鳳丹牡丹PoFAD7基因的克隆及表達分析
2019-11-20 05:46:00李超瓊王雪芹劉紅占王會芳朱英男田輝輝
中國油脂 2019年9期
李超瓊,王雪芹,劉紅占,王會芳,朱英男,田輝輝
(1.周口師范學院 生命科學與農學學院,河南 周口 466001; 2.周口師范學院 新聞與傳媒學院,河南 周口 466001)
牡丹隸屬于芍藥科芍藥屬牡丹組,為多年生小灌木,具有很高的觀賞和藥用價值。自2011年牡丹籽油被批準為新資源食品,油用牡丹作為一種新型木本食用油料植物引起廣泛關注。目前,國家公布的油用牡丹品種為鳳丹牡丹和紫斑牡丹,其中鳳丹牡丹以生長適應性強、種子產量高等顯著特點在全國大面積種植[1-3]。牡丹籽油含有豐富的不飽和脂肪酸,主要為α-亞麻酸、油酸、亞油酸等,總不飽和脂肪酸含量可達93.23%[4-5]。α-亞麻酸屬于ω-3 系列不飽和脂肪酸,為人體不能自主合成的必需脂肪酸,是二十二碳六稀酸(DHA)和二十碳五稀酸(EPA)的前體,具有降血脂、降膽固醇、促進大腦發育和預防心腦血管疾病等作用[1, 6-8]。
ω-3脂肪酸脫氫酶是高等植物脂肪酸生物合成途徑中的關鍵酶,將亞油酸催化合成亞麻酸,是α-亞麻酸合成的關鍵酶[9]。在高等植物中,ω-3脂肪酸脫氫酶可分為定位于內質網中的FAD3和定位于質體的FAD7和FAD8[10-11]。目前,FAD7基因在擬南芥、大豆、油菜、煙草、麻瘋樹、光皮樹等多種植物中得到克隆,且發現FAD7基因能響應多種環境因素,如低溫、創傷等[9, 12-14]。研究發現,超量表達擬南芥FAD7基因的轉基因煙草α-亞麻酸含量增加,亞油酸含量減少,且轉基因植株抗寒能力也得到提高,在1℃低溫下仍能正常生長[15]。另外,超量表達FAD7基因的轉基因番茄抗寒性也有所增加[16]。由此可見,FAD7基因不僅影響植物的脂肪酸組成,還能改良植物對低溫的適應能力。
本研究從鳳丹牡丹中克隆得到α-亞麻酸生物合成的關鍵基因PoFAD7,并進行生物信息學分析及其在不同種子發育時期的表達模式研究,為進一步研究其在油用牡丹不飽和脂肪酸合成過程中的調控機理奠定基礎。
1 材料與方法
1.1 試驗材料
鳳丹牡丹種植于周口師范學院試驗田,2016年4—8月采集鳳丹牡丹不同發育時期的種子,樣本經液氮速凍后于-80℃冰箱保存待用。
多糖多酚總RNA提取試劑盒購自北京天根,Trans 2K plus II DNA Marker、TransStar taq酶和感受態細胞購自北京全式金生物技術有限公司,RACE試劑盒、T/A克隆試劑盒、反轉錄試劑盒(PrimeScript? RT Reagent Kit with gDNA Eraser)購自大連寶生物工程有限公司。
1.2 試驗方法
1.2.1PoFAD7基因的克隆
以鳳丹牡丹種子不同發育期轉錄組測序數據(SRA accession number: SRP051810)為基礎,找到一條含有部分PoFAD7基因的EST序列,根據該序列設計特異引物(見表1)進行3′-RACE。按照多糖多酚總RNA提取試劑盒操作步驟提取總RNA,檢測總RNA完整性和濃度,并按照RACE試劑盒操作步驟反轉錄合成cDNA。利用已設計好的引物進行RT-PCR擴增獲得PoFAD7基因3′端基因序列片段,與轉錄組數據中的EST序列進行拼接獲得PoFAD7基因完整的開放閱讀框。以PoFAD7基因序列為模板,設計含有起始密碼子和終止密碼子的特異引物,擴增獲得PoFAD7基因的開放閱讀框全長序列。

表1 引物序列
1.2.2PoFAD7基因cDNA序列生物信息學分析
使用在線工具ProtParam(http://web.expasy.org/protparam/)進行蛋白質親水性預測和理化性質分析。使用在線工具TMHMM (http://www.cbs.dtu.dk/services/TMHMM/)分析PoFAD7 蛋白質序列跨膜結構域。從GenBank下載其他物種FAD7蛋白氨基酸序列,使用DNAMAN軟件進行序列相似性比對。應用軟件MEGA5.0鄰位連接法構建PoFAD7 蛋白的系統進化樹。
1.2.3 鳳丹牡丹種子亞油酸和α-亞麻酸含量測定
自鳳丹牡丹授粉后每隔10 d采集不同發育時期的種子,在45℃烘箱中烘至恒重。采用索氏提取法提取鳳丹牡丹籽油,所得牡丹籽油經甲酯化處理,以氣相色譜-質譜聯用儀(Agilent 5977A Series GC/MSD System)分離和鑒定脂肪酸組分,測定干燥種子中亞油酸和α-亞麻酸的含量。具體步驟參照秦亞龍等[17]的方法。
1.2.4PoFAD7基因定量表達分析
提取鳳丹牡丹不同發育時期的種子總RNA并反轉錄為cDNA為模板,并以獲得的PoFAD7基因序列為模板設計特異性引物,以UBQ基因為內參基因進行熒光定量PCR試驗檢測PoFAD7基因的表達情況。熒光定量PCR按照SYBR Premix Ex TaqTMⅡ(TaKaRa)試劑盒說明在儀器 CFX96 Real-Time PCR Detection System (Bio-Rad 公司)上進行。擴增反應程序為:95℃ 3 min,95℃ 15 s,58℃ 15 s,72℃ 15 s,擴增40個循環。試驗所用引物見表1。
2 結果與討論
2.1 PoFAD7基因克隆
根據鳳丹牡丹種子轉錄組數據獲得951 bp的PoFAD7基因片段,依據所獲得的片段設計特異性引物,以鳳丹牡丹種子cDNA為模板進行3′-RACE,獲得3′末端序列長度約為800 bp(見圖1)。對所獲得的基因片段進行拼接,獲得全長cDNA序列為1 666 bp。根據所拼接的序列設計包含起始密碼子和終止密碼子的特異性引物進行PCR,獲得包含一個完整的開放閱讀框(1 353 bp)的基因片段(見圖1),編碼450個氨基酸(見圖2)。將該片段與拼接的PoFAD7基因序列進行比對分析,顯示兩序列一致,表明PoFAD7基因拼接正確。
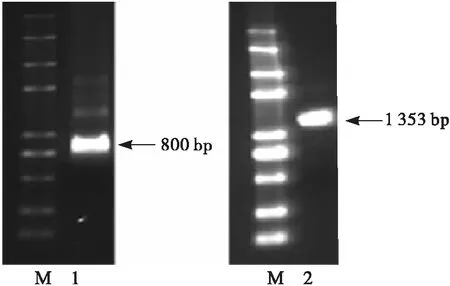
注:M. Trans 2K plus Ⅱ;1. 3′-RACE; 2.鳳丹牡丹PoFAD7基因開放閱讀框PCR產物。
圖1 鳳丹牡丹PoFAD7基因克隆

注:方框表示起始密碼子ATG;“*”表示終止密碼子。
2.2 PoFAD7蛋白理化性質及氨基酸序列比對
ProtParam分析結果顯示,PoFAD7蛋白分子式為C2 359H3 543N643O631S14,理論等電點為9.04,相對分子質量為51.4 kDa,理論半衰期為30 h,不穩定系數為31.75,屬于穩定蛋白,且其親水性平均系數為-0.285。應用TMHMM在線工具分析該蛋白具有2個明顯的跨膜結構域,分別位于131~153、285~307位氨基酸(見圖3),表明PoFAD7蛋白為跨膜蛋白。
將PoFAD7蛋白的氨基酸序列與其他物種中已克隆的FAD7基因的氨基酸序列進行比對分析,發現該蛋白與已報道的FAD7蛋白超家族的保守結構域具有高度一致性。氨基酸序列中有3 個保守組氨酸中心位點(HDCGH、HRTHH和HVIHH)(見圖4),這是維持膜結合類脂肪酸去飽和酶活性所必備的結構域[18]。另外,PoFAD7的氨基酸序列與芍藥FAD7氨基酸序列相似性最高,相似度達95%。

圖3 鳳丹牡丹PoFAD7蛋白氨基酸序列跨膜結構域預測

注:PoFAD7為鳳丹牡丹FAD7;NtFAD7為煙草FAD7;GmFAD7為大豆FAD7;AtFAD7為擬南芥FAD7;BnFAD7為油菜FAD7;RcFAD7為蓖麻FAD7;JcFAD7為麻瘋樹FAD7;PlFAD7為芍藥FAD7。組氨酸保守基序用方框標注。
圖4 鳳丹牡丹PoFAD7與其他物種FAD7蛋白氨基酸序列比對分析
2.3 PoFAD7蛋白的系統進化
利用MEGA5.0軟件,采用鄰位連接法構建PoFAD7 蛋白的系統進化樹,結果見圖5。
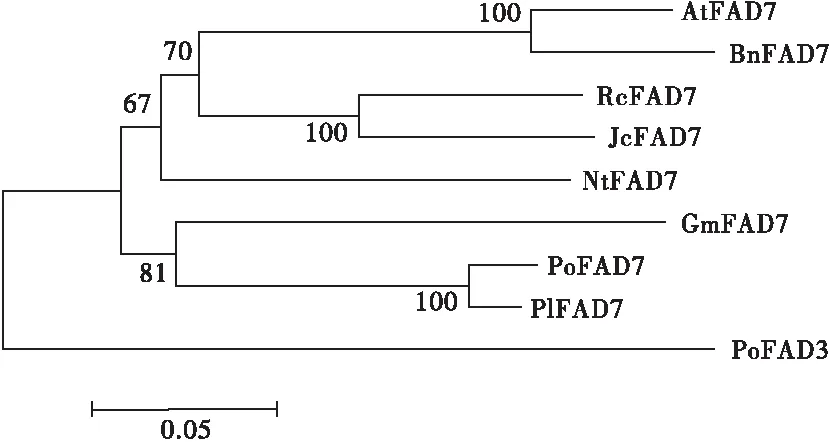
注:FAD7和FAD3蛋白來源為擬南芥AtFAD7(NP_187727);油菜BnFAD7(ACS16270);蓖麻RcFAD7(AAA73511);麻瘋樹JcFAD7(ABG49414);煙草NtFAD7(AIA22325);大豆GmFAD7(AEE25911);芍藥PlFAD7(AKE44631);鳳丹牡丹PoFAD3(ARJ54829)。
圖5 鳳丹牡丹PoFAD7與其他植物FAD7的系統進化樹
由圖5可知,PoFAD7與芍藥聚為一類,親緣關系最近,這也與鳳丹牡丹和芍藥同屬于芍藥屬植物相吻合。另外,在進化樹中發現PoFAD7與已克隆的鳳丹牡丹PoFAD3蛋白[19]聚為不同分支,表明兩者雖均屬于ω-3脂肪酸脫氫酶,但功能可能不盡相同。
2.4 不同發育時期鳳丹牡丹種子的亞油酸和α-亞麻酸含量
提取鳳丹牡丹授粉后10、20、30、40、50、60、70、80、90、100、110、120 d的不同發育時期鳳丹牡丹籽油并進行亞油酸和α-亞麻酸含量分析,結果見圖6。由圖6可知,從授粉30 d后鳳丹牡丹種子亞油酸和α-亞麻酸含量開始急劇增加直至授粉后80 d。在種子發育后期鳳丹牡丹種子α-亞麻酸含量在波動中平穩上升,而亞油酸含量大體平穩略有波動。
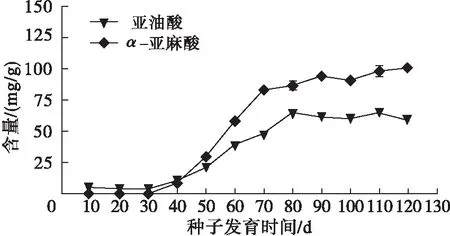
圖6 鳳丹牡丹不同發育階段種子亞油酸和α-亞麻酸的含量
2.5 熒光定量檢測PoFAD7基因表達量
利用qRT-PCR方法和PoFAD7基因的特異引物檢測其在鳳丹牡丹授粉后不同種子發育階段的表達情況,結果見圖7。
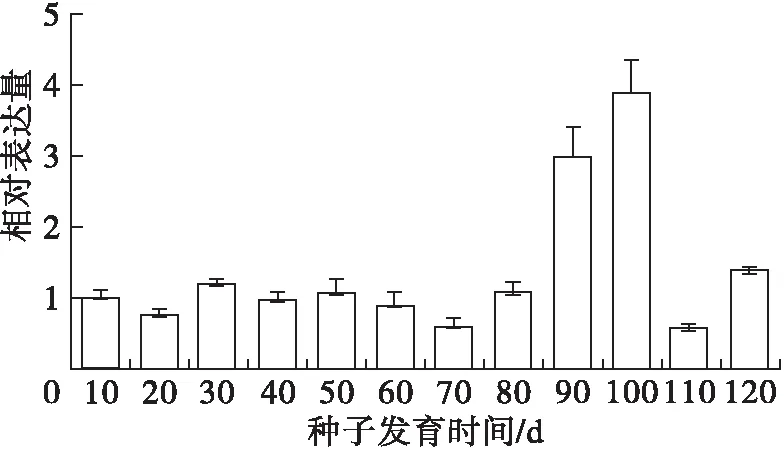
圖7 鳳丹牡丹PoFAD7在種子不同發育階段的相對表達量
由圖7可知,PoFAD7基因在種子發育的前80 d表達比較平穩,但在種子成熟后期的90 d急劇升高并在100 d達到最高后又下降。這與牡丹籽油中α-亞麻酸主要在授粉后30~80 d急劇累積的規律并不相符,說明PoFAD7可能并不是鳳丹牡丹籽油α-亞麻酸合成的主效基因。同時此種表達模式與已報道的光皮樹SwFAD7在果實中的雙峰表達模式也不一致[14]。但牡丹籽油中亞油酸含量在授粉90 d和100 d時含量略有下降,這表明PoFAD7在催化亞油酸合成α-亞麻酸的反應中可能也發揮了一定的作用。
3 結 論
ω-3 脂肪酸脫氫酶是多不飽和脂肪酸亞麻酸生物合成途徑的關鍵酶。本研究利用鳳丹牡丹種子轉錄組數據,采用RACE和RT-PCR方法克隆獲得了鳳丹牡丹PoFAD7基因序列(GenBank登錄號:MK205360),其包含完整的開放閱讀框,推測其編碼的蛋白質含有450個氨基酸。生物信息學分析表明PoFAD7蛋白具有2個跨膜結構域,含有3個保守的組氨酸基序,且系統進化樹分析顯示鳳丹牡丹與芍藥親緣關系較近。基因表達分析表明,PoFAD7在授粉后100 d的種子中具有較高的相對表達量。結合鳳丹牡丹種子中亞油酸和α-亞麻酸累積規律,推測PoFAD7在催化亞油酸合成α-亞麻酸的反應中發揮一定的功能,但可能并不是α-亞麻酸合成的主效基因。
