高油酸花生遺傳選育的研究進展
2019-12-09 08:44:00呂建偉胡廷會成良強饒慶琳
貴州農業科學 2019年11期
關鍵詞:檢測
呂建偉, 胡廷會, 成良強, 饒慶琳, 王 軍
(貴州省農業科學院 油料研究所, 貴州 貴陽 550006)
花生(ArachishypogaeaL.)脂肪酸成分主要包括油酸、亞油酸、棕櫚酸、硬脂酸、山崳酸、花生酸、花生烯酸和二十四烷酸8種。其中,油酸(45%~48%)和亞油酸(32%~35%)的含量占脂肪酸組成的80%以上[1-3],二者都是有益脂肪酸。油酸能夠降低人體血液中的低密度脂蛋白(LDL),保持高密度脂蛋白(HDL),減少心血管疾病的發生,對人體脂質代謝具有重要作用。與亞油酸相比,油酸的抗氧化能力強,高油酸花生(油酸含量80%、亞油酸2%左右)制品的貨架保質期是普通花生(油酸45%、亞油酸35%)的3倍以上[4]。油酸和亞油酸含量的比值決定花生油的品質,高油酸花生有助于提升營養價值和保障其制品質量安全。鑒于高油酸花生的生產意義,近年來國內外育種工作者已將提高油酸含量列為花生遺傳育種的主要目標之一。
當前,高油酸花生品種已在澳大利亞、阿根廷、美國、巴西、南非以及以色列等國家得到大規模推廣種植[5]。近年來,國內育種單位在高油酸花生品種選育方面取得較大進展,相繼育成冀花16、花育961、桂花37、開農176等近30個高油酸品種,有力推動我國普通油酸品種向高油酸品種的更新,為實現我國高油酸花生產業化提供材料支撐。借助當前分子輔助育種技術,制定合理的高油酸花生育種策略,是進一步提高高油酸育種效率的重要措施。因此,圍繞高油酸花生不飽和脂肪酸的生物合成途徑、高油酸性狀的遺傳機制、分子標記輔助育種技術以及育種策略等幾個方面展開綜述,旨在進一步深化高油酸花生育種實踐,加快我國高油酸花生育種步伐。
1 花生油脂中不飽和脂肪酸的合成代謝途徑
1.1 合成不飽和脂肪酸的重要酶類
花生油脂中的不飽和脂肪酸主要為油酸和亞油酸。甘油三酯(Triacylglycerol,TAG)是由1分子甘油與3分子長鏈脂肪酸酯化形成的脂肪分子,在成熟的油料種子脂肪中的含量約占97%,其脂肪酸種類和比例決定了脂肪酸組成。在TAG合成代謝的通路中,卵磷脂(Phosphatidylcholine,PC)是多不飽和脂肪酸合成的重要載體,90%以上的脂肪酸在PC上合成代謝。PC上酯化連接的油酸在脂肪酸去飽和酶(Fatty acid desaturase,FAD)催化下產生亞油酸,是油料種子中亞油酸合成的最重要途徑。在植物不飽和脂肪酸合成代謝途徑所涉及的FAD酶類中,酰基-ACP去飽和酶發揮了重要作用,其包括催化硬脂酸合成油酸的Δ9脂肪酸去飽和酶,以及催化油酸合成亞油酸的Δ12脂肪酸去飽和酶,而Δ12脂肪酸去飽和酶是直接決定花生脂肪酸中油酸、亞油酸含量的關鍵酶。
1.2 油酸、亞油酸的合成代謝途徑
在多不飽和脂肪酸合成代謝途徑中,卵磷脂(PC)上的亞油酸等多不飽和脂肪酸主要經2種途徑被組裝在TAG分子上。第1種途徑是發生在酰基-CoA池和PC間的酰基編輯(Acyl editing)反應,經溶血卵磷脂酰基轉移酶(Llysophosphatidylcholine acyltransferase,LPCAT)的酯化催化,完成油酰-CoA與PC的酰基化組裝,再經FAD2和FAD3酶脫氫后形成不飽和脂肪酸卵磷脂(PUFA-PC),又在LPCAT酶的反向催化作用下PUFA-PC去酰基化,釋放出的亞油酰-CoA(Polyunsaturated fatty acid CoA)進入酰基CoA池[6-7],最終亞油酰-CoA在二酰甘油酰基轉移酶(Diacylglyceryl transferase,DGAT)的作用下組裝在TAG上(圖 1b、c);第2種途徑是發生在二酰甘油和PC間的酯酰交換反應(圖1c),主要在磷脂二酰甘油酰基轉移酶(Phosphatidylglyceryl transferase,PDAT)、磷脂酰膽堿二酰甘油膽堿磷酸酶(Phosphatidylcholine diacylglycerol cholinephosphotransferase,PDCT)和肉堿酰基轉移酶(Carnitine acyltransferase,CPT)等酶催化下的多不飽和脂肪酸摻入TAG的合成途徑[8-12]。

圖1油料作物種子甘油三酯的合成路徑
Fig.1 Triglyceride synthesisi pathway in oil crop seed
2 花生高油酸性狀的遺傳基礎
20世紀80年代,NORDEN等[13]檢測發現高油酸花生種質F435,其油酸含量為80%、亞油酸含量為2%,與橄欖油中的油酸含量相當。高油酸花生種質F435在世界范圍內都有育種利用,我國育成的高油酸花生品種也多數來源于該種質[14]。由于該自然突變種質在育種上的廣泛應用,明晰其高油酸性狀產生的遺傳規律及其分子機制成為推進高油酸育種工作的關鍵。研究者將該突變體及其衍生材料作為研究對象開展了大量的研究工作,為高油酸花生品種選育奠定了理論基礎[15-21]。
2.1 花生高油酸性狀的遺傳規律
在花生高油酸性狀遺傳研究中,研究者采用孟德爾經典分離分析方法和數量性狀遺傳模型分析方法,將普通油酸和高油酸品種進行雜交,統計分離世代的普通油酸、高油酸性狀單株或株系的數量和分離比例,分析高油酸性狀的遺傳規律。研究者將F435與普通油酸品種的雜交分離世代開展油酸含量分析,證實花生高油酸性狀受2對隱性基因控制[18,22-23],基因型命名為ol1ol1ol2ol2或aabb。丁錦平等[24]以親本汕油162和SunOleic95R雜交得到的F2代遺傳群體進行油亞比值(O/L值)分析發現,O/L值受2對加性-顯性-上位性主基因控制;韓柱強等[25]利用高油酸親本8-153和普通油酸親本粵油13進行雜交,對2個親本、F1、F2和F2:3的油酸、亞油酸進行多世代聯合分析認為,花生油酸性狀和亞油酸性狀同時由2對主基因控制,其對油酸、亞油酸的作用效應相反,此外油酸含量還受其他微效基因的影響。BARKLEY等[5]利用高油酸和普通油酸花生種質配制了6個雜交組合,并分析了539個F2的脂肪酸成分和ahFAD2基因型,其中5個雜交組合F2的高油酸性狀遺傳由2對隱性基因(ol1ol1ol2ol2)控制,另1個雜交組合高油酸性狀的遺傳表現單基因(ol2ol2)遺傳分離模式,在分子水平上分析解釋了該組合普通油酸親本含有與高油酸親本相同的ol1ol1隱性基因位點,即由于ol1/FAD2A基因序列突變所致。
2.2 花生高油酸性狀的分子機制
花生油酸到亞油酸的轉化主要由ahFAD2A和ahFAD2B2對分別定位于A09和B09染色體上的同源非等位基因控制[26]。花生高油酸突變體F435的ahFAD2A和ahFAD2B基因序列在ORF區域具有99%的同源性,ahFAD2A基因的ORF-448 bp處發生G/A替換,編碼的天冬氨酸轉變為天冬酰胺,產生的FAD2A酶活性下降;ahFad2B基因的ORF-441_442 bp處發生A堿基插入導致移碼突變,產生的FAD2B酶完全失活。F435的高油酸性狀是由ahFAD2A和ahFAD2B兩對同源非等位基因的共同突變產生[16-17],與F435類型的高油酸突變體不同,化學突變創制的M2-225和C458高油酸突變體的ahFAD2B基因功能喪失是由反向重復轉座元件插入到ORF編碼區導致[27]。總之,花生高油酸性狀產生的分子機制與FAD2A和FAD2B基因的突變導致對應編碼的酶失活或活性下降有關。
2.3 花生 ahFAD2基因的遺傳效應
ahFAD2A和ahFAD2B2個非等位基因共包含9種基因型,其中aabb基因型花生的油酸含量最高[17],這些基因型按照突變位點數量分為0、1、2、3、4個5種類型。研究表明,隨著個體的ahFAD2基因突變位點數量增多,油酸含量升高,亞油酸含量降低,突變位點數量相同(如AaBb、aaBB和AAbb)的則沒有差異[5]。可見,ahFAD2A和ahFAD2B基因表現出一定的劑量效應。有研究發現,ahFAD2A對油酸和亞油酸的表型變異解釋率分別為17.4%和19.5%,ahFAD2B的表型變異解釋率分別為33.8%和41.0%,ahFAD2B的基因效應大于ahFAD2A的基因效應[19]。PANDEY等[26]在研究中同樣發現FAD2B的表型變異解釋分別為26.54%、25.59%和41.02%,FAD2A的表型變異解釋分別為8.08%、6.86%和3.78%。BARKLEY等[5]還認為,ahFAD2A和ahFAD2B基因存在加性效應,協同調控花生種子油酸含量,2對基因具有不完全顯性,子代油酸含量受親本表型影響,其他微效修飾基因也參與了油酸的合成代謝。
花生ahFAD2基因還與其他脂肪酸具有一定相關性。研究表明,油酸與亞油酸、硬脂酸、花生酸的含量呈顯著負相關,其中,與亞油酸相關性最高(相關性系數R=-0.701,P<0.0001)[21]。ahFAD2A和ahFAD2B基因型與油酸、亞油酸、棕櫚酸和油亞比(O/L)含量顯著相關,具有遺傳多效性[5,22,28]。
3 常用花生高油酸分子輔助選擇技術
在作物育種過程中,常規育種方法存在育種周期長、育種群體龐大且選擇低效等問題,為選擇出含有目標性狀的優異品系,需投入大量人力、物力。隨著生命科學技術的進步,分子輔助選擇技術(Marker assisted selection,MAS)已在作物育種上得到應用,其可快速、準確地將含有目標性狀和目的基因的個體從雜交子代中選擇出來。在花生高油酸育種中,根據ahFAD2A、ahFAD2B編碼區及突變位點(ahFAD2A448G>A、ahFAD2B441_442insA)信息開發出了一系列MAS檢測技術,包括酶切擴增多態性(Cleaved amplified polymorphisms,CAPS)[29-30]、qRT-PCR分型檢測[31-32]、PCR產物直接測序及位點特異性PCR(AS-PCR)[33-35]等技術。MAS技術能夠精準地將ahFAD2(a_b_)基因型的單株從F2或F3早期分離世代群體中篩選出來,去除未攜帶目標基因位點的單株,可縮小群體數量,提高育種效率4~16倍[36],育種周期可縮短3年左右。
3.1 CAPS分型檢測技術
酶切擴增多態性(Cleaved amplified polymorphisms,CAPS)技術利用限制性內切酶(如Hpy99I、DdeⅠ等)識別、酶切ahFAD2A/ahFAD2B基因448 bp處的 G>A和442 insA SNP位點,使酶切產物片段呈現大小不同的多態性,以此為判斷依據對FAD2A/FAD2B基因分型[23,29]。根據ahFAD2A基因的上游5′UTR區和編碼區設計特異引物,上游引物序列為5′- GATTACTGATTATTGACTT-3′,含有19 bp特異序列,下游引物5′-CCAACCCAAACCTTTCAGAG-3′。經PCR擴增,目的產物條帶大小826 bp,純化后利用能夠識別CGACG序列的限制性內切酶Hpy99I酶切,識別位點序列的堿基G是ahFAD2A基因序列448 bp處的G堿基,因而只有野生型的ahFAD2A基因擴增片段才能夠被酶切為598 bp和228 bp的產物,而純合隱性突變型的擴增片段則不能發生酶切(圖2)。
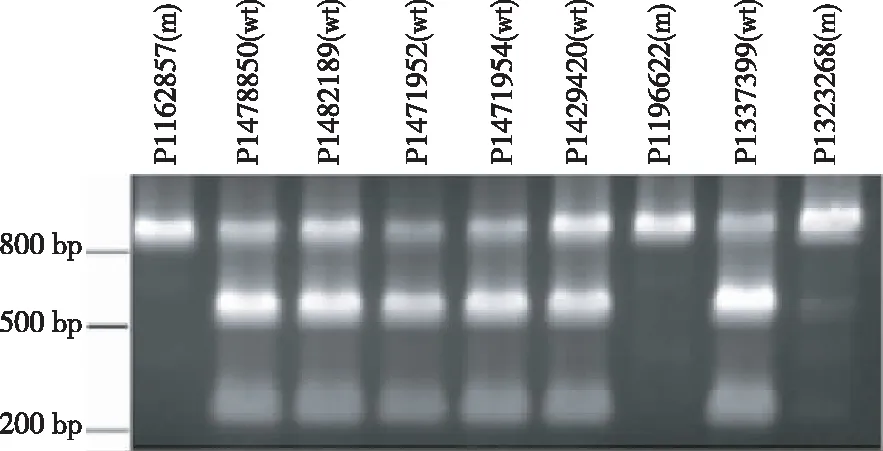
圖2 利用 CAPS鑒定花生核心種質 ahFAD2A的基因型
Fig.2 GenotypingahFAD2Aof peanut core collection employing CAPS technology
3.2 AS-PCR分型檢測技術
等位基因特異性PCR(allele-specific PCR)分型檢測技術基于SNP位點設計特異引物,通過PCR擴增、電泳,根據電泳結果的有無判斷目的基因的基因型。CHEN等[33]最先設計出花生FAD2基因的AS-PCR分型檢測方法,但其方法不能準確分辨出AaBB和aaBB或AABb和AAbb的基因類型。YU等[34]根據ahFAD2A(448G>A)/ahFAD2B(441_442insA)突變堿基的差異共設計7條AS-PCR引物(表1),通過將1個錯配堿基置于其中4條引物的3’端的第二或第三位以確保檢測的特異性。經4個PCR反應來檢測ahFAD2A/ahFAD2B的突變型或野生型以及通過瓊脂糖凝膠電泳條帶大小來判斷各位點的雜合或純合性,按照圖22〗3所示的4個PCR反應進行。其中,反應Ⅰ、Ⅱ、Ⅲ、Ⅳ分別鑒定ahFAD2A/ahFAD2B的野生型(A)、突變型(a)、野生型(B)、突變型(b)位點,反應Ⅰ、Ⅱ和反應Ⅲ、Ⅳ分別能夠雙向驗證ahFAD2A/ahFAD2B的純合性。在PCR反應中,每個反應體系需要加入3條引物,其他反應條件與常規PCR相同。
根據電泳結果,當反應Ⅰ中出現2條帶,則ahFAD2A含有野生型A,出現1條帶,則ahFAD2A不含有野生型A;當反應Ⅱ中出現2條帶時,則ahFAD2A含有突變型a,出現1條帶,則ahFAD2A不含有突變型a;同樣,反應Ⅲ中出現2條帶,則ahFAD2B含有野生型位點B,出現1條帶,則不含有野生型位點B;反應Ⅳ出現2條帶,則ahFAD2B含有突變型位點b,出現1條帶,則ahFAD2B不含有突變型位點b。如圖4所示,泳道1~9代表9種基因型。其中,泳道1表示野生純合基因型(AABB),泳道2表示AABb,泳道3表示AAbb,泳道4表示AaBB,泳道5表示AaBb,泳道6表示Aabb,泳道7表示aaBB,泳道8表示aaBb,泳道9表示aabb。該技術通過4次PCR反應,可快速、有效地將9種基因型分辨出來。
表1ahFAD2A/ahFAD2BAS-PCR引物序列信息
Table 1 AS-PCR primer information ofahFAD2A/ahFAD2B

引物名稱Primer引物序列Sequence (5' to 3') FAD2A-F GATTACTGATTATTGACTTGCTTTG FAD2A-G GTTTTGGGACAAACACTTCTTC FAD2A-A GTTTTGGGACAAACACTTCTTT FAD2B-F CAGAACCATTAGCTTTGTAGTAGTG FAD2B-C AACACTTCGTCGCGGTTG FAD2B-A AACACTTCGTCGCGGTTT FAD2-R CTCTGACTATGCATCAGAACTTGT
注:引物序列中下劃線標注指錯配堿基。
Note: Mismatched bases were underlined in some primer sequences.
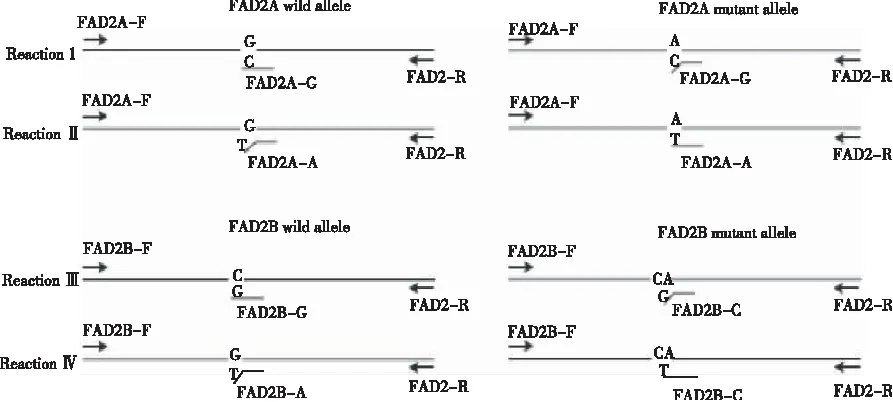
圖3 FAD2 基因分型的4個 AS-PCR反應
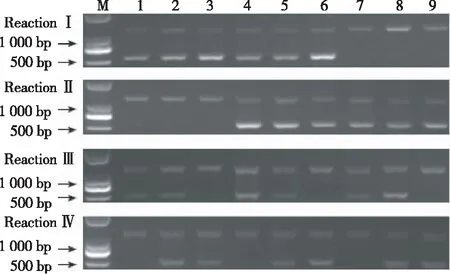
圖4 AS-PCR產物電泳檢測 ahFAD2A/ahFAD2B的基因型
Fig.4 Electrophoresis detection of AS-PCR products for genotypingahFAD2/ahFAD2B
3.3 Real-time PCR分型檢測技術
通過在ahFAD2A和ahFAD2B突變位點上各設計突變型探針和野生型探針,在兩端保守區域設計引物進行qRT-PCR反應,然后根據熒光信號分辨其基因型[31-32,37]。根據ahFAD2A和ahFAD2B的基因序列及其突變位點設計了引物,以及與各個基因對應的突變型6-FAM和野生型VIC熒光基團探針(表2),通過實時熒光定量PCR技術,分別用于檢測ahFAD2A和ahFAD2B的基因型,利用該方法可檢測所有的9種基因型。

表2 熒光定量 PCR技術的引物和探針信息
3.4 KASP分型檢測技術
KASP(Kompetitive Allele Specific PCR,KASP)是一種獨特的競爭性位點特異PCR,基于雙熒光共振能量轉移技術原理對SNP位點檢測分型[38-40]。ZHAO等[41]根據ahFAD2A和ahFAD2B的SNP位點信息,利用KASP技術原理將突變型等位基因引物序列ahFAD2A-Allele-l或ahFAD2B-Allele-l的5’端連接帶有FAM熒光染料的Allele-l Tail引物,同樣在野生型等位基因引物序列ahFAD2A-Allele-2或ahFAD2B-Allele-2的5’端連接帶有HEX熒光染料的Allele-l Tail引物(表3)。當引物與ahFAD2A或ahFAD2B序列結合,熒光染料與淬滅基團分離,產生熒光信號。不同的基因型產生不同的熒光信號,從而完成基因分型。兩條common引物存在堿基序列差異,可將ahFAD2A和ahFAD2B區分開,因此一個反應能夠同時完成ahFAD2A和ahFAD2B的基因分型。
表3ahFAD2A和ahFAD2B基因分型的 KASP引物信息
Table 3 Information of KASP primer sequences for genotypingahFAD2AandahFAD2B

引物Primer序列Sequence(5′ to 3′)AhFAD2A-Allele-lGTTTTGGGACAAACACTTCGTTAhFAD2A-Allele-2GTTTTGGGACAAACACTTCGTCAhFAD2A-commonCGCCACCACTCCAACACCAhFAD2B-Allele-lCAAACACTTCGTCGCGGTCTAhFAD2B-Allele-2CAAACACTTCGTCGCGGTCGAhFAD2B-commonCCGCCACCACTCCAACACAAllele-l Tail (FAM tail)GAAGGTGACCAAGTTCATGCTAllele-2 Tail (HEX tail)GAAGGTCGGAGTCAACGGATT
注:下劃線標識堿基為ahFAD2A和ahFAD2B野生型和突變型的SNP位點。
Note:SNP loci was marked with bold type and underlines both in wild and mutant types.
KASP技術方法中的熒光染料較qRT-PCR的Taqman探針的成本低,該技術還具有高通量、高度特異性和高度靈敏性等優點,因此被廣泛應用于水稻[42]、小麥葉片銹病抗性[43]、柑橘[44-45]、大豆線蟲抗性[46-47]、蘋果抗病性[48]等作物的SNP檢測分型。
3.5 PCR產物測序法
該方法是最直接、最準確檢測親本、雜交F1、BC1F1~BC4F1基因型的方法。親本基因型的鑒定需要2次PCR反應,利用引物對FAD2A-F/FAD2A-R進行單個PCR擴增親本的ahFAD2A基因片段,引物對FAD2B-F/FAD2B-R擴增親本的ahFAD2B基因片段(表4)。利用Internal1和Internal2測序引物對基因擴增片段進行雙向測序,根據突變型ahFAD2A在448 bp處G>A堿基和突變型ahFAD2B在442 bp處insA的SNP信息,讀取該序列位置的測序峰圖,判斷親本基因型[49]。

表4 花生 ahFAD2A和 ahFAD2B的PCR 產物測序的引物信息
3.6 技術評價
由于ahFAD2A/ahFAD2a、ahFAD2B/ahFAD2b均只存在1個堿基的差別,應用AS-PCR、熒光定量PCR和KASP技術進行PCR擴增時,無法避免假陽性結果[33],且利用AS-PCR、CAPS以及熒光定量PCR等技術對每份花生樣品的檢測需要至少2次PCR反應,檢測程序繁瑣。此外,CAPS檢測技術涉及酶切,對DNA質量要求高[29],且不能將純合野生型的AA和雜合基因型的Aa區分,只能用于檢測親本或種質資源等純合基因型材料。熒光定量PCR檢測技術應用的特異性熒光探針成本高[31-32],具有一定假陽性率。與以上技術方法相比,KASP檢測方法具有高通量檢測特點,適用于檢測分離世代中的成百上千個單株,同時結合其他檢測方法剔除假陽性單株。此外,利用F0.7/R3引物對進行一次PCR擴增和測序就能夠準確檢測ahFAD2A和ahFAD2B基因型,較以往報道的測序方法更加準確、高效。
3.7 分子輔助選擇技術應用
隨著花生高油酸性狀分子遺傳機制的解析,基因分型技術日趨成熟,在花生高油酸育種上得到了廣泛應用[50-51]。NAWADE等[28]利用AS-PCR技術和CAPS技術檢測印度174份花生種質的油酸基因型,80份花生種質在ahFAD2A位點的448bp處發生G>A堿基替換。李麗等[52]利用單粒近紅外光譜技術(Near infrared reflectance spectroscopy,NIRS)和氣相色譜(Gas chromatographic,GC)對F2:3家系和F4種子進行初步篩選,將篩選到的高油酸材料進行AS-PCR鑒定,得到純合高油酸材料4個,其油酸含量介于81.1%~82.7%。JANILA等[53]利用標記輔助回交和標記輔助選擇方法將SunOleic 95R高油酸材料的ahFAD2突變位點滲入到ICGV06110、ICGV06142和ICGV06420中,利用AS-PCR技術篩選出469份油酸含量在62%~83%的后代材料,較輪回親本提高50%~110%,亞油酸含量降低40%~100%,棕櫚酸含量降低10%~60%。WANG等[54]以美國花生微核心種質為研究材料,利用SNP功能標記進行熒光定量PCR,檢測ahFAD2A/ahFAD2B基因分型及分布情況,發現ahFAD2B基因未發生突變,所有的102份種質均為野生型;ahFAD2A存在3種基因型,其中58份種質為野生型,2份種質為雜合突變型,42份種質為純隱性突變型。可見,在花生高油酸育種工作中綜合利用分子標記輔助選擇技術和高油酸鑒定方法,可準確、高效地從育種后代群體中選擇出帶有高油酸性狀的目標材料。
4 高油酸花生育種策略
培育高油酸花生新品種的常用技術方法是回交育種,一般以輪回親本作為母本,高油酸供體親本和BCnF1作為父本。該方法具有以下幾方面的優點:一是采用回交法,2年可實現4~5次回交,育種后代的絕大部分細胞核遺傳物質來自輪回親本,可在短時間內穩定;二是在回交過程中,以輪回親本(AABB)做母本,如去雄不徹底則會產生單一基因型的假雜種(AABB),易于與真雜種(AaBb)區分;三是輪回親本作母本有利于將受細胞質遺傳控制的優異性狀傳遞給育種后代,如花生的青枯病抗性;四是以BCnF1作父本提供花粉,同時對其自交產生的F2、BCnF2種子進行系譜育種,從分離世代篩選高油酸單株。
通過連續雜交和回交,在此過程中利用分子標記輔助選擇技術對F1和BCnF1進行雜交種基因型鑒定,獲得基因型為AaBb的種子。在自交分離世代,利用近紅外線初步篩選出油酸含量達60%以上的F2和BCnF2的種子,縮小群體選擇的范圍后再利用KASP技術進行基因型檢測,篩選出基因型為aabb的高油酸單株。下一季將收獲的高油酸單株種子種植成株行,此時aabb基因型的高油酸性狀能夠穩定遺傳,只需從株行中篩選兼具株高、分枝數、種皮顏色、抗病性、產量以及含油量等其他育種目標的單株,在下一季將每株行性狀相近的中選單株混收種成株系,并繼續選擇形成品系參加區域聯合鑒定試驗。利用以上育種策略,每年開展2~3次雜交、回交,結合分子標記輔助選擇技術和近紅外線分析技術進行中間環節控制,可大大縮短育種進程[14,55-56]。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
