胡楊不同發育階段葉片光合作用及其光響應特征
2019-12-24 06:47:18單凌飛丁穎慧王雙蕾張默實韓航石莎馮金朝
生態科學 2019年6期
單凌飛, 丁穎慧, 王雙蕾, 張默實, 韓航, 石莎, 馮金朝
胡楊不同發育階段葉片光合作用及其光響應特征
單凌飛, 丁穎慧, 王雙蕾, 張默實, 韓航, 石莎, 馮金朝*
中央民族大學生命與環境科學學院, 北京 100081
胡楊()為第三紀孑遺植物, 在個體的不同發育階段形成了獨特的異形葉, 具有較強的抗旱性和耐鹽性, 是額濟納綠洲生態系統的主要建群種, 具有重要的研究價值。以內蒙古額濟納旗胡楊林自然保護區胡楊為研究對象, 通過實地調查觀測, 分析了胡楊幼苗期、幼樹期和成熟期葉片的形態變化規律及其性狀特征, 利用Li-6400XT便攜式光合作用系統測定了胡楊幼苗期、幼樹期和成熟期不同形態葉片的光合作用光響應過程, 并采用直角雙曲線修正模型對其進行了擬合分析。結果表明: (1)在胡楊生長發育過程中, 其葉片形態性狀表現出明顯的變化規律。隨著胡楊樹齡的增長, 其葉片的長/寬比值、比葉面積和葉片厚度逐漸減小, 而葉干物質含量、葉面積和葉干重數據增大。(2)胡楊成熟期闊卵圓形葉具有較高光合活性, 通過提高Pmax以增強光能利用率, 使其在干旱荒漠區高溫、強光環境下仍能保持較高的光能轉化效率與光合速率; 幼樹期條形葉具有較寬的光能利用區間, 能夠在弱光環境下保持較高的光合速率; 幼苗期條形葉具有較低的與相對較高的LCP、LSP及Pmax, 主要歸因于其較高的比葉面積, 高效光捕獲面積使其具有較高的光合速率; 上述結果是胡楊在不同光環境下形成的適應性反應和生理生態機制。(3)直角雙曲線修正模型適于胡楊不同形態葉片光合生理參數的定量研究。
胡楊; 葉片形態; 光合作用; 光響應; 直角雙曲線修正模型
0 前言
光合作用是植物將光能轉換為可用于生命過程的化學能并進行有機物合成的復雜生物物理化學過程[1], 而植物凈光合速率與光合有效輻射之間的定量關系是揭示植物光合生理過程對環境響應的基礎[2, 3]。光合作用光響應曲線的測量與模擬可獲得植物光合作用相關的最大凈光合速率(Pmax)、表觀量子效率()、光補償點()、光飽和點()和暗呼吸速率(R)等重要光合參數, 有助于闡明植物光合機構運轉狀況、不同生境的光合能力及適應規律[4]。Blackman于1905年提出了第一個光合作用對光響應模型, 之后許多學者建立了不同的光響應模型, 常用的有直角雙曲線模型[5]、非直角雙曲線模型[6]、指數模型[7, 8]和直角雙曲線修正模型[4, 9]等。前3種模型擬合的光響應參數與實測值差異較大, 難以準確模擬發生光抑制時的響應過程[4, 7, 10-13], 僅適用于擬合正常水分下植物光響應過程和響應參數[12, 13], 且無法適應擬合各種生境的不同植物。同時, 人們對不同光響應適用條件、范圍及擬合出的指標、參數均缺乏深刻的理解, 導致光響應模型選擇時缺乏可信度[10-13]。葉子飄等[10]構建的直角雙曲線修正模型克服了傳統模型的局限性, 能夠較準確地擬合各種生境植物光響應過程及其特征參數[4, 10-13]。目前, 直角雙曲線修正模型應用于冬小麥和不同土壤水分下山杏、油松、酸棗、沙棘光合作用的光響應和CO2響應模擬[10-13], 取得較好的效果, 但對于極端干旱荒漠區植物的光合作用是否適用尚需深入研究。
胡楊()是楊柳科(Salicaceae)楊屬()中古老和珍貴的荒漠樹種, 具有防風固沙、保護綠洲、維持荒漠區脆弱環境生態平衡的功能, 被確定為我國首批388種珍稀瀕危漸危種[14]。在自然條件下, 胡楊葉片在幼苗期全部為條形葉; 隨著樹齡增長, 在同一植株上會同時出現條形葉、卵圓形葉和闊卵圓形葉等不同形態的葉片(異形葉), 這是胡楊對干旱環境長期適應的結果[14, 15]。目前, 關于胡楊異形葉的光合特性、葉綠素熒光特性、水分生理特性等[14-19]方面已有廣泛研究, 但有關胡楊不同發育階段異形葉光合作用-光響應特性研究還較少, 其光合生理參數與光照強度的定量關系尚不清楚。本文以內蒙古額濟納旗胡楊林自然保護區的胡楊為研究對象, 實地觀測胡楊不同發育階段葉片的形態性狀與變化規律, 研究胡楊葉片的光合作用-光響應過程, 探索直角雙曲線修正模型在擬合胡楊光響應過程與特征參數的適用性, 旨在闡明胡楊生長發育過程中不同形態葉片適應強光、高溫荒漠環境的光合生理機制及生存策略, 為天然胡楊林保護與綠洲退化環境的生態恢復提供理論依據。
1 材料與方法
1.1 研究區域
研究區位于內蒙古自治區額濟納旗胡楊林自然保護區, 其地理位置為101°14′14.385″E, 42°1′ 11.441″N, 海拔996 m。該地區氣候極端干旱, 屬典型溫帶大陸性干旱氣候, 主要特點為干旱少雨, 蒸發量大, 溫差較大, 風大沙多。年平均氣溫8.5 ℃, 平均最低氣溫-12 ℃, 平均最高氣溫36.3 ℃; 年平均降水量38.7 mm, 蒸發量高達3706 mm; 日照時數3426.4 h, 無霜期146 d, 盛行風向為西北風。土壤類型為沙壤土, 地下水埋深為0—3 m。
1.2 試驗設計
試驗于2018年8月上旬進行, 在晴朗天氣條件下選擇生長狀況良好的胡楊幼苗(基徑1—1.5 cm)、幼樹(基徑5—8 cm)、成樹(胸徑30—35 cm)各3株進行觀測研究。其中胡楊幼苗葉片為條形葉(A1), 幼樹葉片為條形葉(A2)和披針形葉(A3), 成樹葉片分別為披針形葉(A4)卵圓形葉(A5)和闊葉(A6)。觀測時, 選取向陽枝條上部的第3、4位功能葉作為測定對象, 將枝條剪下迅速插入水中, 再從下部開始用枝剪剪去10—20 cm, 進行葉片光合作用的離體測定。光合測定完成后, 將葉片剪下, 用濕潤的濾紙包裹放入冰盒帶回室內, 進行葉片形態性狀測定分析。
1.3 葉片形態性狀測定
將葉片放在平整的A4紙上, 用數碼相機拍照, 使用Image J軟件處理葉片照片, 進行葉片長度、寬度和葉面積計算。葉片厚度的測定采用游標卡尺測量法, 選用精度為0.005 mm的游標卡尺, 每個葉片均沿主脈方向, 在距離主脈兩側約1 mm處各均勻選取3個測量點進行測定, 取平均值。采用電子天平(精度為0.001 g)測量葉片鮮重。上述測定完成后, 將葉片至于烘箱內, 80 ℃下烘干至恒重, 測定葉片干重。葉片干物質含量和比葉面積計算如下:
干物質含量()=葉片干重()/葉片鮮重()(1)
比葉面積()= 葉片面積()/葉片干重() (2)
1.4 光響應曲線測定
實驗觀測在上午進行, 先在自然光下誘導1.0—1.5 h, 之后使用Li-6400XT便攜式光合儀(Li-Cor, Inc, USA)進行測定。其中氣體流速控制為500 μmol·s-1, 葉室CO2濃度控制為400 μmol·mol-1, 葉室溫度控制為25℃; 利用6400-02B LED紅/藍光源提供不同光合有效輻射(PAR), 光照梯度設定為2000、1800、1500、1200、1000、750、500、300、150、100、50和0 μmol·m-2·s-1; 每種形態葉片處理設3次重復, 實地測定葉片光合作用對光合有效輻射的響應曲線, 由光合儀自動記錄葉片光合速率(P)等光合生理參數。
根據實際觀測數據, 繪制胡楊不同形態葉片的光合作用-光響應(n-)曲線, 確定葉片的最大凈光合速率(Pmax)、光飽和點()和暗呼吸速率(R)等光合生理參數[11, 12]; 同時采用弱光(PAR ≤200 μmol·m-2·s-1)條件下與P的線性回歸方法求得表觀量子效率()[1, 4]。采用直角雙曲線修正模型擬合所測定的光響應曲線, 得出最大凈光合速率(Pmax)、初始量子效率()、暗呼吸速率(R)、光補償點()和光飽和點()等參數指標, 并與實測值進行比較分析。
1.5 模擬分析
采用直角雙曲線修正模型[4, 10, 20]進行胡楊葉片光合作用-光響應曲線的擬合, 其表達式為:
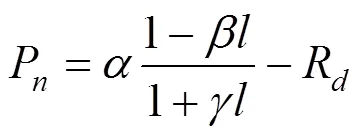
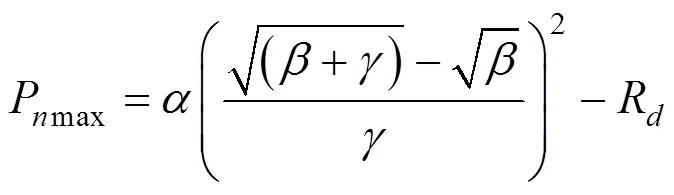

式中,P為凈光合速率,Pmax為最大凈光合速率,R為暗呼吸速率,為光合有效輻射, 本文中用表示,、、是獨立于的系數[4]。
植物在光補償點處的量子效率(φ), 內稟量子效率(φ)和光合作用光響應曲線上=0與=I兩點連線斜率的絕對值(φ)的數學表達式為[12]:




1.6 數據處理
采用Excel 2016軟件進行實驗數據處理, 使用SPSS 23.0軟件進行葉片形態性狀Pearson相關性分析。采用直角雙曲線修正模型擬合葉片的光合作用光響應曲線并計算光合特征參數[21], 使用OriginPro 2016軟件作圖。
2 結果與分析
2.1 葉片形態性狀
胡楊植株不同發育階段的葉片形態表現出顯著的差異性, 并呈現明顯的形態性狀特征和變化規律(表1)。
比較分析不同發育階段胡楊葉片的形態指標(表1)可知, 胡楊葉片的葉干物質含量、葉面積和葉干重均表現出隨胡楊樹齡增長和個體生長發育而逐漸增大的趨勢, 即A1< A2< A3< A4< A5< A6; 而比葉面積和葉片厚度則表現出逐漸減小的趨勢, 其中比葉面積的變化趨勢為A2> A1> A3> A5> A4> A6, 葉片厚度的變化趨勢為A2> A1> A3> A4> A5> A6。
為了準確檢驗葉片各形態性狀參數之間的相關性, 本文對胡楊不同葉片形態性狀之間的Pearson相關系數進行了統計分析。表2結果表明, 葉片干物質含量與葉片長/寬比值和比葉面積呈顯著負相關(< 0.05), 而與葉干重和葉面積呈極顯著正相關(< 0.01), 即隨著葉片面積、厚度、干重的增加, 葉片長/寬比值和比葉面積減小, 胡楊葉片干物質含量增大; 胡楊葉干重與比葉面積呈顯著負相關(< 0.05), 而與葉面積呈顯著正相關(< 0.05), 即比葉面積越小、葉面積越大, 其葉干重越大; 比葉面積和葉面積呈顯著負相關(< 0.05), 即葉面積越大比葉面積越小。
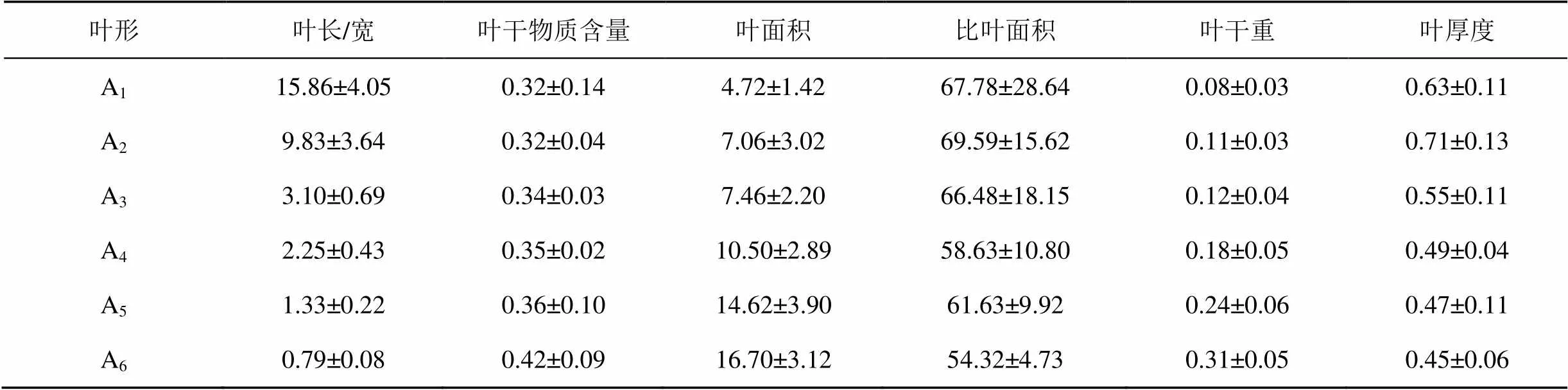
表1 胡楊不同葉片的形態性狀
注: A1幼苗條形葉; A2幼樹條形葉; A3幼樹披針形葉; A4成樹披針形葉; A5成樹卵圓形葉; A6成樹闊卵圓形葉。

表2 胡楊葉片形態性狀指標的相關分析
注:=6, **<0.01, *<0.05,為長/寬,為葉干物質含量,為葉面積,為比葉面積,為葉干重,為葉厚度。
2.2 葉片光合作用與光響應特征
胡楊不同形態葉片的P值隨個體生長發育出現明顯差異。由圖1可知, 6種形態葉片A1、A2、A3、A4、A5、A6在接近光飽和點時的P分別為20.84、21.28、19.18、14.95、17.31、22.55 μmol·m-2·s-1。其中幼苗期和幼樹期胡楊葉片的P相對較高, 成熟期胡楊闊卵圓形葉的P最高, 而生長在植株冠層下部的卵圓形葉和披針形葉的P明顯降低。
植物光合作用-光響應曲線反映了在一定光照強度下植物的潛在光合能力及植物對生長環境的適應性。實地觀測結果(圖1)表明, 胡楊不同形態葉片的光合速率(P)對光合有效輻射()表現出不同的響應規律。在低光強(< 200 μmol·m-2·s-1)條件下, 胡楊不同形態葉片的P表現出相似的變化趨勢, 近似呈線性增加。隨著光照強度的增大, 胡楊葉片P隨的變化呈曲線式上升; 至光飽和點()時,P曲線的變化漸趨平緩, 表現出一定的光飽和現象; 而不同形態葉片的P變化特征表現出較大的差異性。
胡楊幼苗期條形葉、幼樹期條形葉以及成熟期披針形葉的P在為1800 μmolm-2·s-1左右達到飽和, 并維持在較高水平, 即未發生明顯的光抑制。幼樹期披針形葉、成熟期卵圓形葉和闊卵圓形葉的P在達1800—2000 μmolm-2·s-1時仍能夠保持增長趨勢, 而且光抑制現象也不明顯。其中, 胡楊闊卵圓形葉P的上升趨勢最為明顯, 說明胡楊闊卵圓形葉具有適應極端干旱環境和強烈光照條件的能力, 對于胡楊適應干旱環境具有重要作用。
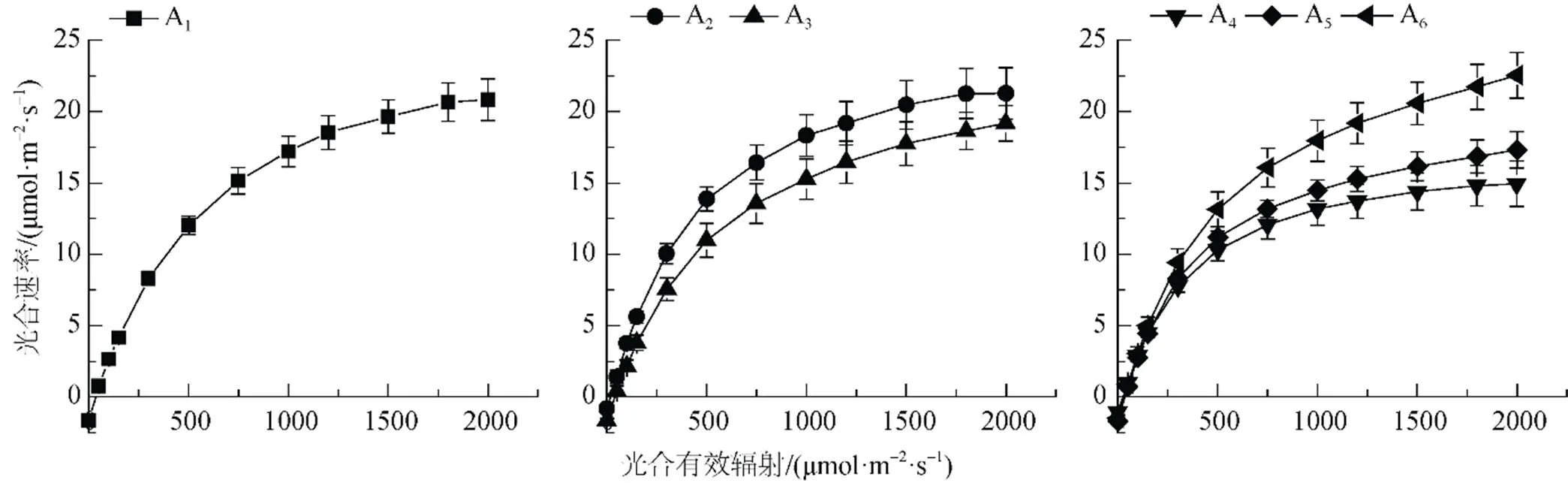
圖1 胡楊不同發育階段葉片光合作用的光響應曲線
Figure 1 The light response curve of leaves ofat different developmental stages.
成熟期胡楊卵圓形葉和披針形葉具有較低的P值, 這可能與其生長在植株冠層下部的空間分布有關。由于光照強度透過植物冠層時呈指數衰減變化, 因此, 位于空間分布的上部葉片(闊卵圓形葉)適應高光強條件, 而位于冠層下部的葉片(卵圓形葉和披針形葉)則適應相對較低的光照條件。這是胡楊成熟植株葉片光合作用從上至下依次降低的垂直空間變化的明顯特征。胡楊幼苗期和幼樹期的條形葉均具有與成熟期闊卵圓形葉幾乎相同的P值, 這與胡楊葉片的比葉面積具有較大的關系, 通常比葉面積較高的植物其葉子的光捕獲面積較高并由此導致較高的凈光合速率[22], 這跟表1顯示的結果相吻合。
2.3 葉片光合作用的光響應模擬
由于直角雙曲線模型、非直角雙曲線模型與指數模型均是一條沒有極值的漸近線, 且無法估算最大凈光合速率和飽和光強[1, 23, 24], 因此本文采用直角雙曲線修正模型對胡楊不同形態葉片光合作用的光響應曲線進行擬合, 得到如圖2所示的擬合結果。統計分析表明, 所有擬合方程系數(R2)均達到0.999以上(<0.01), 直角雙曲線修正模型適于胡楊不同形態葉片的光合作用-光響應曲線。
由表3可知, 擬合的胡楊葉片的最大凈光合速率(Pmax)與實測值較為接近, 其中不同形態葉片的Pmax變化依次為A6>A3>A1>A2>A5>A4, 表明成熟期胡楊闊卵圓形葉對干旱荒漠區高溫和高光強環境的適應能力最強。胡楊個體生長發育進入成熟期后, 其闊卵圓形葉位于樹冠上部, 長期生長在極端干旱的荒漠環境中, 逐漸形成了適應高溫、強光的光合特征, 有利于其干物質積累和適應荒漠逆境, 為較高物質產量的形成奠定良好的生理基礎; 而卵圓形葉和披針形葉由于長期處于遮陰狀態, 表現出較低的量子效率()、光飽和點()、光補償點()與呼吸速率(R), 導致其最大凈光合速率(Pmax)最小, 光合效率最低。此外, 擬合的胡楊闊卵圓形葉最大凈光合速率對應的光飽和點為2244.78 μmol·m-2·s-1, 而實測值只有2000 μmol·m-2·s-1, 說明胡楊闊卵圓形葉具有更高的光合潛力; 而卵圓形葉和披針形葉的擬合值均小于實測值, 說明這兩種形態葉片的光合速率在光強為2000 μmol·m2·s-1時已經達到了飽和狀態。
在低光強(≤200 μmol·m2·s-1)下, 葉片凈光合速率(P)對光強()的響應曲線是評價植物光合特性的重要因素[25], 常采用表觀量子效率來反映出植物對光能的利用效率[1, 24, 26]。但由于低光強條件下擬合植物葉片P與之間的關系時, 因擬合光合有效輻射的范圍不同, 則據此計算出來的表觀量子效率也不同[27]。因此, 表觀量子效率并非描述植物利用光能的理想指標, 而采用植物在光補償點處的量子效率()可以避免這個問題。因為植物的光補償點()只有一個, 植物在光補償點處的量子效率具有唯一性, 故可以用來表示植物葉片把光能轉化為凈能量的能力[1]。由表3可見, 胡楊不同形態葉片的量子效率大小排序為: A2> A6> A5> A4> A1> A3, 表明胡楊幼樹期條形葉的光能利用率最高, 這與其所處位置及其有關。植物反映了植物對弱光的利用能力[28], 胡楊幼樹期條形葉受上部葉片的遮光影響, 因而在一定程度上需要具有較強的弱光利用能力, 其實測的和R均較低, 這與表3中模型擬合的數據相一致。幼樹期條形葉具有較低的及相對高的,使其具有較大的光能利用區間,這是其能在干旱荒漠環境中保持較高光合速率的原因; 而披針形葉也具有較高的Pmax, 可能歸因于其具有較大的比葉面積(見表1), 同時由于該葉形分布于植株上部, 良好的光照使其具有較高的和R, 這是其適應較高光強的結果。
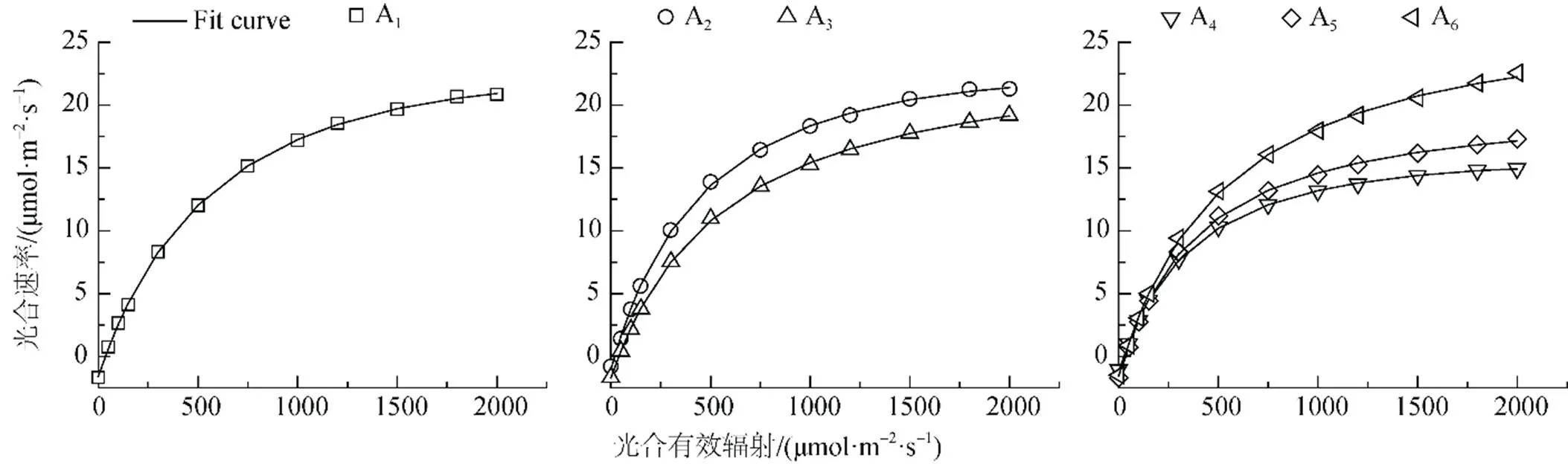
圖2 胡楊不同發育階段葉片光合作用-光響應的模擬
Figure 2 Simulation of photosynthetic-light response curves of leaves ofat different developmental stages.

表3 胡楊葉片光合作用-光響應特征參數實測值與模型擬合值比較
注: A1幼苗條形葉; A2幼樹條形葉; A3幼樹披針形葉; A4成樹披針形葉; A5成樹卵圓形葉; A6成樹闊卵圓形葉; 實測值φc=AQYPAR≤50。
胡楊幼苗期條形葉具有較高的、、R和Pmax, 表明其具有耐高光強的能力[29], 這種葉形的光合能力與其比葉面積具有密切關系。比葉面積是植物碳收獲策略的關鍵葉性狀之一, 具有較高比葉面積的葉片光捕獲面積較高, 具有較高的凈光合速率[22]。在胡楊的幼苗生長期, 條形葉通過維持較高的光合速率積累光合產物, 使其在干旱荒漠環境中具有較高的生長量。這可能是胡楊不同形態葉片著生在樹冠不同空間高度以及胡楊葉片從幼苗到成年葉形發生變化的原因, 這與王海珍等[27]的研究相一致。
3 討論
植物光合作用受諸多環境因素的影響, 如光照強度、溫度、CO2濃度和土壤水分等[30]。本研究中, 胡楊植株生長在同一生境中, 具有相同的空氣濕度, 實際觀測時的光強和CO2濃度保持一致, 因此本試驗的結果具有可比性。光響應參數能較好的反映逆境條件下植物的光合潛能、光能利用率等[31], 應用直角雙曲線修正模型對胡楊不同發育階段6種不同形態葉片光合作用-光響應曲線的擬合結果表明, 胡楊葉片的P隨的增強而增大; 當超過一定數值后,P將不再隨著的增強而增加, 出現光飽和現象。3種不同樹齡的胡楊各種形態葉片對強光環境的反應存在較大差異, 在較強的下, 成熟期闊卵圓形葉的P明顯高于卵圓形葉和披針形葉, 表明闊卵圓形葉具有較高的光和效率, 該結果與相關胡楊葉片光合特性研究結果相似。
在植物葉片光合作用-光響應研究中,、、Pmax和是反映植物光響應特征的重要指標[26],其中與反映了植物對光照條件的要求與適應性強弱[18],、Pmax高低則是植物光能利用效率和光合能力強弱的表征[32]。胡楊成熟期闊卵圓形葉具有明顯高于其他葉形的, 且和Pmax均較高, 表明胡楊闊卵圓形葉具有較高光合活性, 通過提高Pmax以增強光能利用率, 使其在干旱荒漠區高溫、強光環境下仍能保持較高的光能轉化效率與光合速率以及較寬的光能利用區間, 這為其干物質的積累、抗逆能力提高提供了良好的生理生態機制, 有益于胡楊在荒漠環境的生存和生長。
胡楊幼樹期條形葉具有較低的、R和較高的、Pmax, 表明其能夠迅速在弱光環境中達到較高的光合速率, 具有較寬的光能利用區間, 充足的物質積累有助于其適應惡劣的環境[33]; 這是其對不同光強做出的適應性反應和采取的生理適應機制, 這與胡楊從具有條形葉的幼苗成熟植物不同形態葉片發生變化具有密切的聯系。
通過胡楊幼苗期條形葉光合特性和形態性狀的分析可知, 其較高的光合速率歸因于葉片具有較大的比葉面積。條形葉具有較低的與相對較高的、及Pmax, 很大程度上也歸因于其較高的比葉面積, 高效的光捕獲面積使其具有較高的凈光合速率[34], 通過積累光合產物, 使胡楊在極端逆境下得以生存并能達到較高的生長量[14]。
通過采用直角雙曲線修正模型對胡楊葉片光合作用-光響應曲線進行擬合, 結果顯示, 其中胡楊葉片的Pmax、與實測結果基本上一致;明顯低于實測結果; 胡楊不同形態葉片光合作用-光響應曲線的擬合方程系數(2)均達到0.9995以上(<0.01), 說明直角雙曲線修正模型適于胡楊不同形態葉片光合生理參數的定量研究。
[1] 葉子飄. 光合作用對光和CO2響應模型的研究進展 [J]. 植物生態學報, 2010, 34(6): 727–40.
[2] ELFADL M A, LUUKKANEN O. Field studies on the ecological strategies ofin a dryland ecosystem: 1. A leaf gas exchange approach [J]. Journal of Arid Environments, 2006, 66(1): 1–15.
[3] SHARP R E, MATTHEWS M A, BOYER J S. Kok effect and the quantum yield of photosynthesis. Plant Physiology, 75, 95-101 [J]. 1984, 75(1): 95–101.
[4] 葉子飄, 于強. 光合作用光響應模型的比較 [J]. 植物生態學報, 2008, 32(6): 1356–61.
[5] SOCIETY T R. The Kinetics of Photosynthesis [J]. Procee-dings of the Royal Society of London, 1935, 149(868): 596–596.
[6] THORNLEY J H M. Mathematical models in plant physiology [J]. Mathematical Models in Plant Physiology, 1976,10(2):112–112
[7] BASSMAN J H, ZWIER J C. Gas exchange characteristics of,and[J]. Tree Physiology, 1991, 8(2): 145.
[8] PRADO C H, MORAES J A P V D. Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field conditions [J]. Photosyn-thetica, 1997, 33(1): 103–12.
[9] 葉子飄, 于強. 一個光合作用光響應新模型與傳統模型的比較 [J]. 沈陽農業大學學報, 2007, 38(6): 771–5.
[10] TUZET A, PERRIER A, LEUNING R. A coupled model of stomatal conductance, photosynthesis and transpiration [J]. Plant Cell & Environment, 2003, 26(7): 1097–116.
[11] XIA J B, ZHANG G C, WANG R R, et al. Effect of soil water availability on photosynthesis in Ziziphus jujuba var. spinosus in a sand habitat formed from seashells: Comparison of four models [J]. Photosynthetica, 2014, 52(2): 253–61.
[12] LANG Y, WANG M, ZHANG G C, et al. Experimental and simulated light responses of photosynthesis in leaves of three tree species under different soil water conditions [J]. Photosynthetica, 2013, 51(3): 370–8.
[13] 吳芹, 張光燦, 裴斌, 等. 不同土壤水分下山杏光合作用CO2響應過程及其模擬 [J]. 應用生態學報, 2013, 31(6): 4499–508.
[14] 蘇培璽, 張立新, 杜明武, 等. 胡楊不同葉形光合特性、水分利用效率及其對加富CO2的響應 [J]. 植物生態學報, 2003, 27(1): 34–40.
[15] 白雪, 張淑靜, 鄭彩霞, 等. 胡楊多態葉光合和水分生理的比較 [J]. 北京林業大學學報, 2011, 33(6): 47–52.
[16] MA H C, FUNG L, WANG S S, et al. Photosynthetic response of Populus euphratica to salt stress. Forest Ecol Manag [J]. Forest Ecology & Management, 1997, 93(1): 55–61.
[17] WANG H L, YANG S D, ZHANG C L. The photosynthetic characteristics of differently shaped leaves in Populus euphratica Olivier [J]. Photosynthetica, 1998, 34(4): 545–53.
[18] XIONG R, LI X, ZHANG X, et al. The studies about the photosynthetic response of the four desert plants [J]. Acta Ecologica Sinica, 2003, 23(3): 598–605.
[19] 王海珍, 韓路, 徐雅麗, 等. 胡楊異形葉葉綠素熒光特性對高溫的響應 [J]. 生態學報, 2011, 31(9): 2444– 53.
[20] 葉子飄. 光合作用對光響應新模型及其應用 [J]. 生物數學學報, 2008, 23(4): 710–6.
[21] LOBO F D A, BARROS M P D, DALMAGRO H J, et al. Fitting net photosynthetic light-response curves with Microsoft Excel — a critical look at the models [J]. Photosynthetica, 2014, 52(3): 479–80.
[22] ZHANG L, LUO T. Advances in ecological studies on leaf lifespan and associated leaf traits [J]. Acta Phytoecol Sinica, 2004, 28(6): 844–52.
[23] YE Z P, YU Q, KANG H J. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions [J]. Photosynthetica, 2012, 50(3): 472–6.
[24] 王榮榮, 夏江寶, 楊吉華, 等. 貝殼砂生境干旱脅迫下杠柳葉片光合光響應模型比較 [J]. 植物生態學報, 2013, 37(2): 111–21.
[25] OLSSON T, LEVERENZ J W. Non‐uniform stomatal closure and the apparent convexity of the photosynthetic photon flux density response curve [J]. Plant Cell & Environment, 1994, 17(6): 701–10.
[26] XIA J B. Critical responses of photosynthetic efficiency in Campsis radicans (L.) Seem to soil water and light inten-sities [J]. African Journal of Biotechnology, 2011, 10(77): 17748–54.
[27] 王海珍, 韓路, 徐雅麗, 等. 胡楊異形葉光合作用對光強與CO2濃度的響應 [J]. 植物生態學報, 2014, 38(10): 1099–109.
[28] SUN C X, QI H, HAO J J, et al. Single leaves photos-ynthetic characteristics of two insect-resistant transgenic cotton () varieties in response to light [J]. Photosynthetica, 2009, 47(3): 399–408.
[29] TARTACHNYK I I, BLANKE M M. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remo-bilization of apple leaves [J]. New Phytologist, 2004, 164 (3): 441–50.
[30] SOFO A, DICHIO B, MONTANARO G, et al. Photosy-
nthetic performance and light response of two olive cultivars under different water and light regimes [J]. Photo-synthetica, 2009, 47(4): 602–8.
[31] 張彥敏, 周廣勝. 植物葉片最大羧化速率及其對環境因子響應的研究進展[J]. 生態學報, 2012, 32(18): 5907–17.
[32] KRAUSE G H. Photoinhibition of photosynthesis. An eva-lua-tion of damaging and protective mechanisms [J]. Phy-siologia Plantarum, 1988, 74(3): 566–74.
[33] BRODRIBB T, HILL R S. Light Response Characteristics of a Morphologically Diverse Group of Southern Hemis-phere Conifers as Measured by Chlorophyll Fluo-rescence [J]. Oecologia, 1997, 110(1): 10–7.
[34] 姜衛兵, 莊猛, 韓浩章, 等. 彩葉植物呈色機理及光合特性研究進展 [J]. 園藝學報, 2005, 32(2): 352–8.
Leaf photosynthesis and light response characteristics ofin different developmental stages
SHAN Lingfei, DING Yinghui, WANG Shuanglei, ZHANG Moshi, HAN Hang, SHI Sha, FENG Jinchao*
College of Life and Environmental Sciences, Minzu University of China, Beijing 100081, China
is the tertiary relic plant with various forms of leaves at different stages of individual development. It is very important to study on the drought resistance and salt tolerance of, which is the major dominant species of oasis ecosystem in Ejina area, Inner Mongolia. It was analyzed for the leaf morphological characteristics, photosynthesis and light response process ofat different developmental stages. The results showed that the ratio of length to width, specific leaf area and leaf thickness ofdecreased from seedling stage, young tree stage to mature tree stage, while the leaf dry matter content, leaf area and leaf dry weight increased. The higher photosynthetic efficiency and photosynthetic activity of broad oval leaves ofat mature stage were maintained to increase photosynthetic rate under desert environment of high temperature and strong light. The higher photosynthetic rate of strip leaves at young tree stage was related to wider range of light energy utilization under weak light environment. The higher photosynthetic rate of strip leaves at young seedling stage was maintained due to higher specific leaf area and light capture, while light compensation point (LCP) and light saturation point (LSP) were also higher. The above results provided the base of adaptive responses and eco-physiological mechanisms ofunder different light environments. It was proved that the modified model of rectangular hyperbola was suitable for quantitative study of photosynthetic physiological parameters of different leaves of.
; leaf morphology; photosynthesis; light response; modified model of rectangular hyperbola.
10.14108/j.cnki.1008-8873.2019.06.004
Q14
A
1008-8873(2019)06-022-08
2018-10-30;
2019-03-03
國家自然科學基金項目(31570407); 中央民族大學協同創新中心項目(MUC2011-3)
單凌飛(1991—), 男, 河南鄭州人, 中央民族大學生命與環境科學學院碩士研究生, 研究方向為植物生態學, E-mail: shanlingfei@muc.edu.cn
馮金朝(1964—), 男, 博士, 教授, 主要從事植物生態生理學研究, E-mail:fengjinchao@muc.edu.cn
單凌飛, 丁穎慧, 王雙蕾, 等. 胡楊不同發育階段葉片光合作用及其光響應特征[J]. 生態科學, 2019, 38(6): 22-29.
SHAN Lingfei, DING Yinghui, WANG Shuanglei, et al. Leaf photosynthesis and light response characteristics ofin different developmental stages[J]. Ecological Science, 2019, 38(6): 22-29.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
光學精密工程(2016年6期)2016-11-07 09:07:19
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
核科學與工程(2015年4期)2015-09-26 11:59:03
