植被恢復對寒旱區典型草原群落枯落物分解的影響研究
2019-11-30 06:36:04王常順王慧清
生態環境學報 2019年10期
關鍵詞:影響
王常順 *,王慧清
1. 呼倫貝爾學院科學技術處,內蒙古 呼倫貝爾 021008;2. 呼倫貝爾市氣象局,內蒙古 呼倫貝爾 021008;3. 內蒙古自治區草甸草原生態系統與全球變化重點實驗室,內蒙古 呼倫貝爾 021008
枯落物的分解是陸地生態系統能量流動和物質循環的關鍵環節之一(郗敏等,2019)。草地地上初級生產力的大部分物質以枯落物的形式回歸到環境之中(Austin et al.,2006)。有關枯落物分解的研究多集中于森林生態系統和濕潤的草地生態系統,并對枯落物的分解過程和原理取得了較為統一認識(倪惠菁等,2019)。比如,枯落物分解過程的影響因素主要是溫度、降水、枯落物質量、土壤動物和微生物等(Lee et al.,2002);枯落物分解是以生物降解為主導的過程,環境因素直接或間接影響枯落物的分解(楊曾獎等,2007)。但在寒旱的生態系統,由于溫度和水分條件缺失,土壤生物尤其是微生物的活性受到極大的制約,枯落物分解的生物降解過程受限(霍利霞等,2019)。干旱半干旱區覆蓋了40%的陸地面積(Austin et al.,2006),枯落物分解過程與全球碳平衡和碳循環密切相關(呂憲國等,1995)。因此,研究寒旱區枯落物分解特征對全面理解草地碳氮循環變化趨勢具有重要意義(劉白貴,2008)。
在中國北方利用草地的主要方式是放牧(王德利等,2019)。過度放牧會改變群落的物種組成,甚至改變生態系統的物質循環,包括枯落物的降解過程(楊婷婷等,2019)。目前有關放牧對草地生態系統枯落物分解的影響研究工作在北美西部草原(Kemp et al.,2003;Angelina et al.,2007)、南美的潘帕斯草原(Gallo et al.,2006;Yahdjian et al.,2006;Austin et al.,2006)和澳洲的荒漠(Noble et al.,2009)、歐亞草原(Giese et al.,2009)均有開展。盡管這些關于放牧對枯落物分解影響的研究,一定程度上為寒旱區草地植被恢復過程中枯落物分解過程的理解提供了幫助,但植被的重度退化過程和植被的恢復過程顯然不能被簡單地對等(馬維偉等,2019)。目前對寒旱區植被恢復過程中枯落物分解特征尚不清楚,這限制了對寒旱區枯落物分解機制的深入認識。基于此,本文展開了對內蒙古典型草原不同恢復群落枯落物分解特征的研究,并探討環境因子對枯落物分解的影響,以期深入理解植被恢復條件下寒旱區草地枯落物分解過程的響應機制,進而估計和認識區域內的物質循環與能量流動。
1 試驗區概況
試驗樣地位于內蒙古自治區錫林郭勒盟錫林河中游,設于錫林河二級階地與丘陵坡麓之間,地理坐標為 116°05′—117°05′E,53°26′—55°08′N。氣候類型為大陸性季風氣候,年降水量約350 mm,年平均氣溫為-0.1 ℃。試驗樣地地帶性土壤為栗鈣土,pH為7.5—8.5,腐殖質含量1.5%—3%,土壤容重 1.21—1.43 g?cm-3(王海瑞,2011)。地勢微傾斜,地表較平整,具備顯域生境的基本條件。
試驗選取了3種不同植物群落,分別為:恢復25年群落,該樣地1983年圍封,圍封時為嚴重重度退化的冷篙(Artemisia frigida)+糙隱子草(Cleistogenes squarrosa)群落,現為羊草(Leymus chinensis)+大針茅(Stipa grandis)群落;恢復12年群落,該樣地 1996年圍封,圍封時為嚴重重度退化的冷篙+糙隱子草群落;重度退化群落,該樣地是仍處于放牧狀態的嚴重重度退化草原群落,與上兩個群落僅以圍欄相隔。
2 試驗方法
試驗采用分解袋法,分解袋孔徑分別為 0.23 mm(70目)和0.15 mm(100目),由規格為30×20 cm的尼龍網袋制成。試驗于2007年6月中下旬分別在3個群落內設置8個(共24個)1 m×1 m樣方。將原群落植物樣品在105 ℃下殺青15 min,之后在65 ℃下烘干至恒重。植物樣品剪成15 cm的小段,混合均勻分別裝入尼龍袋,每袋裝20 g。尼龍袋于2007年6月30日布設于采樣樣地,用筷子固定分解袋四角,平鋪于去掉地表枯落物的地表,模擬自然狀態分解。試驗設置3個重復,分8期回收,共計144個分解袋。分別于樣品投放后的第15、30、45、60、75、300、330和360天回收分解袋(2007年6月15日—2008年7月1日),同時測定土壤含水量;烘干后計算殘留率和分解速率。
使用球磨儀磨碎所有樣品,根據不同指標測定要求,取出適量樣品用于測定。總有機碳含量測定使用用德國元素Liqui TOC總有機碳測定儀;采用凱氏定氮法測定總氮(TN)含量;枯落物分解殘留物中總磷(TP)含量采用H2SO4-H2O2高溫消解,鉬銻鈧分光光度法測得;所用儀器普析通用 T6分光光度計。用萬分之一天平稱取過 0.25 mm篩的植物樣品1 g,之后在意大利VELPFIWE6纖維素測定儀上測定枯落物酸性纖維素和酸性木質素的含量,利用馬弗爐測定殘留物中不溶性灰分的含量。
枯落物干物質殘留率(RW)計算公式:

式中,Rw為枯落物干物質殘留率(%);mt為t時刻枯落物干質量(g);m0為枯落物的初始干質量(g)。
枯落物干物質殘留率變化運用 Olson單項指數模型進行計算:

式中,a為擬合參數;k為分解系數;t為時間。
采用雙因素方差分析(ANOVA)分析重度退化群落、恢復12年群落、恢復25年群落和不同時間的分解袋之間的殘留率的差異。采用單因素方差分析(ANOVA)分析同期3種群落枯落物中總氮(TN)、總磷(TP)、總有機碳(TC)、木質素、纖維素和灰分的差異。數據分析之前所有數據經過正態分布檢驗。所有數據均在 SPSS 17.0中處理。
3 結果與分析
3.1 植被恢復對枯落物物質殘留率和分解速率的影響
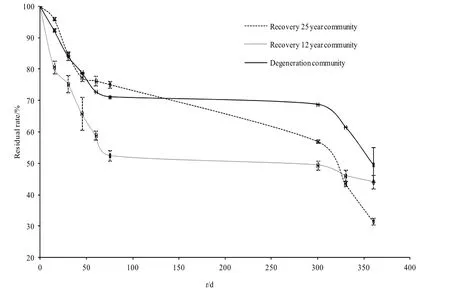
圖1 不同恢復梯度枯落物殘留率變化Fig. 1 Variation of residual rate of litter in different recovery gradients
植被恢復對內蒙古典型草原的枯落物分解過程具有顯著影響。隨植被恢復演替,各群落枯落物分解過程呈現出不同變化趨勢(圖1)。在分解初期,恢復12年群落枯落物的殘留率明顯低于恢復25年群落和重度退化群落。經過75 d的分解,恢復12年群落枯落物的物質殘留率已降低到52.41%,而恢復 25年群落和重度退化群落枯落物的物質殘留率分別為75.13%和71.18%。自當年生長季末開始,即枯落物分解75 d以后,枯落物的物質殘留率降低速率明顯減緩。在經歷一個非生長季,恢復 25年群落枯落物的殘留率降低了18.13%,而恢復12年群落和重度退化群落枯落物的物質殘留率分別降低2.99%和2.93%。自第2年生長季初,即分解試驗進行300 d后,枯落物的物質殘留率下降速率增大,截至試驗結束,恢復 25年群落枯落物的殘留率為31.57%,恢復12年群落和重度退化群落枯落物的物質殘留率分別為44.11%和 49.63%。在枯落物分解的不同階段,恢復 12年群落枯落物的物質殘留率均較重度退化群落低,說明植被恢復加速了典型草原枯落物分解過程。相較于與恢復 12年群落,恢復 25年群落枯落物失重率的變化表現得更為平緩,且最終失重率更低,說明植被恢復可以使得生物地球化學過程更均勻。
植被恢復和分解時間均對枯落物的物質殘留率具有顯著影響(P<0.001)(表1)。對3個群落枯落物物質殘留率進行兩兩比較后發現,恢復 25年群落與另外兩個群落的枯落物物質殘留率無顯著差異(P=0.411和P=0.132),恢復12年群落與重度退化群落枯落物物質殘留率具有顯著差異(P=0.021)(表 2)。說明適度恢復能夠最大程度地實現枯落物物質周轉的效率。

表1 恢復程度和時間對枯落物物質殘留率雙因素方差分析表Table 1 Two-way analysis of variance of recovery degree and time on litter material residual rate
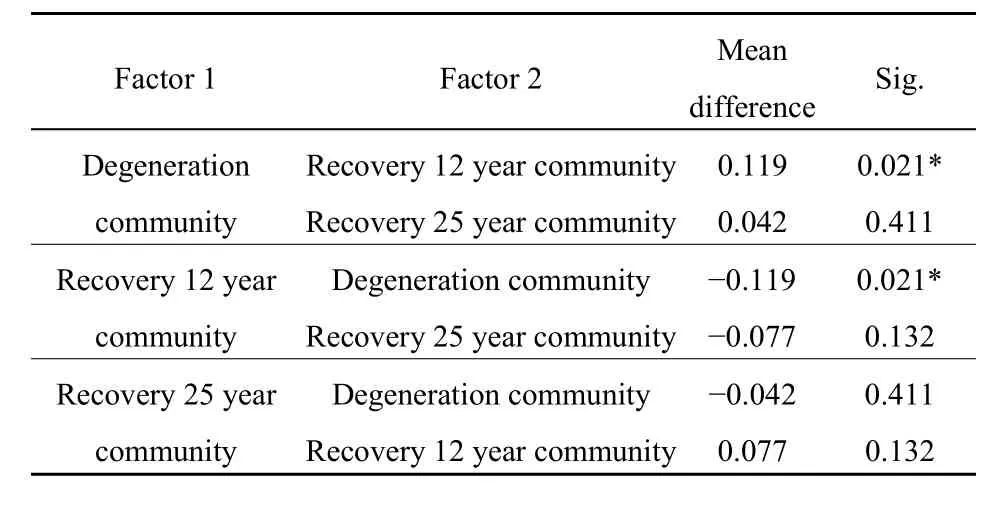
表2 不同恢復群落枯落物物質殘留率成對比較方差分析表Table 2 Paired comparison variance analysis table of litter material residual rate in different restoration communities
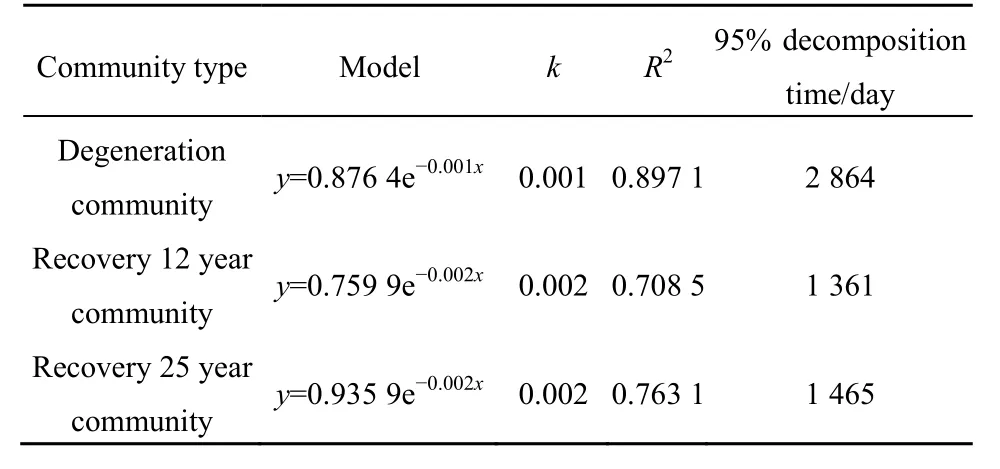
表3 枯落物物質殘留率與分解天數的指數方程及相應參數Table 3 Index equations and corresponding parameters of litter material residual rate and decomposition days
在360 d的分解期間,植被恢復對不同群落枯落物分解系數影響較小(表3),但減少了枯落物分解時間。經過25年和12年的恢復,枯落物分解95%時間分別為1465 d和1361 d,而重度退化群落為2864,植被恢復使得分解時間減少大約一半。
3.2 植被恢復對枯落物物質成分的影響
植被恢復對枯落物中 TOC含量存在顯著影響(P<0.05)(圖2a)。兩個恢復群落枯落物中的TOC含量顯著大于重度退化群落。兩個恢復群落枯落物中的TOC含量穩定;而重度退化群落的TOC含量在前30 d迅速下降,達30%左右,比其他兩個群落少20%,說明植被恢復降低了枯落物中有機碳的損失速率。恢復時間不同的3個群落間,枯落物分解過程中總氮和總磷含量無顯著差異(圖2b,2c)。
3個群落枯落物中的原始纖維素含量無明顯差異(0.1%、0.3%、0.4%)(圖2d)。但隨著分解時間的延長,重度退化群落枯落物中的纖維素含量顯著高于兩個恢復群落(P<0.05)。兩個恢復群落枯落物的木質素含量與重度退化群落在分解初期無顯著差異(P>0.05)(圖 2e),但是在分解后期,重度退化群落的木質素含量顯著高于恢復25年群落和恢復 12年群落(P<0.05)。無論是原始樣品還是在分解的過程中,重度退化群落的灰分含量均顯著高于恢復25年群落和恢復12年群落(圖 2f)。植被恢復降低了枯落物分解過程中的難分解物質含量。
4 討論
4.1 植被恢復對枯落物分解影響的機制
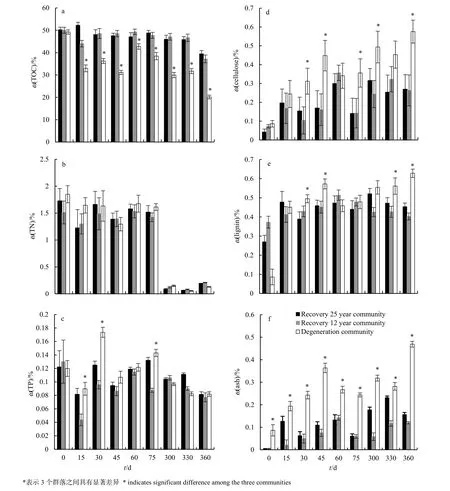
圖2 不同群落枯落物中TOC(a)、TN(b)、TP(c)、木質素(d)、纖維素(e)和灰分含量(f)Fig. 2 TOC (a), TN (b), TP (c), lignin (d), cellulose (e) and ash content (f) of litter in different community
普遍認為,枯落物質量是影響枯落物分解的重要因素(陳婷等,2016)。本研究結果表明,經過長時間的恢復,植物體中的物質成分發生了顯著的變化,其枯落物中的物質組成變化也區別于重度退化群落。恢復后枯落物中的 TOC含量顯著增加,同時又顯著降低了木質素、纖維素和灰分含量;這可能是恢復群落枯落物失重較多和分解周期較短的原因之一。本研究結果與以往部分研究具有一致性。如Bates et al.(2007)研究表明,過度放牧引起的枯落物質量的降低是分解速率下降的原因。但是與部分研究結果存在差異,如Giese et al.(2009)在內蒙古自治區典型草原群落的研究發現放牧對分解的作用不強。其研究認為干旱生態系統由于水分的缺失,引起微生物的分解作用下降。所以枯落物質量的差異對分解速率無明顯影響。植被恢復不僅改變枯落物質量,群落微環境也隨之變化。這也可能是枯落物分解速率變化的原因。Holt(1997)研究表明,過度放牧可能導致土壤酶活性的降低以及土壤微生物生物量下降,不利于枯落物的分解。另外,過度放牧降低了地表植被覆蓋,這使得地表溫度上升,并伴隨著水分散失加劇,從而阻礙凋落物的分解(姚國征,2017)。總之,植被恢復對枯落物分解過程的影響是多方面的,其機制還有待深入研究。
4.2 環境因素對枯落物分解的影響
在眾多氣候因素中,以氣溫和降水對分解過程的影響較為深刻(楊曾獎等,2007)。一般而言,溫度與分解速率呈正相關,主要是由于溫度能夠直接影響微生物活性和土壤動物的新陳代謝活動,致使分解速率隨氣溫的升高而增加(馬維偉等,2019)。試驗開始的6月多年月平均氣溫為18.8 ℃;試驗開展75 d后,10月的多年月平均氣溫為3.8 ℃,直至第2年3月,月均氣溫均低于0 ℃。試驗第2年第一次采樣的4月,多年月均氣溫為5.3 ℃。在非生長季,由于氣溫較低,微生物的活性可能受到極大的抑制。這可能是枯落物殘留率在試驗進行到非生長季時段變化較小的原因。這也進一步說明,氣溫的變化是控制寒旱區草地枯落物分解速率季節變化的重要環境因素。
在干旱生態系統中,微生物的活性可能因為水分條件較差而受到極大的限制,所以水分條件會影響枯落物的分解。在北美干草地研究中,Nakas et al.(1979)發現土壤含水量與土壤微生物活性呈正相關。同樣有研究表明,微生物活性在干燥和濕潤時具有差異顯著;濕潤時枯落物分解排放CO2的量是干燥的情況下的數倍(Kuehn et al.,2002)。Yahdjian et al.(2006)對南美干草地的研究證明分解速率與降水量存在正相關關系。本研究中,生長季階段恢復 25年群落前期枯落物分解失重率與本群落土壤表層0—10 cm含水量具有一定的協同變化關系(圖3)。因此,本研究結果進一步印證了在干旱生態系統中水分條件的變化影響著枯落物的分解。

圖3 典型草原恢復25年群落枯落物分解殘留率與土壤表層0—10 cm含水量Fig. 3 The weight loss rate of litter decomposition and the 0-10 cm water content of soil surface in the 25-year restoration of typical degradation grassland
5 結論
(1)長時間的植被恢復使得植物中難分解物質含量降低,同時也降低了枯落物分解過程中的難分解物質含量,這使得恢復群落枯落物的周轉速率加快。
(2)氣溫是控制寒旱區草地季節枯落物分解的重要環境因素,非生長季的低溫極大抑制了微生物的活性,進而使得枯落物的物質損失減緩;在溫度條件適宜時,水分條件的變化對枯落物分解速率有一定的影響。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
