐度,不利于土壤微生物的固氮功能。本研究中,低氮添加促進土壤的生物固氮過程,但過高氮添加反而會抑制土壤的生物固氮過程。表明未來連續高氮沉降可能不利于貝加爾針茅草原土壤微生物的固氮功能。
3.2土壤氨氧化功能基因對氮沉降響應的綜合分析
硝化作用是氮素轉化的關鍵過程,由含氨單加氧酶基因(amoA)、氨氧化細菌(AOB)和氨氧化古菌(AOA)共同驅動氨氧化過程是硝化作用的限速步驟,其數量和結構組成變化可反映土壤氮素變化[30]。目前氮肥或氮添加對土壤硝化作用的影響研究大多是針對這一步驟開展的。一般情況下,氨是微生物進行硝化作用的直接底物,酸性土壤中銨根離子分解處理的氨較少,從而抑制AOB生長[31]。本研究,低氮添加(N15、N30和N50)對氨氧化細菌AOB相對豐度無顯著影響,而高氮添加顯著提高了氨氧化細菌豐度。說明連續6 a高氮添加,促進了AOB的生長繁殖。這與Li等[32]對內蒙古草原10 a氮添加試驗AOB基因豐度變化結果一致。Ai等[33]在對小麥玉米輪作潮土研究表明,氮肥添加可顯著提高AOB的數量。其可能原因是土壤中存在對NH+4有較高親和力的未知的AOB[34]或者是某些AOB的生長不以NH+4作為唯一底物[35]。與AOB豐度變化情況不同,除N30處理與對照相比無顯著差異外,其余6種氮添加處理均降低了土壤氨氧化古菌基因豐度,說明長期氮沉降增加會導致土壤氨氧化古菌豐度降低。長期施氮肥導致南方紅壤中氨氧化古菌豐度降低[36],這表明盡管草原暗栗鈣土和南方紅壤在pH、有機質和碳氮水平上存在較大差異,但長期氮添加和長期施用氮肥對這兩種土壤中氨氧化古菌的影響存在一致性。在本研究中,各氮添加處理土壤AOB-amoA基因拷貝數比AOA-amoA基因拷貝數高一個數量級,可見本試驗環境下更有利于AOB生長。這與Yao等[37]在茶園酸性土壤AOA主導氨氧化作用的研究結果不一致。前人研究表明,AOB和AOA生長對土壤含氮量響應不同,AOB對高氮環境較為適應,而AOA傾向于低氮環境[38]。本試驗中高氮添加處理土壤硝態氮含量顯著高于低氮添加和對照處理,從而促進了AOB生長。但對于這一原因推測的合理性還需要進一步深入研究。
大量研究表明,多種土壤環境因子,包括pH、溫度、土壤類型和利用方式等都會影響氨氧化微生物的群落組成和數量,從而使氮添加對土壤氨氧化微生物影響存在很大不確定性。本研究氨氧化細菌和氨氧化古菌與土壤理化因子的相關分析表明,影響氨氧化功能基因相對豐度的土壤理化因子為pH、有機碳、硝態氮和微生物生物量氮含量。這與Hayden等[39]對澳大利亞農業與草地氮轉化功能基因驅動因子研究結果一致。相關分析表明,AOB基因拷貝數與土壤有機碳含量呈極顯著正相關,說明連續6 a氮添加提高了貝加爾針茅草原土壤有機碳含量是促進氨氧化細菌生長繁殖的重要因素。一些研究表明,氨氧化細菌適合在氮素含量豐富的土壤中生存,且其數量與土壤pH呈顯著負相關[14,40]。本研究中,氨氧化細菌基因拷貝數與土壤pH呈極顯著負相關。這與其研究結果一致。原因可能是,隨著氮添加水平的增加,土壤pH降低,使得土壤中耐酸性AOB數量增多[41]。氨氧化古菌與土壤化學因子的相關性分析表明,氨氧化古菌基因拷貝數與土壤pH呈極顯著正相關,說明貝加爾針茅草原土壤中AOA基因豐度隨著pH降低而顯著降低。高氮添加增加了土壤中硝態氮含量,相應的AOB基因拷貝數增加,而AOA基因拷貝數則減少,AOB/AOA比值提高,且隨著氮添加水平的增加,提高的幅度明顯增加。表明連續6 a高氮添加,改變了氨氧化微生物相對豐度,氨氧化微生物對氨氧化的相對貢獻也可能發生改變。但氨氧化微生物對氨氧化作用的相對貢獻率是否發生改變,還需采用同位素示蹤技術進一步研究。相關分析表明,AOA基因拷貝數與土壤硝態氮含量呈極顯著負相關關系,這與Di等[38]的AOA適合于低氮環境的研究結論一致。本研究中長期連續氮添加導致的土壤化學性質變化,尤其是土壤pH降低導致土壤中AOA豐度明顯降低,可能降低土壤中AOA的氨氧化功能,進一步影響貝加爾針茅草原土壤的硝化作用。
3.3 土壤反硝化功能基因對氮沉降響應的綜合分析
反硝化作用是將硝酸根還原為N2O或N2的過程,是由許多厭氧或兼性厭氧微生物參與的一系列酶催化過程。這些酶主要由功能基因硝酸還原酶narG/napA、亞硝酸還原酶nirK/nirS、一氧化氮還原酶norB和氧化亞氮還原酶nosZ編碼。不同反硝化功能基因型對環境因子的影響存在較大差異。其中,亞硝酸還原酶nirK/nirS催化亞硝酸鹽還原成NO是微生物驅動反硝化作用關鍵步驟。已有研究表明,nirK型反硝化細菌對施肥種類和施肥量都十分敏感[42]。連續6 a氮添加顯著影響了nirK相對豐度,N30、N50和N100處理顯著提高了nirK相對豐度,N150、N200和N300處理顯著降低了nirK相對豐度。表明,高氮添加(N150、N200和N300)對nirK豐度有明顯的抑制作用。
環境因子如土壤pH、溫度、有機碳、硝態氮含量是影響反硝化作用的重要因素。本研究相關分析表明,nirk相對豐度與土壤pH、有機碳、硝態氮、銨態氮、可溶性有機氮和微生物生物量氮含量均呈顯著相關性(表3)。Xie等[43]研究認為土壤有機碳和硝酸鹽含量是驅動青藏高原草甸反硝化功能基因nirK豐度的重要土壤因子。Jahangir等[44]研究表明,反硝化活性的高低與提供給細菌生長的有機質的質量和數量密切相關。土壤反硝化作用最適pH范圍為7.0~8.0,在此范圍內反硝化作用最好[45]。在本研究中,高氮添加(N150、N200和N300)處理pH顯著低于對照N0,降低了土壤pH,抑制了反硝化功能基因nirK豐度,并由此可能對反硝化作用產生影響。一般情況下,土壤AOB基因豐度增加會提高反硝化作物的底物硝酸鹽含量,進而促進反硝化微生物驅動的反硝化作用,造成氮素流失。但本研究中,高氮(N150、N200和N300)顯著降低了反硝化細菌nirK基因豐度,而硝態氮含量卻顯著提高。說明高氮添加驅動的土壤氨氧化作用高于驅動的土壤的反硝化作用。土壤nirK基因豐度與土壤硝態氮含量呈極顯著負相關驗證了這一觀點。
上述研究表明,貝加爾針茅草原土壤反硝化活性受土壤化學性質特別是pH、有機碳、硝態氮、銨態氮、可溶性有機氮和微生物生物量氮含量的顯著影響。此外,由于不同反硝化過程所涉及的酶不同,同時攜帶不同反硝化基因的微生物種群對環境因子響應的差異,因此,揭示不同氮添加水平對反硝化功能微生物基因豐度的影響,需要對不同類型的反硝化微生物開展深入研究。
4 結論
(1)隨著氮添加水平的增加,固氮細菌nifH基因豐度呈現先升高后降低趨勢。低于200 kg N·hm-2·a-1時促進固氮菌生長。影響固氮菌nifH主要環境因子是土壤有機碳、銨態氮、硝態氮和微生物生物量氮含量。
(2)高氮添加(N100、N150、N200和N300)提高了AOB基因豐度,降低了AOA基因豐度,并顯著提高了AOB/AOA,表明氮添加量增加可能使AOB在貝加爾針茅草原土壤硝化過程中占主導作用。影響AOB、AOA基因豐度的主要環境因子是土壤pH、有機碳、硝態氮和微生物生物量氮含量。
(3)高氮添加(N150、N200和N300)降低了反硝化細菌nirK基因豐度。氮添加量的增加促進了AOB主導的氨氧化過程,而反硝化微生物nirK豐度的降低提高了氨氧化產物的積累,繼而提高了土壤中的硝態氮含量。影響反硝化細菌nirK的主要環境因子是土壤pH、有機碳、硝態氮、銨態氮、可溶性有機氮和微生物生物量氮含量。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
