多糖受體與巨噬細胞吞噬作用關(guān)系研究進展
2019-12-03 01:08:38金周雨
食品科學 2019年21期
趙 麗,金周雨,*,荊 波,宋 慧,2,*
(1.吉林農(nóng)業(yè)大學生命科學學院,吉林 長春 130118;2.吉林農(nóng)業(yè)大學 教育部食藥用菌工程研究中心,吉林 長春 130118)
多糖是由10 個以上單糖之間脫水形成的糖苷鍵連接而成的鏈狀聚合物,是生物體中重要的大分子物質(zhì)之一。多糖糖鏈結(jié)構(gòu)復雜多變,不同的糖苷鍵、單糖組成和分支化程度等,使其具有抗病毒、抗腫瘤、免疫應答、降血脂等多種生物活性[1-2],現(xiàn)已作為藥物、保健食品和食品添加劑得到開發(fā)利用,成為食品科學、生命科學、醫(yī)藥科學等領(lǐng)域研究的熱點。
巨噬細胞屬于免疫細胞,其吞噬與清除能力是啟動機體免疫的重要途徑之一,巨噬細胞表面具有多種受體,能參與介導細胞吞噬作用。一些研究發(fā)現(xiàn)多糖可通過巨噬細胞表面受體激活巨噬細胞釋放免疫因子,且多糖可促進細胞吞噬作用,但關(guān)于多糖增強巨噬細胞吞噬活性與巨噬細胞表面多糖受體之間的關(guān)系及其作用機制研究較少。
1 巨噬細胞吞噬作用
巨噬細胞吞噬作用起始于對目標顆粒的識別,并將目標顆粒攝入質(zhì)膜形成吞噬體。而后吞噬體與溶酶體融合形成吞噬溶酶體,目標顆粒被溶酶體內(nèi)的多種水解酶消化分解后被吸收在細胞質(zhì)內(nèi)[3]。巨噬細胞的吞噬作用是先天免疫中最基本的防御機制,其機制復雜獨特,與胞吞物質(zhì)特點密切相關(guān)。
巨噬細胞激活后,游走和吞噬是巨噬細胞運動的主要特征,其偽足的伸出與肌動蛋白的形成有關(guān)。研究表明,細胞在吞噬過程中其骨架結(jié)構(gòu)發(fā)生改變,作為細胞骨架蛋白的肌動蛋白發(fā)生重排,F(xiàn)-肌動蛋白在膜下聚集形成偽足;如果用抑制肌動蛋白聚合的藥物(如細胞松弛素)處理后,則不能形成偽足[4-5]。Orsi等[6]研究發(fā)現(xiàn),甲殼毒素可通過改變J774巨噬細胞中F-肌動蛋白排布來影響細胞骨架的完整性,從而抑制巨噬細胞的吞噬活性。
巨噬細胞的吞噬作用是識別、攝入與清除的復雜過程,由于胞吞物的專一性,巨噬細胞表面存在多種受體可介導其吞噬作用。例如,巨噬細胞對衰老紅細胞的識別引發(fā)的吞噬作用。衰老紅細胞表面因唾液酸的缺乏而暴露出半乳糖分子,而巨噬細胞表面有一類受體可對半乳糖分子進行識別從而促進其對衰老紅細胞的吞噬作用[7]。有研究表明,細胞表面有些受體與配體結(jié)合后會引起吞噬相關(guān)的信號傳導,但并不引起針對其配體的吞噬體的產(chǎn)生[8]。Bi Decheng等[9]發(fā)現(xiàn)海藻鹽多糖可增強巨噬細胞的吞噬活性,增加Toll樣受體4(Toll-like receptors 4,TLR4)的表達并激活蛋白激酶B(protein kinase B,Akt)/核因子-κB(nuclear factor-κB,NF-κB)和絲裂原活蛋白激酶(mitogen-activated protein kinase,MAPK)p38信號通路。加入TLR4、NF-κB和p38 MAPK抑制劑或敲低TLR4基因后,藻酸鹽促進的吞噬作用下降,其實驗結(jié)果與上述觀點一致。
2 多糖及其受體與巨噬細胞關(guān)系概述
2.1 多糖與巨噬細胞吞噬作用
有研究表明多糖免疫調(diào)節(jié)活性之一是促進巨噬細胞吞噬作用[10]。Yang Jiali等[11]從香蕉中提取的α-(1→6)-D-葡聚糖可通過提高T細胞增殖功能、巨噬細胞的吞噬功能、分化簇(cluster of differentiation,CD)3+T細胞水平等來改善免疫狀態(tài)。Tang Chao等[12]從紫甘薯中分離的水溶性多糖、稀堿溶性多糖和濃堿溶性多糖均可增強巨噬細胞吞噬活性。秋葵豆莢粗多糖可顯著增加腹膜內(nèi)巨噬細胞對金黃色葡萄球菌的吞噬活性[13]。山芝麻多糖可顯著促進巨噬細胞的增殖能力,刺激巨噬細胞的吞噬能力,以及誘導NO和免疫調(diào)節(jié)細胞因子的產(chǎn)生[14]。綜上所述,許多天然多糖均能促進巨噬細胞吞噬活性。這些研究多以巨噬細胞對中性紅的胞飲作用作為證明依據(jù),但在機體免疫中,細菌、病毒等引起的巨噬細胞吞噬作用與巨噬細胞表面多種受體有關(guān),具有特異性。因此,中性紅實驗并不能很好地證明吞噬作用的受體介導機制。
2.2 巨噬細胞表面多糖受體
巨噬細胞表面存在多種受體,其介導巨噬細胞發(fā)揮免疫功能。而在這些受體中有一部分可以識別多糖并與之結(jié)合,被稱為多糖受體。Sun Hongxiang等[15]研究發(fā)現(xiàn),獼猴桃根毛多糖可顯著增強巨噬細胞的胞飲和吞噬活性,誘導NO、腫瘤壞死因子(tumor necrosis factor,TNF-α)、白細胞介素-10(interleukin-10,IL-10)、IL-1β的產(chǎn)生,上調(diào)TLR2、TLR9、髓樣分化因子(myeloid differentiation factor88,MyD88)、β干擾素TIR結(jié)構(gòu)域銜接蛋白(TIR-domain-containing adaptor inducing interferon-β,TRIF)和NF-κB的mRNA表達水平,促進人NF-κB抑制蛋白α(IκB-α)在RA264.7細胞中降解和NF-κB活化。竹蓀多糖可結(jié)合巨噬細胞表面TLR4激活和NF-κB信號傳導途徑誘導巨噬細胞的活化,上調(diào)誘導型一氧化氮合酶(inducible nitric oxide synthase,iNOS)、IL-1β、IL-6和TNF-α的mRNA的表達,促進NO、L-1β、IL-6和TNF-α的產(chǎn)生[16]。多糖激活巨噬細胞首先與細胞表面多糖受體結(jié)合,激活胞內(nèi)信號轉(zhuǎn)導途徑,介導炎性因子如TNF-α、IL-1β、IL-6等釋放,進而增強免疫調(diào)節(jié)能力。現(xiàn)已發(fā)現(xiàn)的多糖受體主要包括TLRs、甘露糖受體(mannose receptor,MR)、Dectin-1、清道夫受體(scavenger receptor,SR)、補體受體3(complement receptor 3,CR3)等。
多糖、多糖受體及細胞吞噬作用三者間的關(guān)系歸納見表1。

表 1 巨噬細胞表面吞噬相關(guān)受體及其配體和相關(guān)機制Table 1 Macrophage surface phagocytosis-associated receptors and their ligands as well as corresponding signaling pathways
3 多糖受體與巨噬細胞吞噬的關(guān)系
巨噬細胞表面受體是激活巨噬細胞啟動免疫應答機制的關(guān)鍵之一。研究表明,多糖對巨噬細胞的活化是通過細胞表面的多糖受體實現(xiàn)的,多糖受體多為模式識別受體(pattern recognition receptors,PRRs)。PRRs可識別病原體表面的一些自身表達或與其產(chǎn)物所共有的、高度保守的病原相關(guān)分子模式(pathogen-associated molecular pattern,PAMP),如細菌表面的脂多糖、肽聚糖或病毒的核酸、蛋白質(zhì)等,介導巨噬細胞對病原微生物的吞噬清除;也可識別細胞或組織受損后釋放的損傷相關(guān)分子模式(damage-associated molecular patterns,DAMPs),進而介導巨噬細胞對凋亡細胞或受損組織碎片的吞噬清除[24]。目前對多糖受體與吞噬的關(guān)系的研究還較少。
3.1 TLRs與吞噬作用
TLRs是在果蠅中發(fā)現(xiàn)的第一個具有分子識別模式的I型跨膜蛋白,由可參與識別微生物病原相關(guān)分子模式的胞膜外區(qū)、能募集TIR信號分子的胞漿區(qū)和跨膜區(qū)3 個部分組成。由于TLRs胞漿區(qū)I與L-1R家族胞漿區(qū)高度同源,因此該區(qū)又被稱為Toll IL-1受體結(jié)構(gòu)域[25]。TLRs可監(jiān)控識別不同的病原分子,在機體抵抗病毒感染中起到重要作用,目前已經(jīng)發(fā)現(xiàn)的人類TLRs家族成員有11 個,其中TLR2和TLR4與巨噬細胞吞噬作用關(guān)系最為密切。
Lee等[26]研究表明蛹蟲草發(fā)酵液多糖可與巨噬細胞表面TLR4和TLR2結(jié)合,激活MAPK及NF-kB信號通路進而促進NO、ROS、TNF-α的產(chǎn)生,并增強巨噬細胞吞噬作用。王梁華等[27]研究表明LPS激活細胞后,產(chǎn)生的TNF相關(guān)的凋亡誘導配體(TNF related apoptosis inducing ligand,TRAIL)與TNF-α可以增強細胞本身的吞噬能力,而使用TLR2多抗則可抑制其吞噬作用。TLR2參與對金黃色葡萄球菌的識別,當通過siRNA敲低TLR2后可減弱RAW264.7中JNK磷酸化,并且降低金黃色葡萄球菌感染時的吞噬作用和細胞自噬[28]。Zhang Pei等[29]研究發(fā)現(xiàn)殼聚糖寡糖(chitosan oligosaccharide,COS)能顯著增強RAW264.7細胞對中性紅的吞噬作用,并且TLR4可識別COS增強巨噬細胞對FITC標記COS的胞飲作用。綜上所述,表明TLRs可直接識別胞吞物質(zhì)介導巨噬細胞吞噬,也可與其配體結(jié)合激活吞噬相關(guān)信號傳導途徑促進吞噬作用。
3.2 MR與吞噬作用
MR屬于C型凝集素超家族成員,是一種鈣依賴性I型跨膜糖蛋白,從N端到C端依次為富含半胱氨酸(cysteinrich domain,CR)結(jié)構(gòu)域,II型纖連蛋白(fibronectin,F(xiàn)NII)結(jié)構(gòu)域,8 個碳水化合物識別域(carbohydrate recognition domain,CRD),跨膜結(jié)構(gòu)域和細胞質(zhì)尾[19]。MR可識別含有甘露糖和巖藻糖殘基的多種糖分子,介導免疫細胞的內(nèi)吞作用來維持內(nèi)環(huán)境的穩(wěn)定,維系先天免疫與后天免疫,組成機體的一種免疫防御系統(tǒng)。有研究表明,黑靈芝多糖可作為巨噬細胞表面MR的配體增強MR的表達,抑制LPS誘導的炎癥反應,增強巨噬細胞吞噬活性,介導機體免疫反應[30-31]。Garcia-Aguilar等[32]通過實驗發(fā)現(xiàn),MR參與分枝桿菌誘導的凋亡細胞的吞噬作用,當?shù)蛲黾毎途奘杉毎佑|時,MR與暴露于凋亡細胞上的相應配體結(jié)合進而引起吞噬作用。Beharka等[33]研究表明,表面活性蛋白A(surfactant protein A,SP-A)可通過上調(diào)MR的表達增加肺泡巨噬細胞(alveolar macrophages,AMs)的吞噬作用,且SP-A可增強MR介導的對涂有莢膜脂多糖微球的吞噬作用,而該作用又可被MR另一配體甘露糖抑制。現(xiàn)階段,關(guān)于多糖、MR與巨噬細胞的研究多集中于多糖通過MR激活巨噬細胞免疫或由MR介導的巨噬細胞的吞噬活性等方面。而關(guān)于多糖在由MR介導的巨噬細胞吞噬活性方面影響的研究鮮見報道。但通過上述報道,可以大膽假設(shè)多糖能否通過上調(diào)MR的表達促進巨噬細胞對病原體的清除。
3.3 SR與吞噬作用
SR是另一類重要的模式識別受體,根據(jù)其結(jié)構(gòu)的多樣性被分為8 個不同的類別,而后又發(fā)現(xiàn)HVACR1、P2X嘌呤受體以及CD163 3 類不同的SR,尚未正式劃入SRs,但都與吞噬相關(guān),各類SR結(jié)構(gòu)特點如表2[34]所示。SR可識別LDL、LPS和LTA等,參與介導病原體、喪失唾液酸的衰老紅細胞和某些凋亡細胞的識別和清除[35]。在脂質(zhì)代謝、動脈粥樣化形成、先天免疫和宿主防御病原體等方面發(fā)揮重要作用[36]。
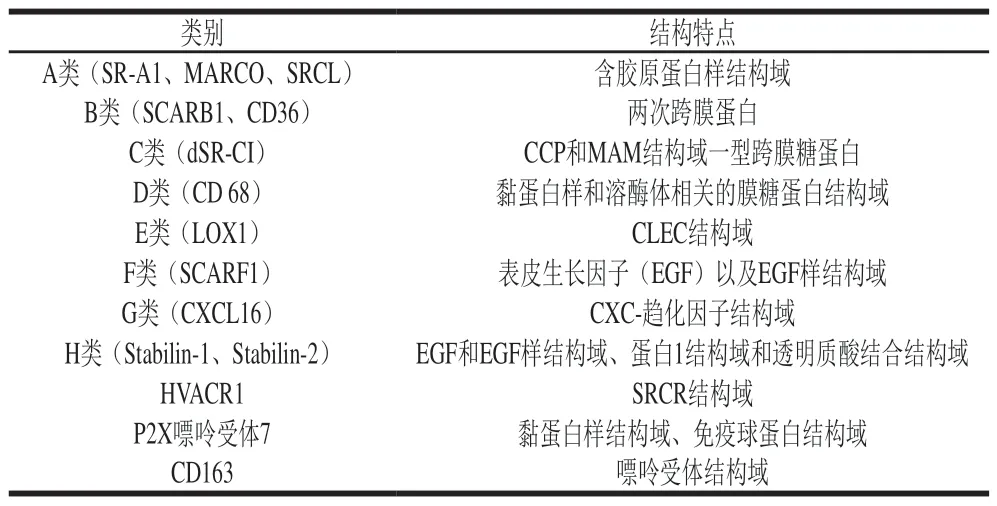
表 2 各類SR結(jié)構(gòu)特點[34]Table 2 Structural characteristics of various scavenger receptors[34]
在關(guān)于SR的研究中報道,如果由SR-A和CD36介導的氧化低密度脂蛋白(oxidize LDL,oxLDL)過度內(nèi)化則會導致泡沫細胞的形成。而β-葡聚糖和杏鮑菇多糖可降低巨噬細胞表面SR-A和CD36的表達,抑制巨噬細胞對oxLDL的攝取[37-38]。但在關(guān)于由SR介導巨噬細胞吞噬病原體的研究中,Seixas等[39]發(fā)現(xiàn),B型SR CD36是介導巨噬細胞吞噬伯氏瘧原蟲和大腸桿菌的受體,而細胞中Rab14的表達可以破壞其吞噬機制,Rab14沉默后可以降低CD36的內(nèi)化速率,增強巨噬細胞表面CD36的表達,促進其對伯氏瘧原蟲和大腸桿菌的吞噬作用。雖然有關(guān)多糖、SR和巨噬細胞吞噬作用三者之間的研究鮮見報道,但可以通過上述文獻推斷多糖對由SR介導的吞噬作用的影響有利有弊,其應用的利弊有待于進一步研究發(fā)現(xiàn)。SR雖然可直接識別配體并將其攝入細胞,但SR并不含細胞內(nèi)活化信號序列,其信號傳導機制可以與其他蛋白如TLRs相互作用進行[40]。高遷移率族蛋白1(high mobility group protein B1,HMGB1)是代表性的損傷相關(guān)分子模式(damage-associated molecular pattern,DAMP),在細胞損傷或壞死后釋放到細胞外空間。HMGB1可以通過TLRs介導胞內(nèi)信號傳導激活巨噬細胞,然而,HMGB1不與高親和力的TLRs結(jié)合,HMGB1的C末端酸性結(jié)構(gòu)域可被A類SR識別并結(jié)合介導巨噬細胞對其的內(nèi)化,HMGB1進入細胞內(nèi)可與由SR介導內(nèi)化的TLRs共定位,進而激活胞內(nèi)信號傳導途徑,釋放炎性因子調(diào)節(jié)免疫。且這種共定位也需要SR介導,其機制尚不明確[41]。由此可表明,SR雖不能直接參與細胞內(nèi)信號活化,但也是體內(nèi)宿主防御機制中介導吞噬與免疫應答的重要受體。
3.4 CR3與吞噬作用
CR3是白細胞整合素β2家族中的一員,主要表達于巨噬細胞、中性粒細胞和自然殺傷細胞中,在細胞免疫中介導細胞黏附和調(diào)理吞噬。CR3是由CD11b/CD18兩種跨膜蛋白組成的二聚體整合素,其CD11b亞基中有一個可以結(jié)合不同配體的共同結(jié)合位點,其配體為補體和β-葡聚糖[42]。由CR3介導的病原體清除作用,CR3與病原體的結(jié)合需要以補體的裂解產(chǎn)物C3b或C4b為中間橋梁。研究表明,CR3可以與經(jīng)iC3b調(diào)理的金黃色葡萄球菌結(jié)合,促進單核細胞衍生的巨噬細胞及樹突細胞對其進行吞噬[23]。CR3可結(jié)合并識別β-葡聚糖,激活FAK信號通路,促進ROS、TNF-α和IL-1β的產(chǎn)生介導豬中性粒細胞對β-葡聚糖的吞噬作用[43]。由此表明CR3也可與其配體結(jié)合,產(chǎn)生對應的信號級聯(lián)反應激活巨噬細胞,然后巨噬細胞通過吞噬作用直接殺死病原體,并呈遞抗原引發(fā)免疫反應,誘導細胞中的iNOS釋放NO,并且分泌炎性因子[44]。據(jù)報道,CR3的存在顯著降低了FcγR介導的吞噬作用中所需結(jié)合免疫球蛋白G(immunoglobulin G,igG)的量,且使用CR3抗體抑制后可顯著降低FcγR介導的吞噬作用,由此表明FcγR和CR3之間存在相互作用[45]。綜上所述,CR3可能與其他受體或蛋白協(xié)同作用,但其機制尚不明確,有待于進一步研究。
3.5 Dectin-1與吞噬作用
Dectin-1屬于膜相關(guān)C型凝集素家族中的一種II型跨膜結(jié)構(gòu)受體,主要表達于單核巨噬細胞、中性粒細胞、樹突狀細胞等細胞中。其結(jié)構(gòu)由C型凝集素樣受體碳水化合物識別域(C-type lectin domain,CTLD)、短桿連接的跨膜結(jié)構(gòu)域和具有免疫受體功能酪氨酸激酶活化基序(immunoreceptor tyrosine-based activation motif,ITAM)3 個部分構(gòu)成,可識別外源性病原菌上的LPS與T細胞表達的內(nèi)源性配體[46]。β-葡聚糖為其主要配體,據(jù)報道,真菌細胞壁β-葡聚糖與Dectin-1結(jié)合后可促進細胞骨架重排,形成吞噬小體,釋放大量的ROS、細胞因子和趨化因子[47],由此可推斷Dectin-1作為免疫相關(guān)的重要受體可參與介導巨噬細胞吞噬作用。
據(jù)報道,Dectin-1可與其他模式識別受體協(xié)同作用啟動免疫反應[48],β-葡聚糖與Dectin-1結(jié)合誘導病原體內(nèi)化,并與TLR-2一起誘導細胞因子的產(chǎn)生。Dectin-1還通過激活T細胞(nuclear factor of activated T cells,NFAT)通路的核因子增加促炎性細胞因子的產(chǎn)生,從而增強TLR2反應[49]。Deng Chao等[16]研究表明TLR4和Dectin-1均可作為竹蓀多糖受體,激活MAPK/NF-κB信號,促進IL-1β和TNF-α的分泌,促進細胞吞噬作用。有研究表明,Dectin-1和TLR9均參與介導巨噬細胞對煙曲霉和白色念珠菌的先天免疫應答反應,但二者作用不同且分工明確[50]。TLR9定位于細胞內(nèi),可識別非甲基化的胞嘧啶-磷酸-鳥嘌呤基序(CpG DNA),從而調(diào)節(jié)炎性細胞因子對真菌病原體的反應。Dectin-1與含有β-1,3葡聚糖的真菌病原體結(jié)合后,Dectin-1的細胞質(zhì)ITAM基序被Src家族激酶磷酸化,使脾酪氨酸激酶(spleentyrosine tyrosine kinase,Syk)活化。觸發(fā)吞噬體的酸化,并使TLR9向含有β-1,3葡聚糖的吞噬體中募集[50]。這種募集的最終目的尚不明確,但這可能為協(xié)調(diào)先天抗真菌防御提供新思路。且有研究表明,Dectin-1與β-葡聚糖結(jié)合后可促使肌動蛋白發(fā)生改變,巨噬細胞進行吞噬,激活胞內(nèi)信號傳導機制引起炎癥反應,但如果阻斷細胞吞噬作用,仍允許細胞表面持續(xù)的受體信號傳導[51]。由此可見,Dectin-1介導的抗菌免疫反應機制復雜,有待于進一步研究。
4 結(jié) 語
巨噬細胞作為一種免疫細胞,對病原微生物及受損組織的吞噬消除是通過細胞表面受體來實現(xiàn)。這些受體可直接識別病原分子或損傷相關(guān)分子,觸發(fā)誘導吞噬作用的信號級聯(lián)反應,也可與其配體結(jié)合,通過信號傳導途徑促進細胞吞噬作用或胞飲作用。多糖作為免疫調(diào)節(jié)劑,可激活巨噬細胞促進其吞噬作用,然而近年來對細胞表面的多糖受體報道多為激活相關(guān)信號通路、促進免疫因子的釋放,而其在細胞吞噬中扮演的角色以及其相關(guān)機制都有待深入研究。有關(guān)巨噬細胞表面多糖受體與多糖促進其吞噬活性之間相互作用的研究將是理解多糖調(diào)節(jié)免疫的關(guān)鍵性環(huán)節(jié),并可為臨床治療提供可靠的依據(jù)。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學報(2017年2期)2017-07-05 08:13:02
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
