鉛脅迫對馬鈴薯幼苗生長形態(tài)及內源激素的影響
2019-12-10 02:34:08郭華春張光海思玲超
西北農業(yè)學報 2019年11期
宋 潔,郭華春,李 俊,張光海,思玲超
(云南農業(yè)大學 農學與生物技術學院, 昆明 650201)
重金屬對生態(tài)系統有潛在的不利影響[1]。土壤重金屬污染造成的農作物減產的問題一直受到世界許多國家的關注[2]。在中國,由于采礦、冶煉、三廢排放、過量施用化肥農藥等人為因素影響,導致污染土地生產出來的糧食重金屬含量超過國家食用標準[3]。鉛(Pb)作為土壤中富含的非必需元素,是關注度較高的重金屬之一,對環(huán)境和人類健康的影響也受到廣泛關注[4-6]。由于Pb毒性較大,在土壤中存在的普遍性和長期性,是中國重點監(jiān)測和控制排放的元素[7]。Pb對植物生命進程的不同影響是多因素共同作用的結果,有研究表明,植物形態(tài)特征與重金屬含量間存在正比例關系[8]。大劑量Pb可能會導致多數植物的代謝紊亂,生長抑制,甚至死亡[9]。已有的研究表明,高水平的重金屬可導致植物早期死亡,并可加速葉片的衰老速度[10-11]。也有研究表明,重金屬可延遲或減少開花而使葉片更新速度減慢,進而影響植物生命進程[12]。植物內源激素在調節(jié)植物生長發(fā)育方面起重要作用,生長素、赤霉素、細胞分裂素、脫落酸、乙烯、水楊酸等,參與植物的重金屬脅迫信號傳導[13-16]。研究顯示,在Pb脅迫下,除了形態(tài)特征、生理指標發(fā)生變化外,其激素水平也會產生明顯響應[17-18]。
馬鈴薯是中國的重要作物之一,隨著中國馬鈴薯主糧化戰(zhàn)略的推進,將利用南方的冬閑田擴大馬鈴薯的種植面積。但中國南方地區(qū)礦藏資源集中,重金屬污染嚴重[19-20]。目前,對馬鈴薯受Pb脅迫的形態(tài)學響應機制和激素變化尚不清楚。本研究基于前期研究,選用2個Pb富集差異顯著的馬鈴薯品種‘青薯9號’和‘麗薯6號’,對其幼苗進行不同質量濃度Pb處理,通過分析Pb處理下馬鈴薯生長特征和內源激素變化,明確其在不同Pb質量濃度下的形態(tài)學響應及激素變化規(guī)律,為進一步揭示Pb對馬鈴薯生理特性的影響提供參考。
1 材料與方法
1.1 供試材料
以高富集品種‘青薯9號’和低富集品種‘麗薯6號’馬鈴薯脫毒苗為試材,由云南農業(yè)大學薯類研究所提供,將脫毒苗在無菌條件下剪成單葉節(jié)段,插入含20 mL固體MS培養(yǎng)基(w=3%蔗糖,pH 5.8)的組培瓶,每瓶7節(jié)莖段,以24 h為1個周期,光照16 h(光照度3 000 lx),黑暗8 h,溫度(21±2)℃的條件下培養(yǎng)30 d。選取長勢一致的脫毒試管苗移栽至帶有基質土(w=80%細碎泥炭、w=20%蛭石)的漂浮盤上(規(guī)格:18×9孔),每個漂浮盤移栽162株,基質土購買于昆明呈貢斗南園藝資材經營部。
1.2 試驗設計
將帶有脫毒苗的漂浮盤放置于含有6 L Hoadland營養(yǎng)液的塑料框內進行Pb(CH3COO)2·3H2O處理,設置5個質量濃度,分別為0、50、250、500、1 000 mg/L,以0 mg/L為對照(CK),溫度23 ℃的條件下培養(yǎng)30 d后進行幼苗形態(tài)學參數測量及內源激素測定。
1.3 測定指標及方法
1.3.1 幼苗形態(tài)學參數 每個處理隨機選取5株幼苗,將整株幼苗清洗干凈,用吸水紙擦干,通過EPSON V700掃描儀將完整幼苗圖像掃描存入計算機,采用WinRHIZO PRO 2012根系分析系統軟件(Regent Instruments,Canada Inc.)分析總根長、表面積、體積和葉面積,分析天平稱取生物量(鮮樣)質量,用卷尺和游標卡尺分別測定株高和莖粗。單株形態(tài)學參數重復測定3次。
1.3.2 葉片內源激素質量分數 稱取馬鈴薯組培苗葉片1.000 g,每處理重復3次,液氮冷凍后迅速放入-80 ℃保存,均采用酶聯免疫吸附法(ELISA),委托上海篤瑪生物科技有限公司測定赤霉素(GA3)、脫落酸(ABA)、水楊酸(SA)、乙烯(ETH)、生長素(IAA)和玉米素(ZT)的質量分數。
1.4 數據處理
試驗設置5個生物學重復,每個重復進行3次測量,采用Excel 2013進行統計、制圖,采用SPSS 21.0軟件進行方差分析和逐步回歸。
2 結果與分析
2.1 不同質量濃度Pb處理對馬鈴薯幼苗生長的影響
隨著Pb處理質量濃度的增加,供試2個品種馬鈴薯幼苗的單株生物量、株高、莖粗、葉片數和葉面積(表1)呈先增后降趨勢,與未添加Pb相比,Pb質量濃度為50 mg/L時生物量和株高最大,‘青薯9號’和麗薯6號’生物量顯著增加 11.51%和7.69%;Pb質量濃度為250 mg/L時2個品種馬鈴薯幼苗生物量和株高均開始降低,當質量濃度為1 000 mg/L時2種生物量和株高降至最低。表明低Pb質量濃度對這2個品種馬鈴薯幼苗的生長有促進作用,高Pb質量濃度對生長有抑制作用。
Pb質量濃度為50 mg/L時,2個品種馬鈴薯幼苗莖粗、葉片數和葉面積顯著高于對照。Pb質量濃度為50、250 mg/L時,‘麗薯6號’莖粗顯著高于‘青薯9號’,與CK相比,‘麗薯6號’增長 73.68%,‘青薯9號’增長22.73%,其余質量濃度下‘青薯9號’顯著高于‘麗薯6號’。說明不同Pb質量濃度處理下這2個品種莖粗、葉片數和葉面積也存在濃度效應。Pb質量濃度為50、250 mg/L時對‘麗薯6號’莖粗有促進作用。Pb質量濃度為50 mg/L時,‘青薯9號’和‘麗薯6號’總根長是CK的3.06倍和2.31倍,1 000 mg/L時開始降低,比CK處理分別降低13.44%和 43.00%;Pb質量濃度為50 mg/L時,‘青薯9號’和‘麗薯6號’根表面積分別是CK的4.92倍和 5.55倍,1 000 mg/L質量濃度下達到最小值;不同質量濃度處理后的2個品種馬鈴薯幼苗根體積較CK均有增加,Pb質量濃度為50 mg/L時增量最大,‘青薯9號’和‘麗薯6號’分別是CK的2.67倍和3.00倍;以上結果說明這2個品種馬鈴薯地下部根系的生長受Pb脅迫與地上部分規(guī)律大致一致,隨Pb處理質量濃度的增加呈先增后降趨勢。
2.2 不同質量濃度Pb處理對馬鈴薯幼苗內源激素的影響
2.2.1 馬鈴薯幼苗ABA和SA質量分數 由表2可知,隨Pb質量濃度的增加,‘青薯9號’和‘麗薯6號’的ABA質量分數均呈上升趨勢,當Pb質量濃度為1 000 mg/L時達最大值,與CK相比,‘青薯9號’ABA質量分數增加29.03%,‘麗薯6號’增加36.91%。且在任一Pb質量濃度下,‘麗薯6號’ABA質量分數均高于‘青薯9號’。不同Pb質量濃度下,2個品種馬鈴薯SA質量分數均低于CK。Pb質量濃度為50 mg/L時,2種馬鈴薯SA質量分數均降到最低,與CK相比,‘青薯9號’降幅為20.17%,‘麗薯6號’降幅為42.46%。表明不同程度Pb處理對重金屬低富集型‘麗薯6號’ABA和SA質量分數的影響強于高富集型‘青薯9號’。
2.2.2 馬鈴薯幼苗ETH和ZT質量分數 隨Pb質量濃度的增加,‘青薯9號’和‘麗薯6號’ETH變化趨勢不同。‘麗薯6號’呈現先升高后下降的變化規(guī)律,在Pb濃度為250 mg/L時,‘麗薯6號’ETH質量分數達到高峰,是CK的2.02倍,‘青薯9號’變化不大。 ZT與細胞分裂有關,2個品種馬鈴薯隨Pb處理質量濃度的增加ZT質量分數變化趨勢一致,均呈現降-升-降的趨勢。Pb質量濃度為250 mg/L時2個品種ZT質量分數降到最低,與CK相比,‘青薯9號’與‘麗薯6號’分別降低9.39%和12.47%。Pb質量濃度為500 mg/L時2個品種ZT質量分數升到最高,‘青薯9號’與‘麗薯6號’較CK分別上升8.01%和 14.93%。以上結果說明,Pb脅迫對馬鈴薯幼苗分裂生長有影響;脅迫初期重金屬低富集型馬鈴薯較高富集型馬鈴薯ETH質量分數更高。
2.2.3 馬鈴薯幼苗IAA和GA3質量分數 生長素對植物的生長調節(jié)具有雙重性,10-10mol/L是促進根系生長的最適濃度[21]。經數據轉換,本研究2個品種的IAA為26.47~38.29 pmol/L,處于促進根系生長的最適濃度。表明Pb脅迫對2個品種IAA質量分數影響不大,均處于最適宜根系生長的濃度范圍。
隨Pb質量濃度的增加,2個品種GA3質量分數均呈現下降-上升-下降-上升的波動性變化。在處理質量濃度為50 mg/L時,2個品種GA3質量分數均降到最低,與CK相比,‘青薯9號’和‘麗薯6號’降幅為12.90%和27.27%。處理質量濃度為250 mg/L時達到最大值,較CK‘青薯9號’增幅為25.81%,麗薯6號增幅為24.24%。
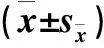
表1 不同質量濃度Pb處理下馬鈴薯幼苗生長狀況Table 1 Growth characteristics of potato seedings under different mass concentration of Pb treatments
注:不同大寫字母表示同一脅迫下不同品種間的差異顯著(P<0.05);不同小寫字母表示同一品種在不同脅迫下的差異顯著(P<0.05)。
Notes: Different capital letters indicate significant difference at 0.05 level between two varieties in each treatment;different small letters indicate significant difference at 0.05 level between different treatments within the same variety.
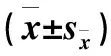
表2 不同質量濃度Pb處理下馬鈴薯幼苗內源激素的質量分數Table 2 Endogenous hormone mass fraction of potato seedings mass concentration of Pb treatments ng/g
3 討 論
3.1 不同質量濃度Pb處理對馬鈴薯幼苗生長的影響
植物在重金屬影響下會產生一系列形態(tài)學響應,但也存在濃度效應,武倩倩[21]研究表明:低劑量的土壤Pb對紅薯生長有一定的促進作用,但土壤Pb質量分數超過350 mg/kg后開始表現出抑制作用。本試驗中,低質量濃度Pb對供試馬鈴薯幼苗的生長有促進作用,高質量濃度Pb對供試馬鈴薯幼苗的生長有抑制作用。一般認為,根系生長情況可作為評判重金屬對植物生長發(fā)育的指標[22]。本試驗發(fā)現,地下部分根長、根表面積和體積與地上部分規(guī)律大致一致,隨Pb處理質量濃度的增加均呈現先增后降的趨勢,該結果與Larbi 等[13]研究結果一致。表明Pb破壞了根系細胞膜的穩(wěn)定性,幼苗通過根長、表面積和體積的響應,從而提高對Pb的耐受性,但對Pb 脅迫耐性有一定的范圍[23-24]。有研究發(fā)現同一Pb濃度對同一植物不同類型的地上、地下生長特征影響不同[25]。本試驗中對Pb脅迫的‘青薯9 號’和‘麗薯6號’的形態(tài)特征變化趨勢基本一致,但具體指標的變化量仍存在品種差異,質量濃度為50 mg/L時,‘麗薯6號’根表面積、根體積較CK的變化量比‘青薯9號’更明顯。這可能與‘麗薯6號’和‘青薯9號’對Pb積累量不同有關,但具體的調控機制還需結合馬鈴薯對Pb的分配、轉移途徑做進一步研究。
3.2 不同質量濃度Pb處理對馬鈴薯幼苗內源激素的影響
ABA通常被認為是信號轉導物質,在植物受到非生物脅迫時發(fā)揮重要作用[26]。ABA可作為擬南芥響應重金屬脅迫的潛在調節(jié)者,其濃度隨重金屬脅迫濃度的增加而增加[27]。SA是第一個被證明在重金屬毒性生物應激反應中發(fā)揮作用的內源性植物生長調節(jié)劑[28]。本試驗中,Pb處理會導致馬鈴薯幼苗中內源激素ABA增加,不同程度Pb處理對重金屬低富集型‘麗薯6號’ABA和SA質量分數的影響強于重金屬高富集型‘青薯9號’,這一結果可能與ABA可誘導氣孔關閉從而抑制蒸騰流,阻礙金屬離子的向上運輸,從而使‘麗薯6號’重金屬積累量較低有關[29]。乙烯可促進莖的橫向生長,重金屬脅迫下也會出現莖的增粗[30]。本試驗重金屬低富集型‘麗薯6號’中ETH質量分數為250 mg/L時達到最大,且高于‘青薯9號’,此質量濃度下的‘麗薯6號’的莖粗大于‘青薯9號’,說明‘麗薯6號’通過ETH質量分數的增加導致莖粗增加,以應對重金屬脅迫。IAA對植物的根、芽、莖的生長有其最適濃度范圍[21],本研究中任意Pb質量濃度下2個品種IAA質量分數均處于促進根系生長的最適濃度。GA3可促進植物生長,打破休眠,提高抗逆性等,在植物應對非生物脅迫的作用越來越明顯[31-32]。本研究中,不同Pb質量濃度下GA3質量分數的變化趨勢表現為下降-上升-下降-上升的波動性變化,反映了植物激素GA3在應對重金屬脅迫時與其他激素共同作用有關。有研究表明赤霉素在調節(jié)激素動態(tài)平衡的基礎上對植株進行調控[33-34]。這一結果與朱珍[35]的研究結果相似,該推斷是否合理還需進一步驗證。
4 結 論
不同脅迫程度下,重金屬富集特性不同的品種會啟動相應的形態(tài)以應對Pb脅迫,使馬鈴薯形態(tài)特征發(fā)生變化,低質量濃度Pb對馬鈴薯幼苗生長有促進作用,高質量濃度Pb對馬鈴薯幼苗生長有抑制作用。
不同質量濃度Pb處理下馬鈴薯幼苗內源激素變化是一個比較復雜的過程, ABA、SA、ETH、IAA、ZT、GA3共同參與了馬鈴薯在重金屬脅迫下的協調機制。不同程度Pb處理對重金屬低富集型‘麗薯6號’ABA和SA質量分數的影響強于重金屬高富集型‘青薯9號’。Pb脅迫對2個品種IAA質量分數影響不大,均處于最適宜根系生長的濃度范圍。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
