短期增溫對長江源區高寒沼澤草甸植物-土壤C,N化學計量及季節變化特征的影響
2019-12-20 01:27:28劉永萬尹鵬松張景然
草地學報 2019年6期
劉永萬, 白 煒, 尹鵬松, 馮 月, 張景然
(蘭州交通大學環境與市政工程學院, 甘肅 蘭州 730070)
近年來,由于工業的發展以及人類生活水平的快速提升,CO2及其他溫室氣體劇增,由此導致的全球氣候變暖已引起世界各國的廣泛關注[1]。據政府間氣候變化委員會第五次評估報告顯示,在過去130年來全球平均氣溫升高了0.85℃[2]。青藏高原作為世界上海拔最高的地區,因其嚴酷而脆弱的環境條件,對氣候變化十分敏感[3]。王朋嶺等[4]研究發現青藏高原地區平均氣溫以每十年0.4℃的速率增加,且青藏高原地區氣溫增加幅度明顯高于同期北半球平均增溫幅度。氣溫的持續升高導致青藏高原產生了一系列的生態問題,凍融周期的變化、氣候暖干化趨勢等直接影響著生態系統的物質循環,進而改變植物-土壤系統的化學計量特征。
碳(C)是植物有機質的主要成分,約占植物干重的1/2[5]。生態系統中植被的C主要來源于植物通過光合作用固定大氣中的CO2;C含量反映了植物的光合碳同化能力,是植物C儲量的一種度量。氮(N)是蛋白質、葉綠素、核酸等的一個重要組成部分,是植物生長所必需的重要營養元素之一。C和N作為植物生長所必需的養分元素,雖然其含量在特定植物種的特定生長階段具有一定的變異性,但生物體具有保持其自身C,N元素組成相對穩定的能力[6],而這種相對穩定狀態容易受到外界環境脅迫而被破壞[7],從而影響植物的生長、土壤C積累動態和N養分限制格局[8]。
到目前為止,基于增溫對我國高海拔凍土區域高寒草地方面的研究多集中在植物群落組成、物種多樣性和生物量等[3,8-9],對其化學計量學方面研究較少,且得出的結果各不相同。羊留冬[7]等人模擬氣候變暖發現貢嘎山峨眉冷杉幼苗在增溫處理后C平均含量和C/N均無顯著變化,N含量顯著降低;江肖杰[10]等人則發現增溫并沒有改變植物葉片的C,N含量,卻使其土壤N含量、葉片C/N呈增加趨勢;彭阿輝[6]等研究發現增溫增加了植物地上部分C/N,但對不同植物C,N含量影響并不一致。上述研究多集中于植物領域,而對植物-土壤C,N營養元素傳遞與調節機理的研究還比較薄弱,對增溫處理下,高寒草地植物、土壤化學計量比之間的關系及其規律更是鮮有報道。沼澤草甸是高寒草地生態系統的三大主要類型之一,下墊面為季節性凍土或永久凍土,溫度變化會對土壤的物理性質、水分含量以及養分狀況產生巨大影響,從而影響植物生長及其C,N含量。藏嵩草(Kobresiatibetica)是青藏高原風火山地區高寒沼澤草甸的優勢種,其生長狀態及C,N含量的改變都會對生態系統造成巨大的影響。因此,本研究以青藏高原腹地長江源風火山地區高寒沼澤草甸優勢種藏嵩草為研究對象,設立增溫樣地,比較分析不同增溫幅度下長江源區草地植物與土壤C,N生態化學計量比的變化與相互關系,為揭示植物-土壤C,N化學計量特征對溫度變化的響應和調控機制提供科學依據。
1 材料與方法
1.1 試驗地位置及植被概況
研究區域位于高寒沼澤草甸分布較為典型的青藏高原腹地風火山地區,海拔4 600~4 800 m,相對高差200~300 m,南北寬約40 km,整體呈北西西向延伸,屬于可可西里山系的東南支[11]。屬于高原大陸性氣候區,寒冷干燥,凍結期 9月至翌年4月,年平均氣溫—5.3℃,年均降水量270 mm,且80%的降雨發生在植物的生長季(5-9月),年均蒸發量1 478 mm[12-13]。選取的樣地位于34°43′43.9″ N,92°53′34.1″E,海拔4 754 m,其植物群落主要由濕中生、濕生多年草本植物群落構成。優勢種為藏蒿草(Kobresiatibetica)、小蒿草(Kobresiahumilis)、針茅(Stipacapillata)、羊茅(Festucaovina)和粗喙苔草(Carexscabrirostris)等高寒植物。
1.2 試驗設計
2015年7月,在研究區域內,隨機選取植被相對分布均勻一致的區域,設置2種模擬增溫處理:T1,增溫 1.5~2.5℃;T2,增溫 3~5℃。以自然樣地為對照(CK),每種處理設置5個重復。
模擬增溫的方式采用采用國際凍原計劃(International Tundra Experiment,ITEX) 所采用的被動式增溫法——開頂式氣室法(Open-top chambers,OTC),建造開頂式小暖室,小室為正六邊形圓臺狀結構,六面均為有機玻璃,透光率95%,小室的高度分別為40 cm和80 cm,并保證小室的上開口均為60 cm,圓臺的斜邊與地面的夾角均為60°。同時,采用EC-TM溫濕度傳感儀(Decagon Decices Inc.,Pullman,WA)記錄空氣溫度與濕度。
1.3 研究方法
1.3.1取樣方法 于2018年的6月(返青期)、8月(結實期)和9月(枯黃期),在每個處理樣地內各隨機選取10株藏嵩草,連同根一起取得完整的植株樣品,將其周圍枯枝和凋落物收集并分別裝進信封袋,做好標記。同時采集植物根部周圍層0~10 cm的土壤裝入自封袋,做好標記。帶回實驗室的植物洗掉根部泥土后快速陰干,將地上、地下分開裝袋,做好標記。于65℃烘箱內烘干至恒重,研磨、過篩用于有機碳和全氮的測定。同時,將土樣陰干、研磨、過篩用于有機碳和全氮的測定。
1.3.2測定項目與方法 有機碳測定采用重鉻酸鉀外加熱法;全氮測定采用半微量開氏法,經EHD20型Digi Block智能樣品消解爐(萊伯泰科有限公司)消解后,由半微量蒸餾裝置蒸餾,后經硫酸滴定測定[14]。
1.3.3數據分析 使用Origin2018進行數據制圖,在SPSS 25中對各項數據進行整理和分析。采用單因素方差分析(One-Way ANOVA)對植物-土壤C,N含量及其化學計量比進行差異顯著性檢驗,并采用Sparman相關系數法評價不同指標之間的相關性。
2 結果與分析
2.1 模擬增溫的效果
由于溫室的阻擋作用,增溫棚室內風速降低,空氣湍流減弱,使熱量不易散失,加之玻璃纖維被太陽輻射中紅外線穿透的能力較好[15],所以室內溫度升高。在整個植物生長季內,沼澤草甸的平均氣溫分別比CK提高了3.1℃(T1)和5.5℃(T2),其中7月月平均氣溫達到最大值。根據以往研究,青藏高原地區平均氣溫以每十年0.4℃的速率增加[4],就這種趨勢而言,本研究采用的增溫系統在未來80~140 a,增溫效果都是顯著的(圖1)。
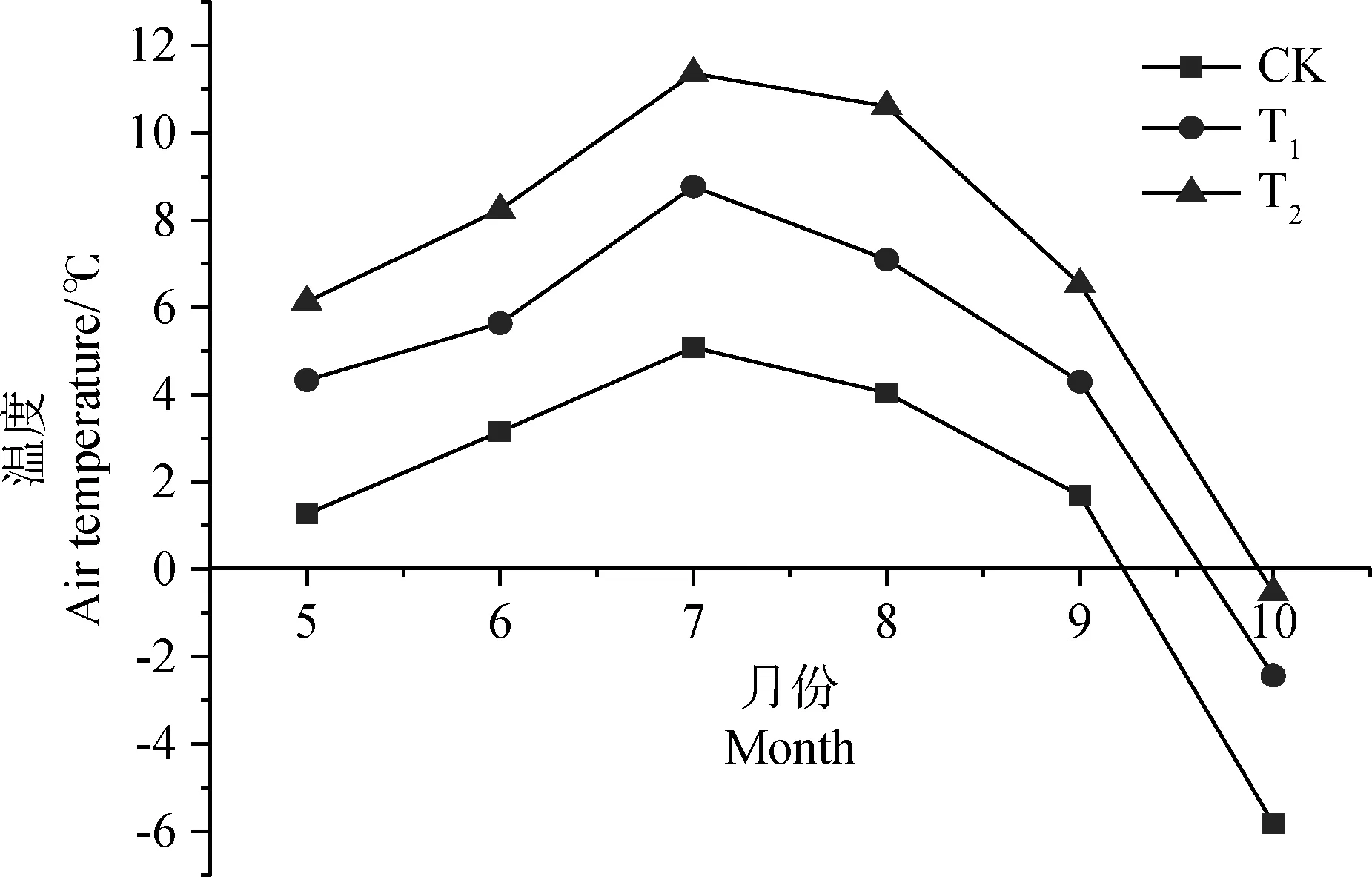
圖1 5-10月月平均氣溫Fig.1 Average temperature from May to October注:CK,自然樣地;T1,增溫 1.5~2.5℃;T2,增溫 3~5℃,下同Note:CK,natural sample;T1,warming 1.5~2.5℃;T2,warming 3~5℃,the same as below
2.2 增溫對植物地上部分C,N的影響
增溫使得植物地上部分C含量減少,但只有結實期T2處理下差異顯著(P<0.05),T1處理組相比CK減少了0.1%~3.2%,T2處理組相比CK減少了0.8%~9.8%,表明增溫降低了植物地上部分C含量,但低增溫對其影響并不顯著,隨著增溫幅度的增加,會加劇這種效果。對增溫處理后植物地上部分N含量來說,增溫顯著降低了返青期植物地上部分N含量(P<0.05),增加了結實期植物地上部分N含量,對9月枯黃期植物地上部分N含量并無明顯影響(圖2)。
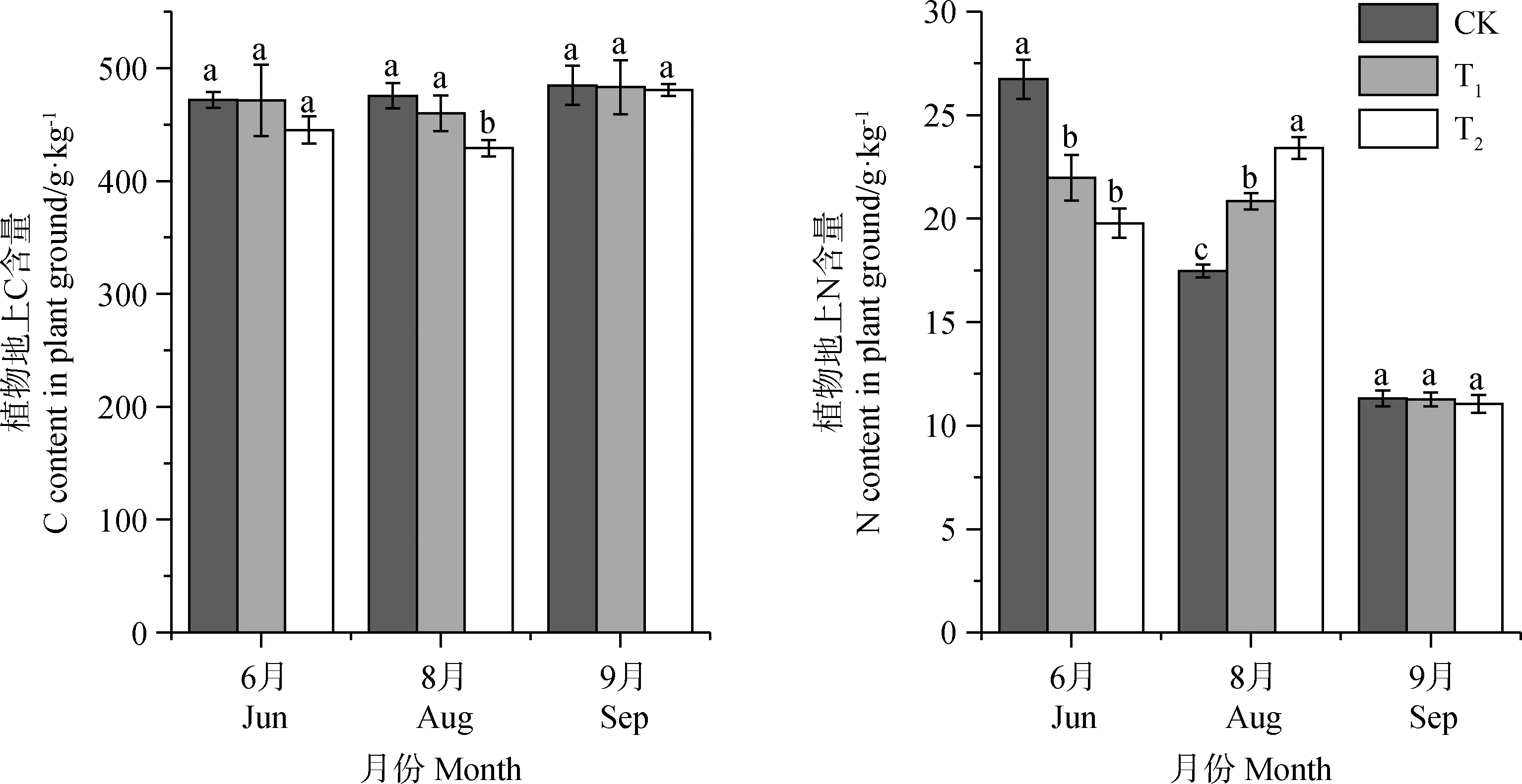
圖2 不同增溫幅度對植被地上部分C,N含量的影響Fig.2 The effect of different temperature increase range on the aboveground part of C and N content in different seasons注:不同小寫字母表示不同增溫幅度下的差異顯著性(P<0.05),下同Note:Different lowercase letters indicate significant differences between different temperature increase range at the 0.05 level,the same as below
2.3 增溫對植物枯落物C,N的影響
增溫處理下6月返青期植物枯落物C含量顯著增加,T1處理下增加了5.2%,T2處理下增加了15.8%。而8,9月枯落物C含量明顯減少,T1處理組相比CK減少了8.2%~10.1%,T2處理組相比CK減少了1.7%~5.9%。對于增溫處理后的枯落物N含量來說,增溫顯著降低了其N含量,T1處理組相比CK減少了9.2%~19.8%,T2處理組相比CK減少了6.1%~28.1%(圖3)。
圖3 不同增溫幅度對植被枯落物C,N含量的影響Fig.3 The effects of different temperature increase ranges on the content of C and N in litter of different seasons
2.4 增溫對植物根系C,N的影響
增溫在6月返青期對植物根系C含量影響并不顯著。T1和T2處理下8,9月根系C含量均顯著低于對照(P<0.05),T1處理下其根系C含量相比CK降低了13.2%~25.1%;T2處理組相比CK降低了19.4%~25.7%,表明增溫處理明顯減少了植物根系C含量,但在返青期表現并不顯著(圖4)。
植物根系N含量對增溫的響應有所不同,T1處理組顯著降低了8月結實期植物根系N含量(P<0.05),使其減少了14.6%,對其他時期植物根系N含量影響并不顯著。T2處理組在6月返青期、8月結實期顯著增加了植物根系N含量(P<0.05),使其增加了10.1%~13.1%,在9月枯黃期則有一定程度的降低,但這種影響幅度較小,統計結果也表明差異并不顯著。表明低增溫減少了結實期植物根系N含量,對其他時期并無顯著影響;高增溫則促進了植物根系N含量。
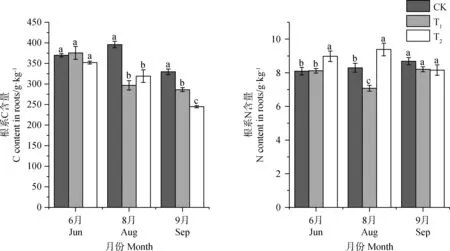
圖4 不同增溫幅度對植被根系C,N含量的影響Fig.4 The effect of different temperature increase range on the content of C and N in roots of different seasons.
2.5 增溫對表層土壤C、N的影響
與CK相比,T1處理組明顯促進了土壤表層有機碳含量,使其增加了12.2%~34.7%,T2處理同樣促進了土壤有機碳含量,但返青期與CK之間差異并不顯著,T2處理組相比CK使之增加了3.6%~214.0%。表明增溫對土壤表層有機碳含量有明顯的促進作用(圖5)。
增溫處理后,就土壤表層全氮含量來言,T1,T2處理下返青期土壤全氮含量均顯著低于CK(P<0.05),分別減少了17.2%和18.7%;T1處理顯著促進了8,9月土壤全氮含量(P<0.05),使其相比CK增加了17.2%~32.0%;T2處理同樣表現為促進作用,但9月并不顯著,使其增加了0.17%~99.2%。表明增溫處理減少了返青期的土壤表層全氮含量,其他時期使土壤表層全氮含量明顯增加。
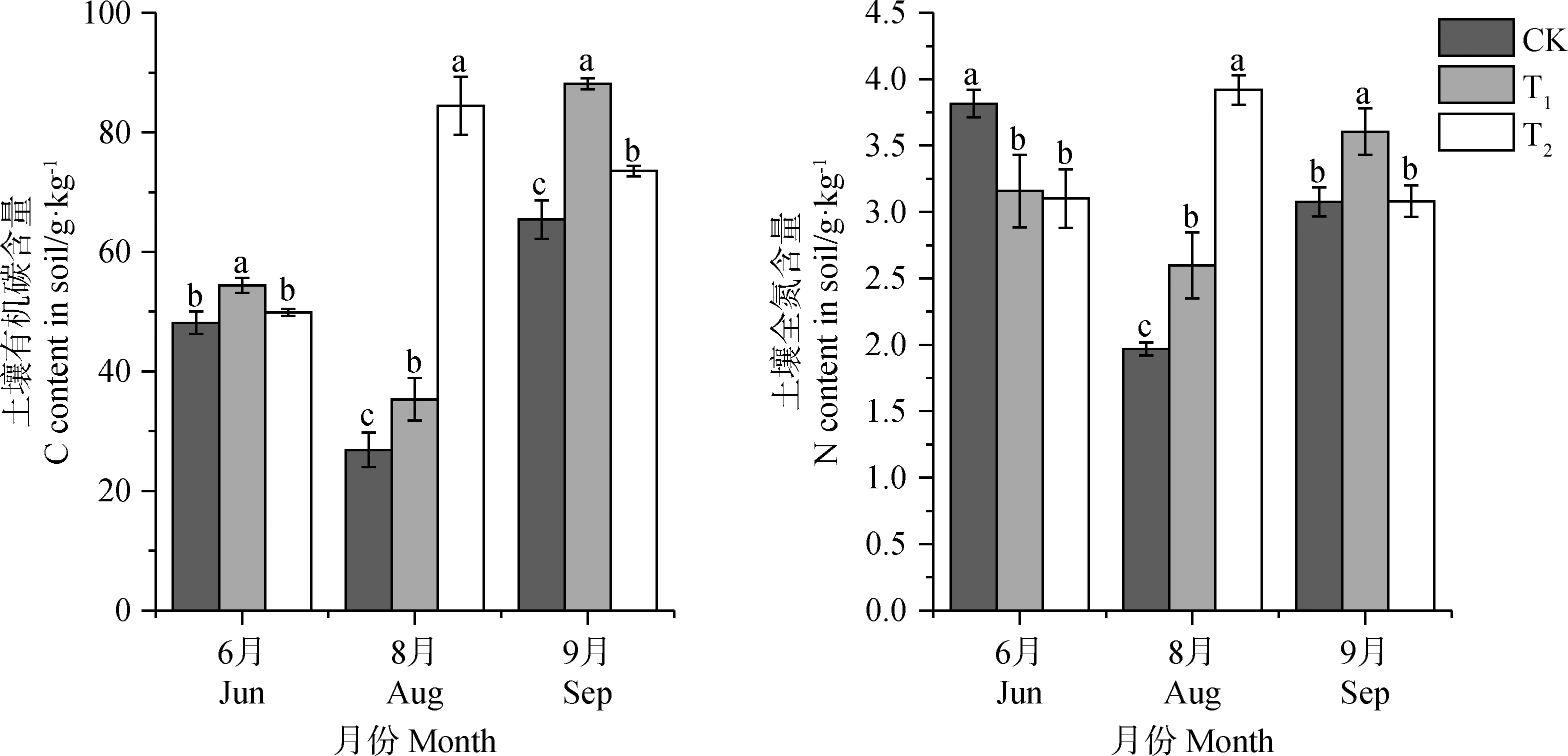
圖5 不同增溫幅度對土壤有機碳、全氮含量的影響。Fig.5 The effect of different temperature increase ranges on soil organic carbon and total nitrogen content in different seasons
2.6 增溫處理下植物器官及土壤C/N的比較
增溫處理對6月植物地上部分C/N表現為顯著的促進作用(P<0.05),但8月顯著低于對照(P<0.05),9月有一定的促進作用但差異并不顯著。植物枯落物C/N明顯增加,但其中8月T2處理及9月T1處理下差異并不顯著,T1處理組相比CK增加了1.6%~14.5%,T2處理組相比CK增加了0.5%~36.7%。而對于增溫后的植物根系C/N來說,除6月T1處理下差異并不顯著外(P>0.05),其他不同時期、不同處理下的植物根系C/N顯著降低(P<0.05)。T1處理下根系C/N降低了8.1%~12.2%,T2處理組相比CK使根系C/N降低了13.9%~28.8%,表明增溫處理減少了植物根系C/N。表層土壤C/N與之相反,除8月T1處理下差異并不顯著外(P>0.05),其他不同時期、不同處理下的土壤C/N明顯增加。T1處理組相比CK增加了0.8%~37.6%,T2處理組相比CK增加了14.4%~57.8%。說明在整個生長季內增溫明顯影響了植物對氮素的利用、枯落物的分解以及土壤C,N供給和儲存(表1)。

表1 不同處理對各組分C/N的影響Table 1 Effect of different treatments on C/N of each component
注:不同小寫字母表示同一指標在同一時間下不同處理差異顯著(P<0.05)
Note:Different lowercase letters indicate that the same index differs significantly at the same time at the 0.05 level
同時,對不同增溫處理下的植物與土壤C,N及其化學計量比之間的相關性分析得出:植物地上部分全氮和植物根系有機碳、植物根系C/N,植物地上部分C/N和土壤C/N,根系有機碳和枯落物全氮,根系C/N和枯落物全氮,土壤有機碳和土壤全氮均存在顯著的正相關關系(P<0.01);植物地上部分全氮和枯落物全氮、土壤C/N,植物根系有機碳和枯落物C/N、土壤有機碳、土壤C/N,植物根系C/N和土壤有機碳、土壤C/N均存在顯著的負相關關系(P<0.01);枯落物有機碳和植物根系有機碳、根系C/N間均存在顯著的正相關關系(P<0.05);植物地上部分有機碳和植物地上部分全氮、植物根系全氮,植物地上部分C/N和枯落物全氮、植物根系有機碳、植物根系C/N,枯落物有機碳和土壤有機碳、土壤C/N,植物根系C/N和土壤全氮,土壤C/N和枯落物全氮之間均存在顯著的負相關關系(P<0.05)(表2)。
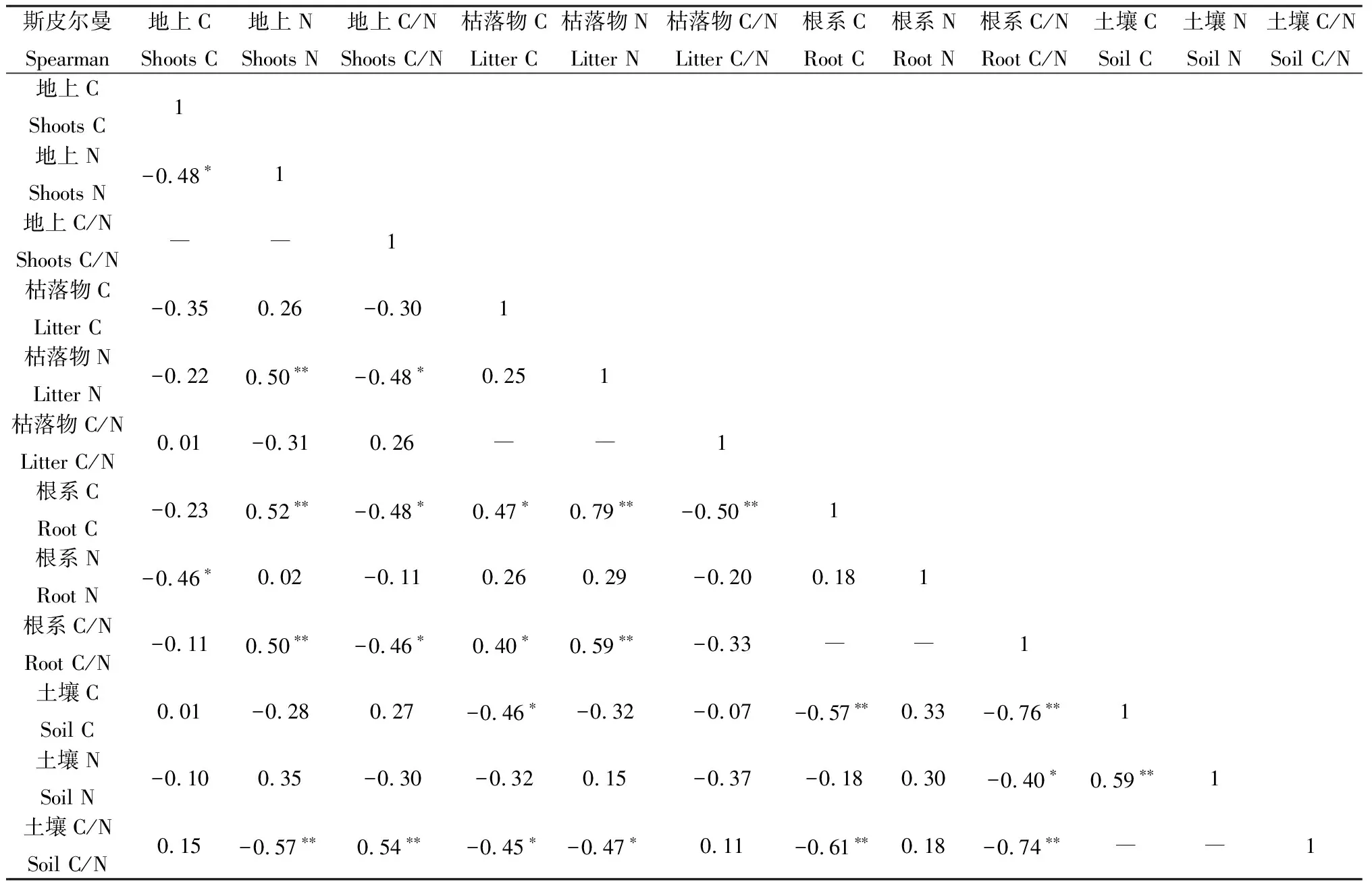
表2 植物、枯落物、土壤間C,N的相關性Table 2 The correlations of plants,litter and soil C and N
注:*,**分別代表顯著性水平小于0.05和0.01;“―”表示存在自相關關系,不宜進行相關分析
Note:*,** represent the significance difference at the 0.05 and 0.01,respectively;"―" indicates that there is an autocorrelation relationship,and it is not appropriate to conduct correlation analysis
3 討論
3.1 模擬增溫對植物C,N的影響
本研究發現在整個生長季內植物地上部分碳含量除結實期T2處理下顯著降低外,其他不同時期處理下并無顯著變化,而根系碳含量除返青期外顯著減少。這可能是由于植物在生長旺盛期,為了吸收更多的水分和養分,大量的碳水化合物被輸送到根部以促進根系的生長,造成其根部碳水化合物的積累小于根系生長,從而導致根系C濃度顯著降低。與本研究結果不同的是,Yang[16]等通過模擬增溫對青藏高原植物養分分配的影響試驗中發現,增溫和干旱會降低植物地上部分C含量,增加植物根系C含量;而石福孫[17]等對川西北高寒草甸進行模擬增溫試驗卻發現增溫對植物地上部分C含量影響并不顯著,使其根系C含量明顯增加。由此可見,植物C含量對增溫的響應并不一致,原因可能與植被類型的不同、所處季節的不同、增溫方式、增溫時間以及環境因素等有著密切聯系。
本研究中返青期植物地上部分N濃度隨溫度升高而降低,結實期N濃度隨溫度升高而增加,枯黃期N濃度降低但差異不顯著。可能是由于增溫環境下,植物的生物量和氮利用效率均有所提高,從而降低了植物組織的N濃度[18-19];同時,由于結實期溫度升高,光合作用增強,植物生長迅速,有機質積累量增加,增溫加劇了微生物從有機物質中分解氮的能力[20],植物對土壤中可利用氮的吸引也隨之增加,從而導致結實期地上活體N濃度隨溫度升高而增加。
地下根系N濃度與地上不同,T1處理下除結實期顯著降低外(P<0.05),其他時間并無明顯變化;在T2處理下返青期、結實期均顯著增加(P<0.05)。可能是因為7,8月環境溫度達到最大值,增溫處理下溫度超過了植物生長的適宜溫度,使其處于半干旱條件下,植物除了進行光合作用,還需要增加氮和磷的吸收量,用以增加自身抗旱能力[21]。低增溫處理下,處于8月結實期的植物根系向更深生長,以便吸收所需要的水分和營養物質帶來了稀釋效應,植物根對營養元素的吸收趕不上細胞膨脹的速率,使得根系N元素降低[22];而高增溫處理下,表土的過于干旱對根的伸長產生強烈的機械阻礙作用[23],因此其N濃度高于對照;Yang等[16]研究也發現由于可溶性蛋白質的積累,干旱條件下植物N濃度顯著增加。
此外,植物葉片C/N的大小可以表示植物吸收單位養分元素含量所同化C的能力,在一定程度上反映了植物體內養分元素的利用率[6]。有研究表明,增溫通過增加植物生產能力以及植物的生物量來影響它們對營養的吸收,會降低植物中的C/N[24]。還有一些研究則表明,在氣候寒冷的地區,增溫通過增加生物量的生產,反而會使C/N有所增加[25]。Yang等[26]對青藏高原高山篙草的研究表明,增溫降低了根部的C/N,提高了植物葉片的C/N,這意味著植物可能會以提高N的利用效率來應對未來的氣候變暖。本研究發現增溫降低了不同生長季植物根系C/N,且除結實期植物地上部分C/N低于CK外,其他不同時期、不同處理的植物地上部分C/N均高于對照,表明增溫條件下返青期、枯黃期植物能夠更有效地利用N素,增加其利用效率。結實期植物地上部分C/N低于CK可能是由于8月處于生長旺季,增溫處理下溫度升高,光合作用增強,植物生長迅速,有機質積累量增加,微生物從有機物質中分解氮的能力加劇,對土壤中可利用氮的吸引增強,從而導致地上部分C/N降低。增溫處理后植物地上部分、根系C/N均發生改變,說明溫度升高將會導致植株C/N改變,從而影響其對養分的利用效率。
3.2 模擬增溫對枯落物C,N的影響
植物在自身的生長過程中,通過光合作用產生有機物,并通過枯落物的分解以及根系的分泌作用將養分歸還于土壤中[27]。因此,枯落物及其分解過程是聯系植被和土壤的紐帶,它對土壤植物養分自身的需求和調節起著關鍵的作用[28],而微生物的分解活動與待分解物的C/N密切相關[29]。有研究表明,當枯落物N含量升高或C/N值降低會加快枯落物的分解速率,加速養分循環[30]。本研究發現,不同季節的植物枯落物有機碳對增溫的影響并不一致,增溫增加了返青期植物枯落物有機碳含量,減少了結實期、枯黃期枯落物有機碳含量,枯落物全氮含量在整個生長季內明顯降低,枯落物C/N明顯增加,且不同增溫幅度對其影響并不一致,說明植物氮素在不同季節不同增溫條件下發生了不同程度的釋放和遷移,溫度升高將會導致枯落物C/N改變,從而影響沼澤草甸生態系統的分解速率。安卓等[31]研究發現,在環境中養分供應短缺時,植物以提高養分重吸收率為其養分利用的主導方式適應環境脅迫,這恰好與本研究結果相互印證。
3.3 模擬增溫對土壤C、N的影響
土壤是草地生態系統的重要組成部分,為植物提供必需的營養和水分,是各種物質能量的轉化場所[32],其養分含量的高低影響著植被生物量生產及其分配[33]。本研究表明,溫度升高增加了整個生長季0~10 cm土壤表層中的有機碳含量,可能是因為溫度升高雖然加快了土壤有機碳的分解速率[34],但同時溫度升高也增加了植物生產力,地表植物枯落物、根系以及根系分泌物補充了土壤中的有機質[35],從而使得土壤有機碳含量增加。
就增溫處理后的表層土壤全氮來看,如圖8所示,增溫降低了返青期0~10 cm土壤表層中的全氮含量,增加了結實期、枯黃期0~10 cm土壤表層中的全氮含量。這可能是由于返青期植物生物量小,增溫促進植被生長,需要大量的蛋白質和核酸來滿足植物逐漸增加的生長速率,從而吸收利用大量的土壤氮素[23],導致土壤氮的減少。而土壤氮素在夏季則以氮素固持為主[36],使得土壤表層全氮含量增加。亓偉偉[37]等研究也表明,短期溫度增加有利于有機質分解,使更多有機氮分解為植物可利用的礦化氮,促進土壤表層全氮含量增加。
研究還發現增溫明顯增加了整個生長季0~10 cm土壤表層中的C/N,使之從12.63~20.87增加到13.76~24.50,且對照和增溫處理下的土壤C/N均明顯高于全國范圍(11.9±0.4)內C/N值的平均值[38],可以發現風火山地區高寒沼澤草甸土壤在整個生長季內均處于氮素限制狀態,增溫促進植物生長,加劇了這種限制。
3.4 植物—土壤C,N化學計量比之間的關系
土壤作為植物養分的主要來源[33],對于植物C,N含量具有十分重要的影響;植物枯落物及其分解速率又反過來影響土壤C,N含量。通過分析植物地上部分、枯落物、根系以及土壤C,N化學計量比之間的相關性發現,枯落物C,N含量與植物地上部分相關,土壤養分含量受枯落物限制,根系C,N含量又與土壤養分含量有關,植物地上部分C、N含量又與根系C、N含量有關,生態系統內部C元素和N元素在植物地上部分、枯落物、根系與土壤之間實現了運輸和轉換。植物根系從土壤中吸收養分供植物生長,地上部分通過光合作用合成有機物質,又以枯落物分解的形式將養分歸還于土壤或損失[39],從而使得不同季節不同增溫處理下C元素和N元素地上部分>枯落物>根系>土壤。
4 結論
通過在青藏高原腹地風火山地區高寒沼澤草甸開展的模擬增溫試驗可知,無論地上植物、枯落物還是地下根系,其C,N含量與土壤C,N含量之間均存在顯著相關(P<0.05)或極顯著相關(P<0.01);同時發現該研究區土壤處于氮限制狀態(C/N>12),整個生長季內增溫和對照處理下的植物生長均受N素限制,增溫在一定程度上促進植物生長,加劇了該研究區氮素的缺乏狀況,從而導致植物對C,N的利用率和枯落物的分解速率改變,土壤C/N明顯增加。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
