糖原合成激酶3β抑制劑通過p38負調節感染性炎性反應的研究*
2019-11-14 07:50:12盧建強梁培松王偉佳
國際檢驗醫學雜志 2019年21期
關鍵詞:水平
陳 康,傅 強,盧建強,梁培松,韓 慧,董 謙,王偉佳
(廣東省中山市人民醫院檢驗醫學中心,廣東中山 528403)
銅綠假單胞菌是一種條件致病菌,可引起醫院獲得性肺炎、角膜炎等多種機會感染[1-2]。盡管抗菌藥物可以殺傷細菌,卻不能抑制因感染而激發的過度免疫應答。因此,如何有效抑制過度免疫應答是預防免疫病理損傷和治療疾病的關鍵。糖原合成激酶3β(GSK3β)是組織器官中廣泛表達的絲氨酸/蘇氨酸蛋白激酶,可參與糖原合成,多種病理生理過程,如高血壓、糖尿病、炎性腸病等[3-5]。此外,還可參與多種病原體感染引起的免疫應答,如病毒、真菌等[6-7]。磷酸化是調節GSK3β活性最常見的方式,其中以失活型9位絲氨酸(Ser9)的磷酸化最為常見,當Ser9的磷酸化水平降低時,提示在該疾病中GSK3β活性增高,破壞細胞的穩態,導致細胞發生氧化應激和炎性反應。絲裂原活性蛋白激酶(MAPK)是調節炎性反應通路中的重要分子,GSK3β可與MAPK家族中成員包括p38、c-Jun氨基末端激酶(JNK)、細胞外調節蛋白激酶(ERK)協同調控細胞的炎性反應。GSK3β還可參與多種疾病的調控,被用作疾病治療的靶點。GSK3β的小分子抑制劑SB216763可抑制其活性,調節免疫應答[8],因此本研究旨在探討感染銅綠假單胞菌時,GSK3β抑制劑調節細胞因子分泌及免疫功能的作用及機制。
1 材料與方法
1.1試劑 GSK3β抑制劑SB216763、p38抑制劑SB203580、JNK抑制劑SP600125均購自美國MCE公司;p38、磷酸化p38(P-p38)、JNK、磷酸化JNK(P-JNK)、ERK、磷酸化ERK(P-ERK)、磷酸化GSK3β失活型9位絲氨酸[P-GSK3β(Ser9)]和β-actin的抗體均購自美國CST公司;Trizol、反轉錄試劑盒購自美國Invitrogen公司;異丙醇和無水乙醇購自中國廣州鼎國生物技術有限公司;實時熒光定量PCR(qPCR)檢測用SYBR Green試劑盒購自美國Bio-Rad公司。
1.2細胞培養 小鼠巨噬細胞購自美國菌種保藏中心,細胞培養于DMEM培養基中。將4×105個/mL 小鼠巨噬細胞鋪入12孔細胞培養板中,采用GSK3β抑制劑(10 μmol/L)、p38抑制劑(1 μmol/L)或JNK抑制劑(1 μmol/L)預處理細胞1 h,加入銅綠假單胞菌(MOI=1)刺激后,檢測蛋白或mRNA水平的表達。
1.3免疫印跡試驗(Western blot) 用冷的磷酸鹽緩沖液沖洗細胞培養板中的細胞2次,加入蛋白裂解液,收集細胞裂解碎片,冰上裂解20 min。12 000 r/min離心10 min,收集蛋白裂解上清,考馬斯亮藍法進行蛋白定量。接著進行聚丙烯酰胺凝膠電泳,并電轉至硝酸纖維素膜上。將膜用5%脫脂奶粉室溫孵育2 h,4 ℃過夜分別孵育一抗p38(1∶1 000)、P-p38(1∶1 000)、JNK(1∶1 000)、P-JNK(1∶1 000)、ERK(1∶1 000)、P-ERK(1∶1 000)、P-GSK3β(Ser 9)(1∶1 000)、β-actin(1∶5 000),室溫下孵育二抗1 h,采用電化學發光方法檢測蛋白水平。
1.4qPCR RNA抽提試劑法進行RNA提取,分光光度計測RNA濃度和純度,用于下一步實驗。將提取后的RNA(1 μg)轉錄為cDNA,詳細步驟如下:RNA 1 μg,oligo(dT)1 μL,補DEPC水至體積12 μL,混勻;65 ℃,5 min;每孔加入5×反應液4 μL,20 U/μL RNA酶抑制劑1 μL,10 mmol/L dNTP 2 μL,200 U/μL反轉錄酶1 μL,至總體積20 μL,混勻;42 ℃,60 min,70 ℃,5 min終止反應。反轉錄的cDNA保存備用。應用SYBR Green試劑盒進行擴增,詳述如下:SYBR Green(2×) 10 μL,引物上下游混合液(5 μmol/L) 1.6 μL,cDNA模板2 μL,滅菌H2O補齊總體積20 μL充分混勻;按照兩步法PCR擴增程序,第1步,95 ℃ 30 s,1個循環;第2步,95 ℃ 5 s,60 ℃ 30 s,40個循環PCR反應。反應結束后確認qPCR的擴增曲線和溶解曲線,得到循環閾值(Ct)值,采用2-ΔΔCT法進行相對定量。IL-10、IL-6和β-actin的引物序列見表1。

表1 引物序列
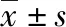
2 結 果
2.1GSK3β參與到銅綠假單胞菌感染中 實驗結果顯示,在銅綠假單胞菌感染小鼠巨噬細胞6 h后,P-GSK3β(Ser9)水平增高,然后隨感染時間的增加,水平下調。這些結果提示,GSK3β參與到銅綠假單胞菌感染中。見圖1。
2.2GSK3β抑制劑調節細胞因子的表達 實驗結果顯示,在銅綠假單胞菌感染巨噬細胞后,GSK3β抑制劑SB216763可促進細胞因子IL-10水平的表達,同時抑制細胞因子IL-6水平的表達。結果提示抑制GSK3β可在銅綠假單胞菌感染中發揮免疫負調的作用。見圖2。
2.3GSK3β抑制劑調節MAPK家族成員的磷酸化水平 實驗結果顯示,在銅綠假單胞菌感染后15 min、30 min和2 h,GSK3β抑制劑可上調p38和JNK的磷酸化水平,而對ERK的磷酸化水平無調節作用。實驗結果同時發現,細菌感染前后,GSK3β抑制劑對p38、JNK和ERK的蛋白水平無調節作用。見圖3。

注:A表示P-GSK3β(Ser9)蛋白表達水平隨感染時間的變化情況;B表示P-GSK3β(Ser9)蛋白表達定量隨感染時間的變化情況;***P<0.001
圖1 GSK3β參與到銅綠假單胞菌感染中

注:A表示IL-10基因水平表達情況;B表示IL-6基因水平表達情況;*P<0.05;***P<0.001
圖2 GSK3β抑制劑通過p38調節細胞因子的表達

圖3 GSK3β抑制劑調節MAPK的磷酸化水平
2.4GSK3β抑制劑通過p38調節細胞因子的表達 實驗結果顯示,GSK3β抑制劑可上調細胞因子IL-10的表達,在應用GSK3β抑制劑的同時,應用p38抑制劑可抑制細胞因子IL-10的表達,應用JNK的抑制劑不可抑制IL-10的表達;GSK3β抑制劑可下調細胞因子IL-6的表達,在應用GSK3β抑制劑的同時,應用p38抑制劑可上調細胞因子IL-10的表達,而同時應用JNK的抑制劑不可上調IL-10的表達。這些結果提示,GSK3β抑制劑通過p38 MAPK信號通路調節細胞因子的表達。見圖2。
3 討 論
本研究發現,在感染銅綠假單胞菌時,GSK3β活性增高;GSK3β抑制劑可通過p38調節細胞因子的分泌,負調節免疫反應。
GSK3β參與到多種疾病的致病過程和病原體感染中[3-7],因此被用作疾病治療的靶點。研究報道,GSK3β的抑制劑可發揮免疫調控的作用[8]。MARTIN等[9]報道,抑制GSK3β可抑制多種配體激活的炎性反應,抑制促炎性細胞因子的表達,同時促進抑炎性細胞因子IL-10的表達。在敗血癥休克中,抑制GSK3β可上調抑炎性因子IL-10的表達[10]。然而,SHEN等[11]報道,GSK3β抑制劑SB216763可在心肌細胞中促進脂多糖誘導腫瘤壞死因子的表達。本課題組研究發現,GSK3β抑制劑SB216763可促進抑炎性細胞因子IL-10的表達,同時抑制促炎性細胞因子IL-6的表達,在銅綠假單胞菌感染中負調節免疫應答。
GSK3β可參與多個信號通路的調控,可通過抑制NF-κB信號通路或上調環磷腺苷效應元件結合蛋白抑制免疫應答[12]。NOH等[10]報道,GSK3β抑制劑可通過抑制ERK的活性上調抑炎性因子IL-10的表達。本課題組研究發現,GSK3β抑制劑可上調p38和JNK的磷酸化水平,并通過p38 MAPK信號通路調節細胞因子IL-10和IL-6的表達,發揮免疫負調節作用。
4 結 論
在感染銅綠假單胞菌后,GSK3β的活性增高;應用GSK3β的抑制劑SB216763可調節細胞因子的表達,負調節免疫應答;進一步實驗發現,GSK3β抑制劑通過p38 MAPK信號通路調節細胞因子的表達。這些研究提示在銅綠假單胞菌感染中GSK3β可作為免疫調控的靶點。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
