氮磷肥配施對西北羊茅開花期葉片光合特性日變化的影響
2019-11-18 06:03:38石正海劉文輝張永超秦燕魏小星
草業學報 2019年11期
關鍵詞:效率
石正海,劉文輝,張永超,秦燕,魏小星
(青海省青藏高原優良牧草種質資源利用重點實驗室,青海省畜牧獸醫科學院,青海大學,青海 西寧 810016)
土壤氮(N)、磷(P)等營養元素的有效供應有利于植物生長[1-2]。N是參與光合作用相關酶的主要組成元素,而這類酶主要在核糖體(主要元素為P)合成且P直接參與碳同化和磷酸化過程[3-4]。由于氮沉降,多數生態系統目前由N限制轉向N、P共同限制[5-6]。青藏高原地區氮沉降速率為8.7~13.8 kg·hm-2·yr-1,高于全國平均水平[7],不同植物對氮沉降響應機制不同[8],因此有必要對高寒植物的氮磷響應機制展開研究。人工草地常添加氮肥、磷肥以補充土壤有效N、P,植物對N、P吸收和利用相互依賴[9],Marklein等[10]發現施N增強植物磷酸酶活性,從而降低P限制對植物的影響。通過凈光合速率、氣孔導度、蒸騰速率及水分利用效率等氣體交換參數可反映施肥對植物光合作用的影響[2,4]。氮素添加不僅提高羊草(Leymuschinensis)凈光合速率,還延長其峰值持續時間[11]。凈光合速率的變化是氣孔和非氣孔因素的共同作用結果,Farquhar等[12]認為通過氣孔導度和胞間CO2濃度兩個參數變化,可判斷光合速率是否受氣孔限制。葉綠素熒光參數真實反映植物內在的生理狀態,以彌補氣體交換參數的不足。氮肥提高早熟禾(Poapratensis)和剪股穎(Agrostismatsumurae)葉片PSⅡ活性[10,13],磷肥能減少葉片熱耗散和提高光能利用能力[3,14]。葉片將吸收的光能分配給相互競爭的光化學反應、熒光耗散和熱耗散3個方面[15],其中光化學反應的比例大小體現植物對光能的利用率。
西北羊茅(Festucakryloviana)是高寒地區優良牧草,具有較小的莖葉比,且適口性好[16]。開花期是植物由營養生長到生殖生長的轉化時期,也是牧草營養價值與生物量最佳耦合時期[17-19]。植物葉片作為吸收光能的主要器官,其光合能力直接影響生物量積累。植物“午休”現象體現光合有效輻射對光合特性影響[20],研究西北羊茅光合特性的日變化,有助于確定合理的利用時間,而關于西北羊茅光合特性的報道尚無。因此本研究通過對開花期西北羊茅的氣體交換參數和熒光參數研究,以揭示西北羊茅對光照強度的適應機理及對氮肥和磷肥添加的響應,為西北羊茅的利用提供理論依據。
1 材料與方法
1.1 研究地概況
試驗地位于青海省海北州草原站多年生牧草種質資源圃(36°59.36′ E、100°52.85′ N),海拔3156 m;年平均氣溫0.5 ℃,年降水量約369.1 mm,且多集中在7-9月;無絕對無霜期,平均無霜期93 d。土壤為栗鈣土,有機質含量為3%~4%,旱作。
1.2 試驗設計
于2018年在建植第4年的西北羊茅種子田(2015年建植,播量22.5 kg·hm-2,行距30 cm)進行。為保障土壤能供給足量的N、P以滿足西北羊茅的生長需求,試驗設計2個施氮水平(0和60 kg·hm-2;記為N0和N60)和3個施磷水平(0、60和90 kg·hm-2;記為P0、P60和P90),共計6個處理組合[N0P0(CK)、N0P60、N0P90、N60P0、N60P60、N60P90],隨機區組排列,3次重復,小區面積(3×5) m2。氮肥(尿素,含N 46%)和磷肥(過磷酸鈣,含P2O512%)混勻于西北羊茅返青期(5月中旬)在行間開溝(溝深20 cm)一次性施入。
1.3 生理生態指標的測定
1.3.1光合輻射日動態測定 于2018年7月18-22日晴朗天氣的8:00-19:00時,采用LI-6400便攜式光合測定儀(LI-COR,Lincoln,USA)內置光量子傳感器每隔1 h測定光合有效輻射值(photosynthetically active radiation, PAR)。
1.3.2氣體交換參數測定 于2018年7月18-22日(開花期)晴朗無風的9:00-11:00,采用LI-6400便攜式光合測定儀(LI-COR,Lincoln,USA)測定西北羊茅葉片氣體交換參數。每小區選擇5株生長良好的莖生葉片,測定過程使用內置紅藍光源,為模擬光照強度的日動態,PAR設定7個水平(分別為0、400、800、1000、1200、1600和2000 μmol·m-2·s-1),進行光合-光強響應測定,且控制葉室溫度25 ℃,二氧化碳濃度400 μmol·mol-1,流速500 μmol·s-1。測定凈光合速率(net photosynthetic rate,Pn)、蒸騰速率(transpiration rate,Tr)、氣孔導度(stomatal conductance,Gs)和胞間CO2濃度(intercellular CO2concentration,Ci)等氣體交換參數,并計算氣孔限制值(stomatal limited value,Ls),Ls=1-Ci/Ca(Ca為大氣中CO2濃度)和水分利用效率(water use efficiency, WUE),WUE=Pn/Tr。
1.3.3葉綠素熒光參數測定 測定氣體交換參數后,采用LI-6400便攜式光合測定儀繼續測定西北羊茅葉片葉綠素熒光參數。將葉片在錫箔紙遮光處理下暗適應30 min后,測定初始熒光(minimal fluorescence,Fo)、最大熒光(maximal fluorescence,Fm)。打開光合儀內置紅藍光源,PAR設定同上(不設0 μmol·m-2·s-1),在光下光適應穩定后,測定光適應下最小熒光(Fo′)、最大熒光(Fm′)、穩定熒光(fluorescence instable,Fs)等熒光參數,以計算PSⅡ最大光化學效率(maximum photochemical efficiency of PSⅡ,Fv/Fm),Fv/Fm=(Fm-Fo)/Fm、PSⅡ有效光化學效率(effective photochemical efficiency of PSⅡ,Fv′/Fm′),Fv′/Fm′=(Fm′-Fo′)/Fm′、PSⅡ實際光化學效率(actual photochemical efficiency of PSⅡ,ΦPSⅡ),ΦPSⅡ=(Fm′-Fs)/Fm′、光化學猝滅系數(photochemical quenching coefficient,qP),qP=(Fm′-Fs)/(Fm′-Fo′)、非光化學猝滅系數(non-photochemical quenching coefficient, NPQ),NPQ=Fm/Fm′-1和電子傳遞速率(electron transport rate, ETR),ETR=PAR×ΦPSⅡ×0.87×0.5。并參照Demmig等[21]的方法計算光合系統吸收的光能用于光化學反應的比例(proportion of photosynthetic electron transport, P),P=Fv′/Fm′×qP,用于天線熱耗散的比例(versus thermal dissipation, D),D=1-Fv′/Fm′和用于反應中心非光化學反應耗散的比例(excess, E),E=Fv′/Fm′×(1-qP)。
1.4 數據處理
用Excel 2013對數據初步整理,SPSS 19.0軟件進行數據統計和分析,Origin 2018作圖。為了解在不同施肥處理和光合有效輻射間是否差異顯著,對各處理的氣體交換參數及葉綠素熒光參數等數據進行多因素方差分析,并采用Duncan法進行多重比較(P<0.05)。
2 結果與分析
2.1 光合有效輻射日動態
光合有效輻射的日動態為先增后降,通過對其擬合得開口向下“拋物線”,擬合方程為Y=-64.971t2+1736.954t-9600.744 (R2=0.9482,P<0.001)(圖1)。

圖1 光合有效輻射的日動態Fig.1 Dynamic changes of photosynthetically active radiation in days
PAR在11:00-16:00時均高于1500 μmol·m-2·s-1,在下午13:00時甚至超過2000 μmol·m-2·s-1;變化最快的兩個時間段為上午9:00-10:00和下午17:00-18:00,其值變化范圍為400~1200 μmol·m-2·s-1。
2.2 施肥和光合有效輻射對凈光合速率的影響
施肥和光合有效輻射均極顯著(P<0.001)影響凈光合速率(表1)。結合表1、圖 2A得,光合有效輻射為0 μmol·m-2·s-1時西北羊茅僅呼吸消耗光合同化物,此時凈光合速率為細胞呼吸速率,均大于1.37 μmol CO2·m-2·s-1。 凈光合速率隨光合有效輻射增強先增加后降低,在光合有效輻射0~400 μmol·m-2·s-1段快速增加,在800~1200 μmol·m-2·s-1段相對穩定,平均達6.81 μmol CO2·m-2·s-1,而后開始下降,尤其在1600~2000 μmol·m-2·s-1段快速下降。最適合CK處理的光合有效輻射在800 μmol·m-2·s-1左右,施肥不僅延后最適光合有效輻射值還提高凈光合速率;各施肥處理凈光合速率均為CK的1.4倍以上。施氮和施磷提升凈光合速率幅度分別達93%和56%,另外,氮肥和磷肥配施有效抑制了強光照下凈光合速率的減小,在光合有效輻射值2000 μmol·m-2·s-1時,其值為CK的4倍以上。
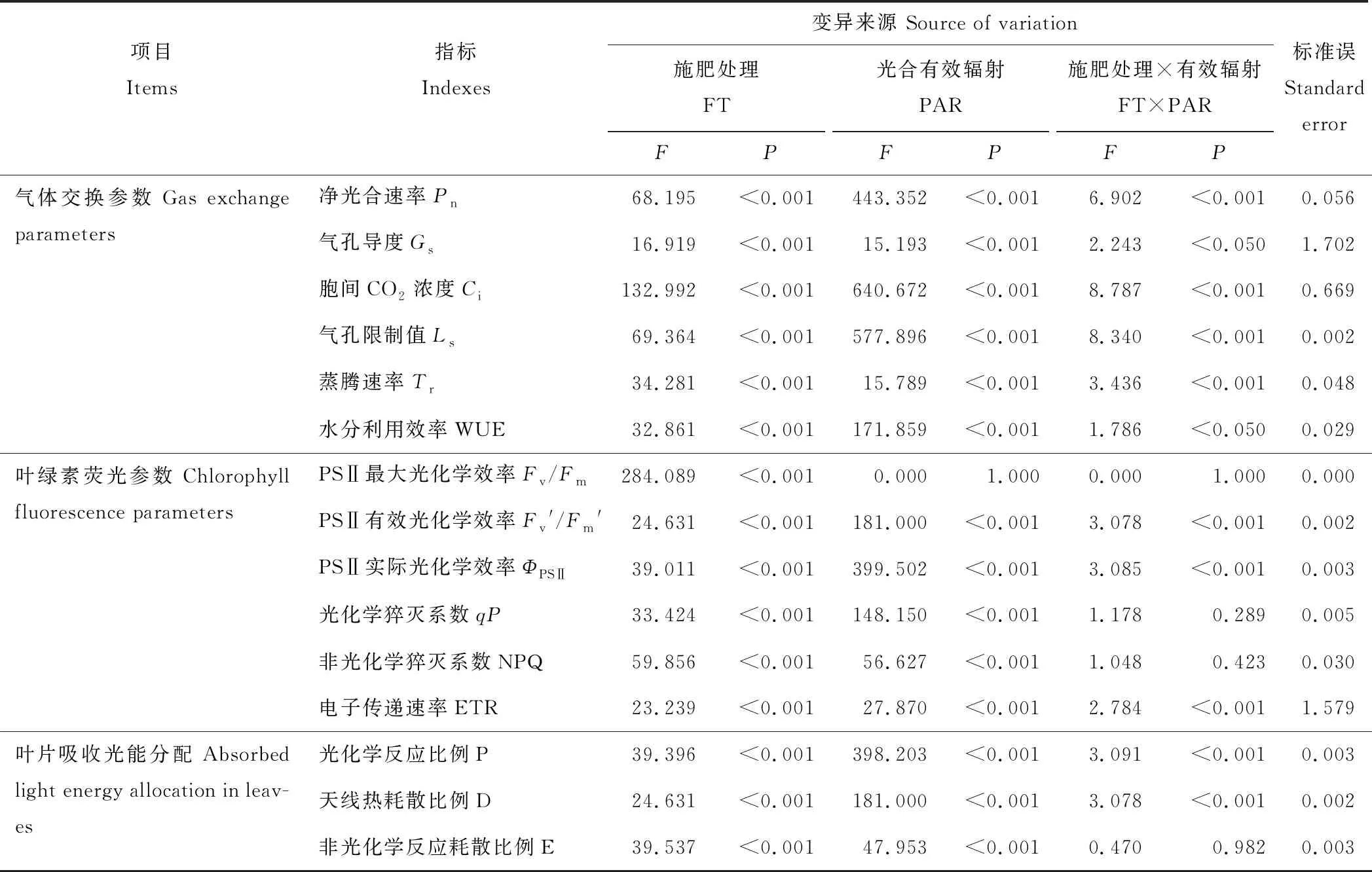
表1 施肥和光合有效輻射變化對光合特性的影響Table 1 Effect of fertilization and photosynthetically active radiation on photosynthetic characteristics
注:表中數據表示F值和P值。FT:施肥處理;PAR:光合有效輻射;Pn:凈光合速率;Gs:氣孔導度;Ci:胞間CO2濃度;Ls:氣孔限制值;Tr:蒸騰速率;WUE:水分利用效率;Fv/Fm:PSⅡ最大光化學效率;Fv′/Fm′:PSⅡ有效光化學效率;ΦPSⅡ:PSⅡ實際光化學效率;qP:光化學猝滅系數;NPQ:非光化學猝滅系數;ETR:電子傳遞速率;P:光化學反應比例;D:天線熱耗散比例;E:非光化學反應耗散比例。下同。
Note: Data are shownFandPvalue. FT: fertilization treatment; PAR: photosynthetically active radiation;Pn: net photosynthetic rate;Gs: stomatal conductance;Ci: intercellular CO2concentration;Ls: stomatal limited value;Tr: transpiration rate; WUE: water use efficiency;Fv/Fm: maximum photochemical efficiency of PSⅡ;Fv′/Fm′: effective photochemical efficiency of PSⅡ;ΦPSⅡ: actual photochemical efficiency of PSⅡ;qP: photochemical quenching coefficient; NPQ: non-photochemical quenching coefficient; ETR: electron transport rate; P: proportion of photosynthetic electron transport; D: versus thermal dissipation; E: excess. The same below.
2.3 施肥和光合有效輻射對葉片氣體交換參數的影響
胞間CO2濃度、氣孔限制值、氣孔導度、蒸騰速率及水分利用效率均受施肥和光合有效輻射極顯著影響(P<0.001),通過F值可得光合有效輻射對氣體交換參數的影響大于施肥措施(表1)。進一步分析(圖2B~F),隨光合有效輻射增強,西北羊茅的氣孔導度、氣孔限制值、蒸騰速率和水分利用效率均先增后降,胞間CO2濃度則相反,但各參數變化速率與凈光合速率類似,在光合有效輻射800~1200 μmol·m-2·s-1間均出現極值。不同處理間,CK處理由于較小的氣孔導度和較大的胞間CO2濃度,進而使氣孔限制值較小,從而維持低水平的凈光合速率;而低的凈光合速率和蒸騰速率反而使葉片水分利用效率較高。施肥處理的各項氣體交換參數與CK差異顯著(水分利用效率除外);胞間CO2濃度在施肥后平均降低10%,且隨施磷量增加進一步降低,其值在N0P90處理最低,僅為259.037 μmol CO2·mol-1,氣孔限制值變化與胞間CO2濃度相反。氣孔導度在氮肥和磷肥配施處理有較大值;光合有效輻射值1200~2000 μmol·m-2·s-1時,其值在僅施磷處理降低幅度高達48%。施肥增加了蒸騰速率,但各處理間無統一規律,相對于氮肥或磷肥單施,其配施在一定程度上減小水分的散失,蒸騰速率降低幅度達22%,進而提高水分利用效率。綜合各氣體交換參數,施肥后西北羊茅通過增加氣孔導度促進氣體交換以保持高的凈光合速率,并維持高效的水分利用效率。
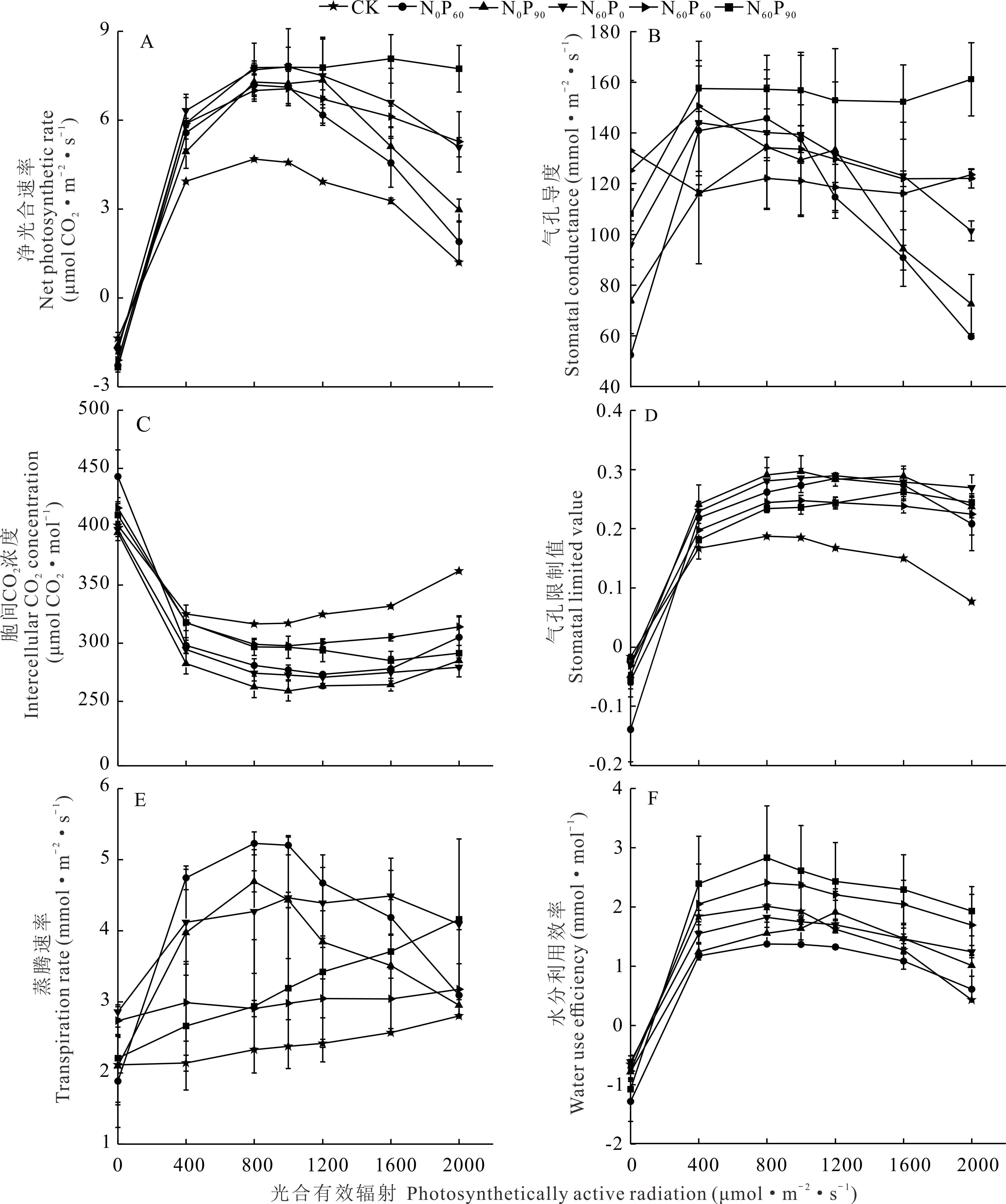
圖2 施肥和光合有效輻射梯度對氣體交換參數的影響Fig.2 Effect of fertilization and photosynthetically active radiation on gas exchange parameters
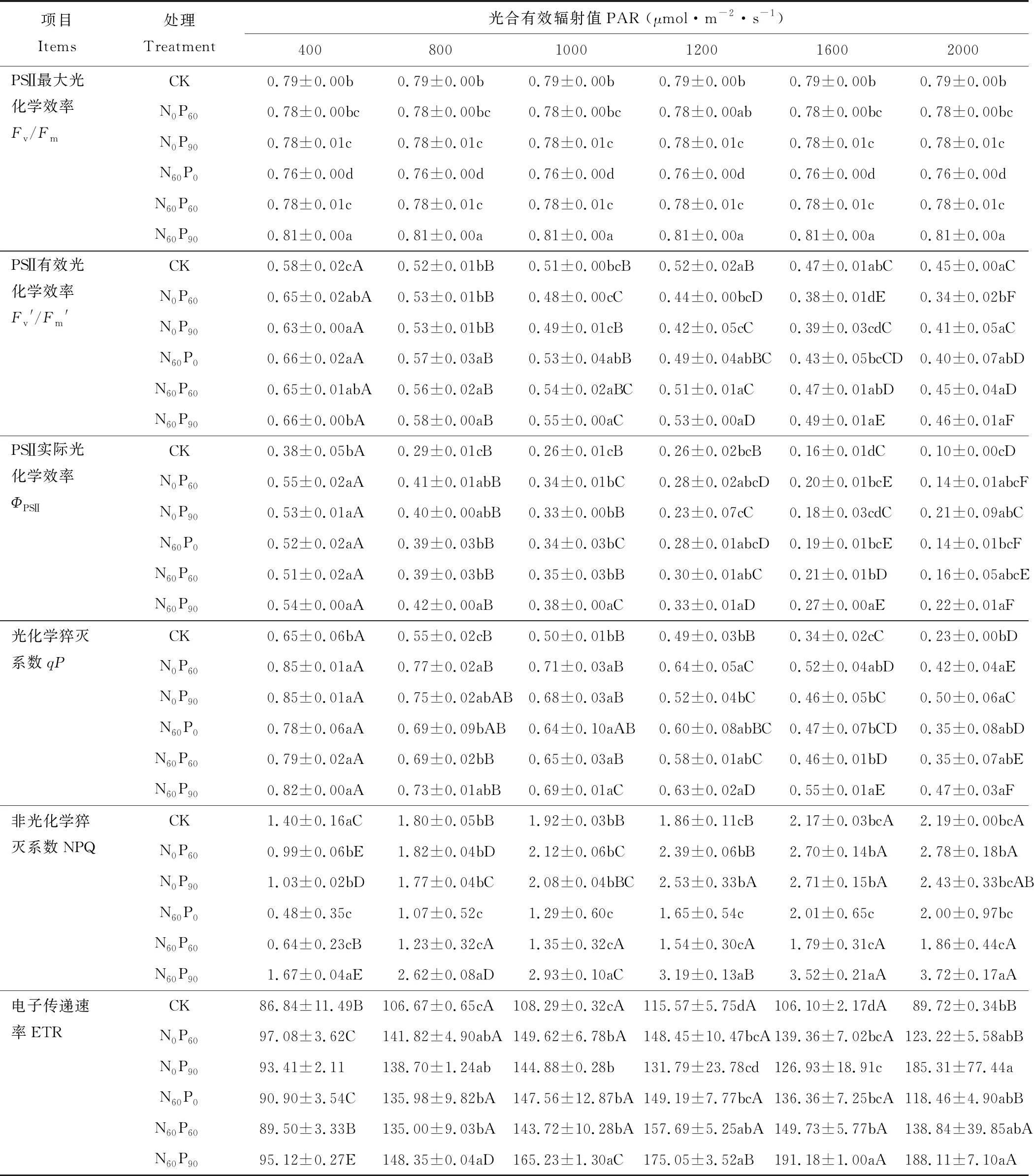
項目Items處理Treatment光合有效輻射值PAR (μmol·m-2·s-1)4008001000120016002000PSⅡ最大光化學效率Fv/FmCK0.79±0.00b0.79±0.00b0.79±0.00b0.79±0.00b0.79±0.00b0.79±0.00bN0P600.78±0.00bc0.78±0.00bc0.78±0.00bc0.78±0.00ab0.78±0.00bc0.78±0.00bcN0P900.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01cN60P00.76±0.00d0.76±0.00d0.76±0.00d0.76±0.00d0.76±0.00d0.76±0.00dN60P600.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01c0.78±0.01cN60P900.81±0.00a0.81±0.00a0.81±0.00a0.81±0.00a0.81±0.00a0.81±0.00aPSⅡ有效光化學效率Fv'/Fm'CK0.58±0.02cA0.52±0.01bB0.51±0.00bcB0.52±0.02aB0.47±0.01abC0.45±0.00aCN0P600.65±0.02abA0.53±0.01bB0.48±0.00cC0.44±0.00bcD0.38±0.01dE0.34±0.02bFN0P900.63±0.00aA0.53±0.01bB0.49±0.01cB0.42±0.05cC0.39±0.03cdC0.41±0.05aCN60P00.66±0.02aA0.57±0.03aB0.53±0.04abB0.49±0.04abBC0.43±0.05bcCD0.40±0.07abDN60P600.65±0.01abA0.56±0.02aB0.54±0.02aBC0.51±0.01aC0.47±0.01abD0.45±0.04aDN60P900.66±0.00bA0.58±0.00aB0.55±0.00aC0.53±0.00aD0.49±0.01aE0.46±0.01aFPSⅡ實際光化學效率ΦPSⅡCK0.38±0.05bA0.29±0.01cB0.26±0.01cB0.26±0.02bcB0.16±0.01dC0.10±0.00cDN0P600.55±0.02aA0.41±0.01abB0.34±0.01bC0.28±0.02abcD0.20±0.01bcE0.14±0.01abcFN0P900.53±0.01aA0.40±0.00abB0.33±0.00bB0.23±0.07cC0.18±0.03cdC0.21±0.09abCN60P00.52±0.02aA0.39±0.03bB0.34±0.03bC0.28±0.01abcD0.19±0.01bcE0.14±0.01bcFN60P600.51±0.02aA0.39±0.03bB0.35±0.03bB0.30±0.01abC0.21±0.01bD0.16±0.05abcEN60P900.54±0.00aA0.42±0.00aB0.38±0.00aC0.33±0.01aD0.27±0.00aE0.22±0.01aF光化學猝滅系數qPCK0.65±0.06bA0.55±0.02cB0.50±0.01bB0.49±0.03bB0.34±0.02cC0.23±0.00bDN0P600.85±0.01aA0.77±0.02aB0.71±0.03aB0.64±0.05aC0.52±0.04abD0.42±0.04aEN0P900.85±0.01aA0.75±0.02abAB0.68±0.03aB0.52±0.04bC0.46±0.05bC0.50±0.06aCN60P00.78±0.06aA0.69±0.09bAB0.64±0.10aAB0.60±0.08abBC0.47±0.07bCD0.35±0.08abDN60P600.79±0.02aA0.69±0.02bB0.65±0.03aB0.58±0.01abC0.46±0.01bD0.35±0.07abEN60P900.82±0.00aA0.73±0.01abB0.69±0.01aC0.63±0.02aD0.55±0.01aE0.47±0.03aF非光化學猝滅系數NPQCK1.40±0.16aC1.80±0.05bB1.92±0.03bB1.86±0.11cB2.17±0.03bcA2.19±0.00bcAN0P600.99±0.06bE1.82±0.04bD2.12±0.06bC2.39±0.06bB2.70±0.14bA2.78±0.18bAN0P901.03±0.02bD1.77±0.04bC2.08±0.04bBC2.53±0.33bA2.71±0.15bA2.43±0.33bcABN60P00.48±0.35c1.07±0.52c1.29±0.60c1.65±0.54c2.01±0.65c2.00±0.97bcN60P600.64±0.23cB1.23±0.32cA1.35±0.32cA1.54±0.30cA1.79±0.31cA1.86±0.44cAN60P901.67±0.04aE2.62±0.08aD2.93±0.10aC3.19±0.13aB3.52±0.21aA3.72±0.17aA電子傳遞速率ETRCK86.84±11.49B106.67±0.65cA108.29±0.32cA115.57±5.75dA106.10±2.17dA89.72±0.34bBN0P6097.08±3.62C141.82±4.90abA149.62±6.78bA148.45±10.47bcA139.36±7.02bcA123.22±5.58abBN0P9093.41±2.11138.70±1.24ab144.88±0.28b131.79±23.78cd126.93±18.91c185.31±77.44aN60P090.90±3.54C135.98±9.82bA147.56±12.87bA149.19±7.77bcA136.36±7.25bcA118.46±4.90abBN60P6089.50±3.33B135.00±9.03bA143.72±10.28bA157.69±5.25abA149.73±5.77bA138.84±39.85abAN60P9095.12±0.27E148.35±0.04aD165.23±1.30aC175.05±3.52aB191.18±1.00aA188.11±7.10aA
注:不同小寫字母表示同一光合有效輻射,不同施肥處理間差異顯著(P<0.05);不同大寫字母表示同一施肥處理,不同光合有效輻射間差異顯著(P<0.05),無大寫或小寫字母表示差異不顯著(P>0.05)。下同。
Note: The different lowercase letters indicate significant differences (P<0.05) among the same photosynthetically active radiation and different fertilization treatment, the different capital letters indicate significant differences (P<0.05) among the same fertilization treatment and different photosynthetically active radiation, no capital or lowercase letters mean no significant differences (P>0.05). The same below.
2.4 施肥和光合有效輻射對葉綠素熒光參數的影響
葉綠素熒光參數常用作探測植物葉片光合能力。由表1,西北羊茅葉片PSⅡ實際光化學效率、光化學猝滅系數、非光化學猝滅系數和電子傳遞速率均受施肥和光合有效輻射極顯著(P<0.001)影響;但光化學猝滅系數和非光化學猝滅系數受施肥和光合有效輻射交互影響不顯著(P>0.05);PSⅡ最大光化學效率僅受施肥極顯著(P<0.001)影響。進一步分析(表2),隨光合有效輻射增強,PSⅡ最大光化學效率變化不顯著;而PSⅡ有效光化學效率顯著減小,其值在400 μmol·m-2·s-1最大,均大于0.58。PSⅡ實際光化學效率與PSⅡ有效光化學效率保持一致,在2000 μmol·m-2·s-1時降低幅度大于60%,其中在1200~2000 μmol·m-2·s-1降低幅度最大。光化學猝滅系數和PSⅡ實際光化學效率類似,在400 μmol·m-2·s-1是2000 μmol·m-2·s-1的1.8~2.8倍。隨光合有效輻射增強,非光化學猝滅系數增大2.7倍;電子傳遞速率則先增加后降低,在1000 μmol·m-2·s-1有最大值,但在強光下電子傳遞速率仍高于弱光。PSⅡ最大光化學效率在各施肥處理間差異顯著,但無統一規律,僅在N60P90處理高于對照(0.79)2%;其中以單施N60P0處理最低,降幅高達3.7%。施肥顯著提高弱光下PSⅡ有效光化學效率,在強光下各處理間的差異不顯著,但N60P90處理均有較大值。施肥顯著增加PSⅡ實際光化學效率,尤其N60P90處理,在光合有效輻射2000 μmol·m-2·s-1時為對照的2.2倍。光化學猝滅系數在N0P60和N60P90處理均有較大值。非光化學猝滅系數在N60P0和N60P60處理較小,較對照降低幅度達48%,在N60P90處理均最大。施肥提高電子傳遞速率,尤其N60P90處理(光合有效輻射為400 μmol·m-2·s-1時除外)顯著高于其他施肥處理。
2.5 施肥和光合有效輻射對光能分配影響
施肥和光合有效輻射均極顯著(P<0.001)影響葉片對吸收光能的分配,其中施肥和光合有效輻射的交互對非光化學反應耗散比例影響不顯著(P>0.05) (表1)。光合有效輻射為 400 μmol·m-2·s-1時,西北羊茅將吸收光能用于光化學反應、天線熱耗散和非光化學反應耗散的比例分別占51%、36%和13%;隨光合有效輻射增強,西北羊茅葉片用于光化學反應比例減少,天線熱耗散和非光化學反應耗散的比例增加;在2000 μmol·m-2·s-1時變化幅度分別達60%、55%和78%,主要變化在400~800 μmol·m-2·s-1和1200 μmol·m-2·s-1后(表3)。單施磷肥處理較CK處理,在同一光合有效輻射下用于光化學反應的比例顯著提高,用于天線熱耗散的比例和非光化學反應耗散的比例均顯著降低,但在施磷60和90 kg·hm-2水平差異不顯著。在氮肥和磷肥配施處理下,用于光化學反應比例較對照均大于30%,在光合輻射值大于1600 μmol·m-2·s-1后更是超過72%;在低光合有效輻射時用于天線熱耗散的比例低于對照10%以上,而在高光合有效輻射時與對照無顯著差異;非光化學反應耗散方面,各光合有效輻射均降低26%以上。
3 討論
3.1 施肥和光合輻射對西北羊茅氣體交換參數的影響
凈光合速率是單位時間內單位面積的干物質積累量,其值大小反映植物對光能的轉化能力[22]。植物光合生理特征(如:凈光合速率、氣孔導度、蒸騰速率、水分利用效率等)存在顯著的種間及環境差異[23]。本試驗發現在光合有效輻射800~1000 μmol·m-2·s-1時,西北羊茅通過較大的氣孔導度、蒸騰速率和較低胞間CO2濃度,從而保持較高的凈光合速率,與在羊草的研究類似[24],不同于玉米(Zeamays)的研究在1500 μmol·m-2·s-1時凈光合速率較高[20]。張歲岐等[25]認為氮、磷營養的合理補充能調節植物的滲透和氣孔大小,進而改善植物光合特性。本試驗,CK處理氣孔導度和蒸騰速率均低于施肥處理,與其較為一致。可能是CK處理土壤內可獲得N、P營養較低,使西北羊茅通過根系吸收的營養不能滿足細胞的N、P代謝循環[26]。已有研究表明,P影響卡爾文循環酶活性及RuBP羧化酶的再生[27],施磷增強小麥(Triticumaestivum)旗葉ATP酶和光合磷酸酶活性[28],N影響光合過程的C代謝相關酶活性[29]。本試驗在氮肥或磷肥添加后西北羊茅最適光合有效輻射值均增大,可見植物會改變自身光合特性以適應土壤養分的缺乏。
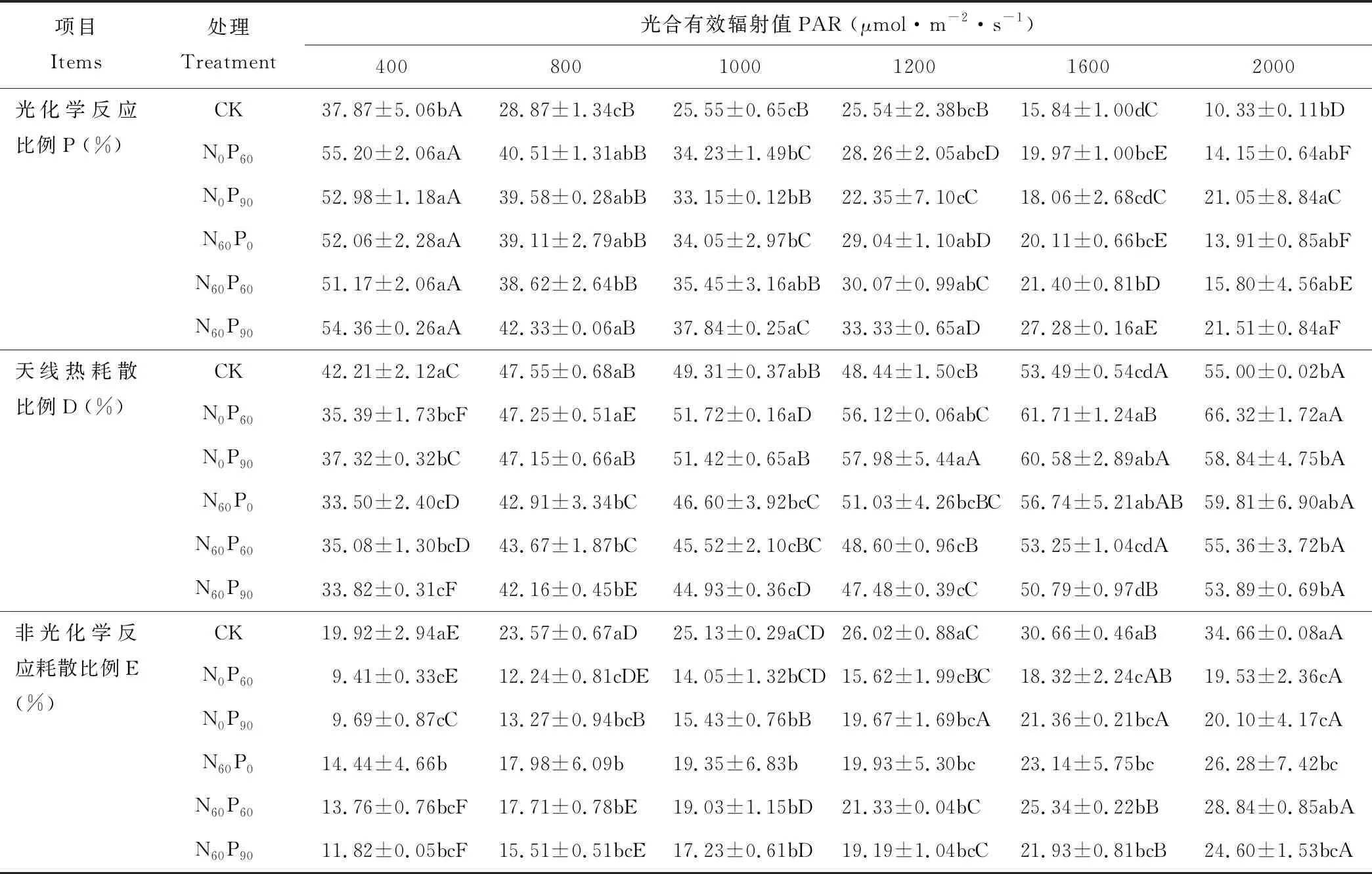
表3 施肥和光合有效輻射對西北羊茅葉片吸收光能分配的影響Table 3 Effect of fertilization and photosynthetically active radiation in absorbed light energy allocation in leaves of F. kryloviana
施肥是常見的農藝措施,施氮提高小麥和黑麥草(Loliumperenne)葉片光合速率[30-31],肖勝生等[11]發現施氮不僅提高羊草的凈光合速率,還延長其峰值持續時間。氮素添加提高滲透脅迫下的黑麥草葉片氣孔導度、蒸騰速率和氣孔限制值[31]。植物缺P影響木質部的發育進而降低植物水分吸收[32],從而減小氣孔導度和蒸騰速率[33],施磷增加苜蓿(Medicagosativa)葉片葉綠素含量和氣孔導度,提高光合特性[34]。與本試驗保持一致,氮肥和磷肥均提高了氣孔導度、氣孔限制值和蒸騰速率,降低了胞間CO2濃度,從而促進凈光合速率提高;而本試驗氣孔導度與胞間CO2濃度均變化相反,可見光合有效輻射和施肥主要通過調節氣孔影響西北羊茅光合能力[35]。水分利用效率常作為植物節水能力的評判標準,體現在3種尺度水平:葉片水平、植株水平和產量水平[36]。氮素促進羊草葉片增大氣孔導度,減小胞間CO2濃度和降低蒸騰速率,從而提高水分利用效率[11]。本試驗氮肥和磷肥添加均增大氣孔導度和減小胞間CO2濃度,而蒸騰速率的相對凈光合速率的慢速增加使水分利用效率提高,可能與種間的光合響應機制差異有關。施肥減弱植物細胞強光下的“午休”現象[37],N、P共同添加較單獨添加表現更優[38]。本試驗也有類似發現,氮肥和磷肥配施顯著提高光照強度超過1200 μmol·m-2·s-1后的凈光合速率,從而一定程度的減弱了強光造成的“午休”現象。另外,由于本試驗的最高氮肥施用量為60 kg·hm-2,據青藏高原地區的氮沉降速率[7],可見未來5~7年的氮沉降有利于西北羊茅生長。
3.2 施肥和光合輻射對西北羊茅葉綠素熒光參數的影響
葉綠素熒光參數反映植物光合生理狀況對環境因子的響應[31]。本試驗發現,隨光合有效輻射增強,PSⅡ有效光化學效率、PSⅡ實際光化學效率和光化學猝滅系數均降低,非光化學猝滅系數增加。可見西北羊茅通過關閉PSⅡ開放程度,降低對光能的捕獲和轉化,將過剩光量通過天線色素以熱能形式散失,減輕對光合器官的破壞,增強對強光的適應性[21]。而電子傳遞速率與凈光合速率保持一致,可證實通過較高的電子傳遞以維持PSⅡ活性。研究表明,受環境脅迫時植物葉片光合細胞的PSⅡ最大光化學效率值低于0.8[39],本試驗僅N60P90處理大于0.8(表2),且CK處理較施肥處理有更小的PSⅡ實際光化學效率和光化學猝滅系數,可確定為本試驗區域西北羊茅受到了N、P營養脅迫或N、P營養不平衡[33]。氮肥提高早熟禾和羊草PSⅡ實際光化學效率、電子傳遞效率和光化學猝滅系數[5,13]。外源NO添加提高黑麥草PSⅡ有效光化學效率[31]。施磷提高玉米PSⅡ實際光化學效率和光化學猝滅系數[14],本試驗施肥后PSⅡ有效光化學效率、PSⅡ實際光化學效率、光化學猝滅系數和電子傳遞速率的顯著增加與此一致,施肥增強對光能的捕獲和轉化能力。
3.3 施肥和光合輻射對西北羊茅葉片吸收光能分配的影響
植物葉片將吸收的光能用于競爭性的光化學反應、天線熱耗散和光反應中心非光化學反應耗散三部分[21]。本試驗,西北羊茅減少光化學反應比例,天線熱耗散和非光化學反應耗散部分增加,尤其在光合有效輻射超過1200 μmol·m-2·s-1后。可進一步證實植物為減小對光合機構的破壞,采取增加光能熱耗散的適應策略[5]。施肥主要通過減小非光化學反應比例進行補償用于光化學反應比例(表3),以促進光能的利用,與在黑麥草的研究保持一致[31]。
4 結論
1) 西北羊茅通過調節葉片氣孔導度以促進水分和CO2的交換速率從而提高凈光合速率,電子傳遞速率是凈光合速率增加的主要光合生理因子。
2) 未來幾年內西北羊茅生長受益于氮沉降,氮肥和磷肥配施提高西北羊茅葉片用于光化學反應比例,促進光能的傳遞和轉化,提高光能利用率。
3) 從葉片光合特性看,光合有效輻射值為800~1000 μmol·m-2·s-1時凈光合速率較高,即上午9:00-10:00和下午17:00-18:00是西北羊茅光合產物的快速積累時期。
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經濟(2017年6期)2017-07-12 09:27:16
中國衛生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛生(2014年11期)2014-11-12 13:11:32
