GJB2基因突變在長島型掌跖角化病發病中的作用
2020-01-10 06:43:24劉慧茹葉碧珍魏轉弟
安徽醫藥 2020年1期
關鍵詞:檢測
劉慧茹,葉碧珍,魏轉弟
作者單位:東莞市中醫院皮膚科,廣東 東莞523000
長島型掌跖角化癥(Nagashima-type palmoplantar Keratosis,NPPK)是一種常染色體隱性遺傳病,男女發病率相似,臨床表現的特征為掌跖角化癥,該病于1977年首次在長島被報告[1]。其臨床特征包括手掌和腳掌上界限清楚的紅斑并輕中度角化過度;長島型掌跖角化癥通常在嬰幼兒期開始發病,表現為輕微的掌跖斑疹病,皮疹進展慢[2]。
GJB2屬于β-族間隙連接蛋白家族,按蛋白分子量命名為Cx26。間隙連接蛋白與縫連接蛋白組成一個完整的縫隙連接通道,在細胞間信息傳導和物質交換起重要作用。表皮的基底層含有增殖的角質形成細胞,分化產生的細胞可遷移到棘突和顆粒層[3]。角質形成細胞具有許多細胞間連接,包括間隙連接[4]。β-族間隙連接蛋白突變可能誘導皮膚病發生,其中Cx26的GJB2突變會導致皮膚病綜合征,包括耳聾的掌跖角化病(PPK),Vohwinkel綜合征(VS)和角膜炎魚鱗病耳聾綜合征(KID)[5-7]。Cx31(GJB3)和Cx30.3(GJB4)突變引起的皮膚病癥狀相似,主要表現為變異性紅皮膚角化癥(EKV)[8]。目前大量研究集中在間隙連接蛋白導致KID綜合征的作用機制,但對于與NPPK相關的突變鮮有研究。
本研究于2017年收集了家系的2例NPPK病人,對病人的外周血提取DNA,進行SERPINB7基因外顯子8常見突變位點c.796C>T(p.Arg266Ter)進行測序,未發現突變。因此,我們推測該家系病人可能是由于其他相關基因(GJB2、GJB3、GJB4、GJB6、GJA1)突變引起。本研究擬通過鑒定NPPK的突變位點,闡明NPPK的發病機制,為NPPK的治療提供一定的理論依據。
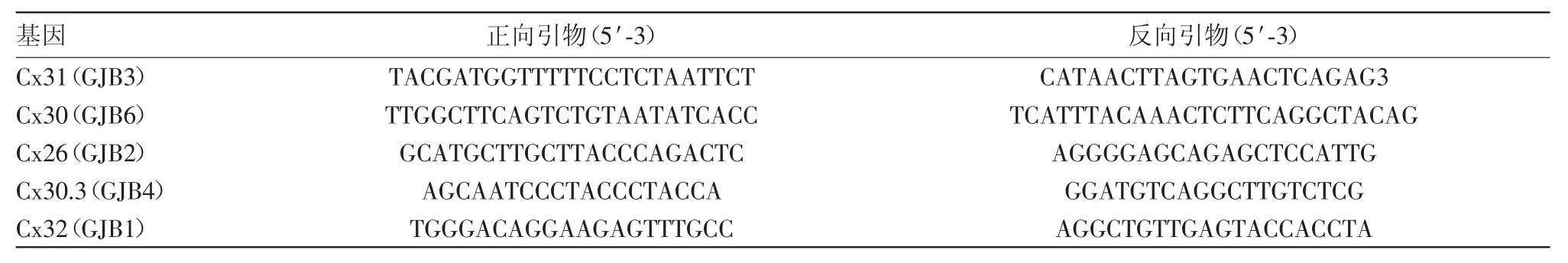
表1 長島型掌跖角化癥(NPPK)中GJB2、GJB3、GJB4、GJB6、GJA1基因PCR擴增引物序列
1 資料與方法
1.1 一般資料 研究對象為收集的2例家系NPPK病人,分別于3歲和4歲開始發病,臨床表現為雙手掌、指背、足底、手腕、踝部、跟腱等對稱性紅斑、角化性斑丘疹、斑塊伴脫屑。組織病理學表現為表皮角化過度、顆粒層和棘層增厚,無表皮松解性角化過度的特點,真皮淺層少量毛細血管擴張,少量炎癥細胞浸潤。該例病人的臨床及組織病理符合長島型掌跖角化癥。
1.2 實驗方法
1.2.1 DNA提取與sanger測序 收集NPPK病人外周血3~5 mL,根據DNA提取試劑盒說明書提取血液DNA,利用紫外分光光度計檢測DNA的濃度和純度,保存-70℃備用。從美國國家生物技術信息中心(https://www.ncbi.nlm.nih.gov/)中 獲 得 GJB2、GJB3、GJB4、GJB6、GJA1這5個基因的序列,設計引物(表1)。以下條件進行PCR擴增:95℃預變性3 min,95℃變性30 s,58℃退火30 s,在72℃延伸2 min,30個循環,72℃延伸10 min終止反應。1%瓊脂糖凝膠電泳檢測PCR產物,將獲得的5個基因的PCR產物送華大基因進行正反向引物測序。用Gene tool軟件將測定的序列與標準序列進行比對,標 準 序 列 參 考 :GJB2:NM_004004,GJB6:NM_001110219, GJB3: NM_001005752, GJB1: NM_000166,GJB4:NM_153212。
1.2.2 體外轉錄及卵母細胞顯微注射 將Cx26和Cx31克隆到pCS2+表達載體中,用于非洲爪蟾卵母細胞的功能研究[9]。通過定點誘變制備Cx26-S183F結構體[10],并將DNA結構體克隆到pBlueScri1ptⅡ載體中并測序,然后亞克隆到pCS2+表達載體中。將定點誘變的結構體使用SP6 mMessage mMachine體外轉錄試劑盒線性化并轉錄。從非洲爪蟾雌性中收集卵母細胞,并在改良的Barth(MB)培養基中培養[11],向卵母細胞注射10 ng反義非洲爪蟾Cx38寡核苷酸[12],然后單獨或組合注射連接蛋白轉錄物,注入水的卵母細胞作為陰性對照。Cx31 RNA和其他cRNA都以相同的濃度注射。
1.2.3 記錄半通道電流 卵母細胞注射各種cRNA 24h后使用Gene Clamp 500放大器檢測其半通道電流。將卵母細胞培養在不添加Ca2+的MB培養基中,在室溫條件下進行膜片鉗實驗。將電極拉至1~2 MΩ的電阻,電極內灌注電極內液。通過記錄半通道電流:鉗制電壓為-40 mV,電壓從-30 mV去極化至+40 mV,階躍10 mV每次持續5 s,從而獲得半通道電流-電壓(I-V)曲線。并在初始鉗制電壓-40 mV情況下,測量兩個細胞的電流變化來計算連接電導(Gj)。一個細胞受±20 mV的交變脈沖,在另一個細胞中記錄由電壓變化產生的電流,其大小與連接電流(Ij)相等,通過Ij除以電壓差計算電導率,即Gj=Ij/(V1-V2)。
1.2.4 蛋白質印跡法 卵母細胞如1.2.2做相同的處理,24 h后提取蛋白。在12%的SDS凝膠上電泳分離并轉移到硝酸纖維素膜上,用5%的脫脂奶粉37℃封閉 1 h,用 CX26(GBJ2)或 Cx31(GBJ3)(ABCAM,Cambridge)的多克隆抗體進行檢測,然后用辣根過氧化物酶標記的二抗37℃孵育1 h。用GADPH(ABCAM,Cambridge)作為內參。使用Image J軟件對條帶灰度值進行量化分析。
1.2.5 免疫共沉淀 卵母細胞如1.2.2做相同的處理,24 h后利用膜蛋白提取試劑盒提供的說明書提取膜蛋白。用PBS清洗兩遍珠子,用PBS將Protein A agarose配制成50%濃度,蛋白樣品中加入Protein A瓊脂糖珠去除非特異性蛋白。用Cx26抗體孵育,加入Protein A瓊脂糖珠來捕捉抗原抗體復合物,收集瓊脂糖珠-抗原抗體復合物,樣品煮沸5 min變性。在SDS凝膠上進行電泳,轉膜后使用Cx26或Cx31的抗體進行WB檢測蛋白質。
1.3 統計學方法 采用GraphPad Prism 6.0和SPSS 11.0軟件對實驗數據統計分析,進行單因素方差分析One-Way ANOVA,并用Student’s t檢驗進行差異比較分析,P<0.05為差異有統計學意義。
2 結果
2.1 sanger測序篩選出GJB2突變引起NPPK GJB2基因核苷酸序列第584位堿基由C突變成T(c.548C>T)發生了突變,導致編碼的蛋白第183位的絲氨酸被苯丙氨酸取代(p.Ser183Phe)。NPPK病人的父母均為GJB2p.Ser183Phe雜合子突變攜帶者,100個與該家族不相關的正常漢族人中沒有檢測到p.Ser183Phe突變位點。
2.2 Cx26突變體無功能且抑制野生型連接蛋白Cx31 已有研究證實Cx31突變體可引起嚴重的皮膚病,當其與野生型Cx31共表達時,通道電導率降低,影響間隙連接通道的能力[13]。因此我們猜測Cx26突變體可能通過與Cx31互相作用從而出現NPPK。為證實這一點,Cx26-S183F及其他表皮連接蛋白在非洲爪蟾卵母細胞中表達及Cx31在非洲爪蟾卵母細胞中表達,檢測間隙連接電導Gj。用水注射的卵母細胞陰性對照組的電導幾乎可忽略(Gj=0.18 μS),而只注射Cx26的細胞的平均Gj為8.6μS。在單獨注射Cx26-S183F的卵母細胞中測量的電導與陰性對照組細胞相同(Gj=0.13μS),說明功能喪失。當共同注射Cx26-S183F和Cx26時,平均電導為2.9 μS,顯著低于單獨注射Cx26的(P<0.05,圖1A)。Cx26-S183F還顯示出對Cx31的反式顯性抑制,當兩者都存在于卵母細胞中時,Cx26-S183F顯著抑制Cx31(Gj=0.77 μS,P<0.05,圖1B)。結果表明Cx26-S183F能有效抑制野生型Cx31,Cx26突變體與Cx31互作可能會產生NPPK。

圖1 檢測各處理組(注入水、Cx26、Cx26-S183F、Cx26-S183F+Cx26)卵母細胞的間隙連接的電導率:A為檢測野生型Cx26、Cx26突變體及Cx26-S183F+Cx26的電導率,B為檢測野生型Cx31、Cx31+Cx26及Cx31+Cx26-S183F的電導率
2.3 突變蛋白表達缺失不會引起電導率的降低用蛋白質印跡法檢測野生型和突變體連接蛋白的表達。結果顯示,注射Cx31、Cx31+Cx26、Cx31+Cx26-S183F的處理組中均檢測到Cx31蛋白(31kDa)(圖2A),灰度值分析表明各處理之間差異無統計學意義(P>0.05,圖2B)。同理,在Cx26、Cx31+Cx26、Cx26-S183F、Cx26+Cx26-S183F及Cx31+Cx26-S183F處理組中均檢測到Cx26(26kDa)(圖2C),且表達量一致,定量分析顯示各處理組之間差異無統計學意義(P>0.05,圖2D)。說明Cx26-S183F突變體存在時功能活性的喪失與Cx31的翻譯效率無關。
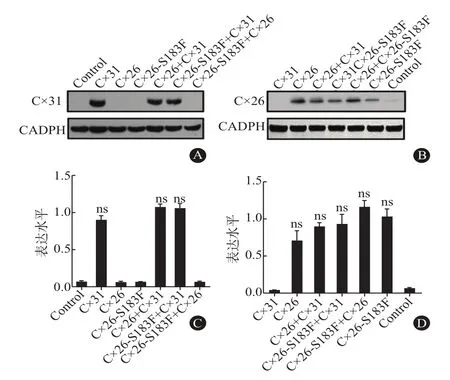
圖2 蛋白質印跡法檢測長島型掌跖角化癥(NPPK)突變體及野生型Cx31的表達水平:A為WB檢測各處理組中Cx31的表達情況;B為WB檢測各處理組中Cx26的表達情況;C為量化分析Cx31的蛋白表達量;D為量化分析Cx26的蛋白表達量
2.4 Cx26突變體與Cx31的互相作用 為了證實Cx26突變體和Cx31可能形成異型通道,我們對共表達Cx31和Cx26突變體的細胞進行免疫共沉淀實驗。結果表明,在Cx26相關樣品中檢測到Cx26蛋白,而在含有Cx31的樣品中均檢測到Cx31蛋白(圖3A和圖3B),說明細胞裂解液可進行后續CO-IP實驗。CO-IP顯示Cx26蛋白在野生型和突變體樣品中有表達,Cx26+Cx31樣品在31KDa出現較弱的條帶,說明野生型Cx26與Cx31結合較弱。共表達Cx31和Cx26-S183F樣品中蛋白的表達量較高,表明Cx26突變體蛋白與Cx31發生免疫共沉淀(圖3C)。
2.5 Cx26突變體不能形成功能性半通道 KID綜合征突變體單獨存在時通常形成活躍的半通道[14-16],而NPPK突變是否會出現類似的情況,為證實這一結果,在單個卵母細胞中表達Cx26和Cx26-S183F來分析半通道活性。結果顯示,注射水的卵母細胞對照在所有電壓階段被限流(圖4A),Cx26半通道活性在去極化時出現外向電流(圖4B),與注射Cx26的細胞相比,Cx26-S183F的膜電流顯著地降低(圖4C)。以平均膜電流和膜電壓作圖,表達CX26的細胞出現較高的外向電流,并隨著去極化的增加而增加,在+60mV時,Cx26產生的電流顯著高于對照及Cx26-S183F突變體(P<0.05,圖4D)。結果說明,注射突變體的細胞天然半通道活性喪失,Cx26-S183F與野生型Cx26的共表達導致半通道活性降低。
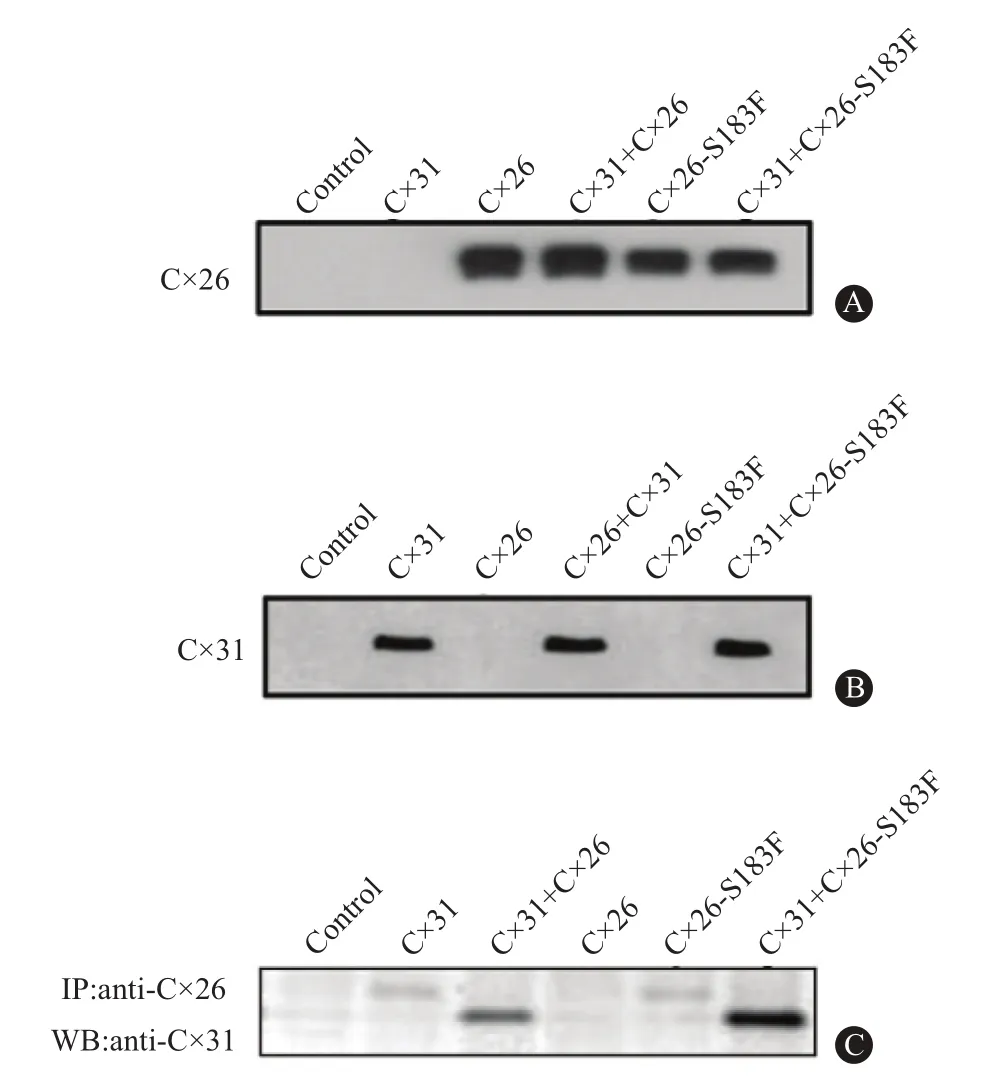
圖3 免疫共沉淀(CO-IP)檢測長島型掌跖角化癥(NPPK)中Cx26突變體與Cx31的互作情況:A和B分別為WB檢測細胞裂解液中Cx31、Cx26的表達情況;C為CO-IP檢測Cx26突變體與Cx的互相作用
圖4 膜片鉗實驗檢測長島型掌跖角化癥(NPPK)野生型Cx26和突變體的膜電流:A為檢測注射水的對照組卵母細胞的電流,B為檢測單獨注射Cx26的卵母細胞的電流,C為檢測單獨注射Cx26突變體的卵母細胞的電流,D為半通道I-V曲線圖
2.6 Cx26突變體能增強Cx31半通道 CO-IP實驗證明Cx31與異型半通道中的Cx26突變體結合,從而導致Cx26 NPPK突變體改變了Cx31的活性。因此驗證了Cx26-S183F是否也會影響Cx31半通道活性。結果顯示,單獨注射Cx31的卵母細胞沒有半通道活性(圖5A),由Cx31和Cx26-S183F組成的異型半通道出現較高的電流(圖5B)。以電流和電壓作圖,結果發現在電壓的增加下,共表達Cx26-S183F和Cx31的細胞中的半通道活性顯著增加,說明具有半通道功能。Cx31和Cx26-S183F共表達細胞產生的電流在-30、-20、-10、0、10、20、30、40、50、60 mV分別達到 0.135、0.21、0.235、0.3、0.315、0.38、0.40、0.41、0.44、0.46 μA,顯著高于單獨表達Cx31的細胞產生的電流(P=0.0316,圖5C)。這些結果說明,當共表達形成異型連接子時,Cx31和Cx26突變體能夠形成功能性半通道。
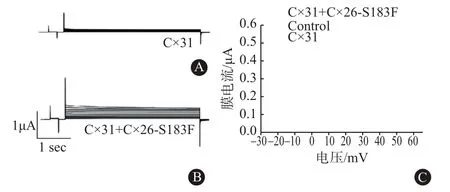
圖5 膜片鉗實驗檢測長島型掌跖角化癥(NPPK)Cx31和Cx26-S183F共表達時的膜電流:A為檢測單獨注射Cx31的卵母細胞的電流,B為檢測注射Cx26-S183F+Cx31的卵母細胞的電流,C為半通道I-V曲線圖
3 討論
NPPK由絲氨酸蛋白酶抑制劑B7(SERPINB7)基因突變引起。在本實驗中,測序發現2例病人的NPPK并不是由SERPINB7突變引起,通過測序發現一個與NPPK發生相關的GJB2突變。
當單獨表達時,Cx26-S183F不能形成功能性同源半通道或間隙連接,但與Cx31共表達后,降低了Cx31間隙連接的活性,并增加了半通道活性。Cx31和Cx26突變體的免疫共沉淀顯示形成異型連接子,Cx26突變體具有修飾Cx31半通道和縫隙連接的能力。
目前,大量研究表明間隙連接半通道在病理條件下能開放,為細胞間的物質交換提供通路。間隙連接半通道過量或長時間的開放會導致離子調節的失常,細胞內ATP濃度的變化和代謝產物丟失,最終導致細胞死亡[17]。不同間隙連接蛋白基因的突變可以導致同一種遺傳學疾病,已有研究發現,Cx26突變與Cx43互作會引起掌跖角化病[18]。本項研究發現了Cx26-S183F與Cx31共表達會增強間隙連接半通道活性,從而引起NPPK。
Cx31在表皮上表達,而且Cx31突變會引起遺傳性皮膚病,被稱為變異性紅皮膚角化病(EKV),其特征為軀干和四肢的皮膚過度角化,并伴有紅斑出現[19]。研究發現,與Cx26突變相關的引起的皮膚病有KID綜合征、PPK、先天性耳聾伴發角質厚皮病等[20-23]。目前為止關于Cx26突變引起皮膚病的機制有兩種:(1)抑制其他角質形成細胞連接蛋白;(2)異常的半通道的形成。反式顯性負效應作用會減少連接蛋白類型的數量從而影響表皮細胞間通訊,導致通道功能異常,而活躍的半通道會產生代謝物向細胞外釋放,可能對鄰近細胞產生不利影響,從而引發細胞凋亡[24]。
在Cx26和Cx31突變引起皮膚病的病人中,觀察兩者之間的臨床特征存在相似性。四肢出現過度角化病變,這表明可能與NPPK出現的機制相似。除此之外,還有Cx26突變引起的Bart-Pumphrey綜合征KHLS也有相似的臨床癥狀,包括角化過度和白細胞減少癥[25]。盡管癥狀相似,但仍需要進一步研究突變體確認其相關機制,Cx31-G8V突變的表達導致半通道的形成,允許鈣離子流入細胞,從而造成KHLS[26]。有研究發現Cx30.3突變體會引起變異性紅皮膚角化病(EKV),當Cx31和Cx30.3-F137L共同轉染時,在細胞膜上檢測到與Cx31形成間隙連接,并能形成異聚體連接子[27]。最近的一項研究表明,造成KID形成的Cx26-S17F突變體存在的情況下,Cx31半通道活性增加[28]。這與我們的研究結果一致,Cx26-S183F不能單獨形成半通道或間隙連接,但在與Cx31共表達時可以增強半通道活性,從而引起NPPK。
這些研究結果表明Cx26在皮膚疾病中的重要性,并進一步突出了Cx26突變體和Cx31互相作用在遺傳性皮膚病中的作用機制,為進一步治療皮膚病提供一定的理論基礎。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
