煙草開花期全基因組關聯分析
2020-01-18 13:35:41孫瀅姜自鵬劉洪泰孫明銘蔣彩虹任民劉旦羅朝鵬張劍鋒楊軍楊愛國程立銳
中國煙草科學 2020年6期
關鍵詞:煙草
孫瀅 姜自鵬 劉洪泰 孫明銘 蔣彩虹 任民 劉旦 羅朝鵬 張劍鋒 楊軍 楊愛國 程立銳


摘要:煙草開花期影響煙葉的產量和品質,是決定生態適應性和產量的重要性狀,與各種生物或非生物脅迫密切相關。研究煙草開花期的遺傳規律,進而解析調控開花時間的分子基礎,對煙草品種改良具有重要意義。本研究選用煙草MAGIC(Multiparent Advanced Generation Intercrossing)群體為材料,利用高密度煙草SNP(Single Nucleotide Polymorphism)芯片進行基因型分析,進而對開花期性狀進行全基因組關聯分析(Genome-Wide Association Study,GWAS)。結果表明,共檢測到13個與煙草開花期性狀顯著關聯位點,其中位于12號染色體109616410bp位置上的SNP為最顯著關聯位點。根據參考基因組,初步確定了控制開花期的關鍵候選基因Mab02790610,該基因與擬南芥控制開花的關鍵基因SOC高度同源,可能在控制煙草開花時間上起到關鍵作用。研究結果為進一步揭示煙草開花期遺傳調控機制及育種改良奠定了基礎。
關鍵詞:單核苷酸多態性(SNP):全基因組關聯分析(GWAS);煙草;開花期
煙草(Nicotiana tabacum L.)是我國重要的經濟作物,在國民經濟中具有十分重要的地位。作為茄科作物之一,其開花時間提前或延遲,不僅影響產品器官的產量和品質,還與各種生物或非生物脅迫密切相關。
開花期作為重要的煙草農藝性狀,屬于數量性狀,遺傳復雜,易受光周期、溫度等環境影響。普通煙草的一個野生親本林煙草為長日照植物,而另外一個野生親本絨毛狀煙草表現為短日照植物。其后代普通煙草大多數品種都為中性或者偏短日照。經典的遺傳學研究表明,普通煙草的開花期受一對隱性基因控制。目前,關于煙草開花期遺傳機制的報道,主要集中在控制開花的關鍵基因以及基因與環境互作上。FT、SOC(suppressor of overexpression of constans 1)、AGL24(gamouslike24)和LFY(e/)等都是成花過程中重要的調控基因,其中,FT是決定是否成花的最關鍵基因,最早在擬南芥晚花突體中發現。HARING等在煙草中鑒定到4個Fr同源基因FT、MF2、NF73和NFT4,并證明其只受短日照條件影響。SMYKAL等從煙草中克隆到SOC,過表達煙草會提前開花。李元元等克隆獲得了MFLC全長序列,并分析了NC82品種移栽后不同時間進行低溫處理該基因的表達特征,證明該基因與低溫敏感、早花特性有關;白戈等從煙草中克隆了一個高度保守的MYB類啟動子基因(NTMYB15),過表達該基因的煙草K326經低溫脅迫出現晚花;楊永銀等發現,低溫會使抗壞血酸氧化酶(AO)表達水平增加,脫氫抗壞血酸(DHA)含量增加,而且DHA能誘導SOC1表達水平升高,使開花提前。
隨著分子生物學技術的不斷發展,在煙草上針對農藝、抗性和品質等重要性狀,利用關聯分析或連鎖分析方法進行數量性狀位點(QL)發掘和標記開發研究已有多篇報道131,但是有關煙草開花期的遺傳定位研究報道很少。蔡長春等利用早花品種K326和遲花品種興煙1號組合的D群體和遺傳連鎖圖譜對煙草開花期進行了初步的遺傳分析,檢測到2個主效QL位點,表型變異貢獻率分別為24.5%和17.3%。而針對煙草開花期的全基因組關聯分析未見報道。
本研究利用8個不同類型煙草種質為親本組配的MAGIC群體為材料,對煙草開花期性狀進行全基因組關聯分析,發掘與花期關聯的SNP位點,預測候選基因,不僅為揭示煙草控制花期遺傳機制提高煙草適應性提供理論基礎,也為煙草廣適性育種奠定基礎。
1材料與方法
1供試材料與試驗設計
1.1.1試驗材料供試群體由本研究組配置
Beinhart1000-1、Florida301、Samsun、Basma、紅花大金元、VAM、塘蓬和小花青等8個不同煙草類型種質的雜交組合,進而構建了基于有限親本的高代互交系MAGIC群體(Multiparent Advanced Generation Inter-cross),群體材料有519個株系。
1.1.2試驗地點2019年3月15日,在中國農業科學院煙草硏究所諸城試驗基地種植8個親本及MAGIC群體材料,行距60cm,株距為40cm,每材料種植1行10棵,2次重復。
1.2表型調查
以2019年3月15播種日期為T1,某株系內50%植株中心花開放的日期為T2,T2與T1之間間隔的天數即為該株系的開花期。
1.3MAGIC群體基因型鑒定
2019年8月18日針對8個親本和MAGIC群體600個株系,選取株系內長勢一致單株的鮮嫩葉片混合取樣,利用DNA提取試劑盒提取DNA。應用高密度煙草SNP芯片(Gene Tian芯片,美國Affymetrix公司)對不同樣品進行全基因組掃描。參照張劍鋒等的方法略有調整,進行SNP芯片檢測。并對Poly high resolution類型的位點數據進行相應處理和分析,共獲得在親本間具有多態性的高質量SNP位點124153個,用于群體基因型分析。
1.4全基因組關聯分析
應用TASSEL5.0軟件中的混合線性模型MLM(Mixed Linear Model)進行性狀與標記之間的全基因組關聯分析,以軟件給出的p值作為顯著關聯的閾值。當標記的p≤0.0001時,則認為標記與性狀之間存在關聯。
2結果
2.1表型變異及SNP標記多態性
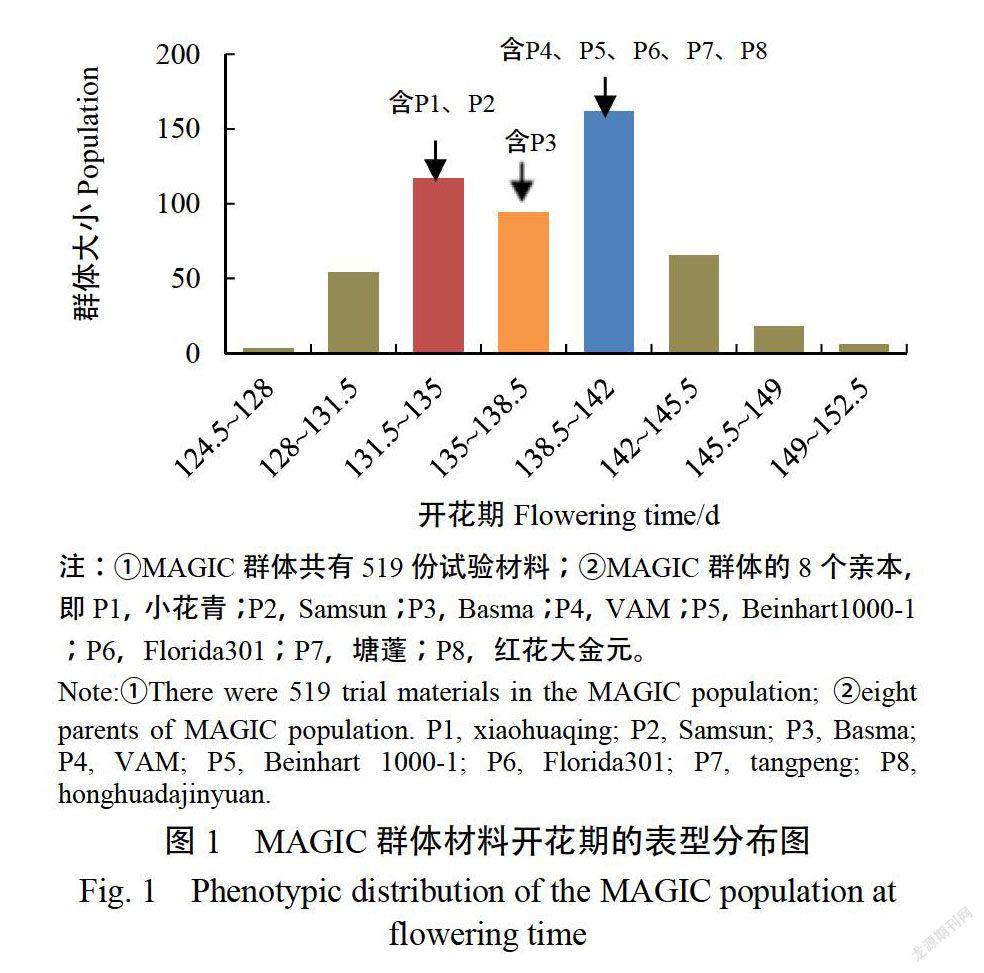
如圖1所示,煙草開花期在親本及群體中存在顯著差異。親本小花青和Samsun開花期較早,分別是132和133d;剩余6個親本Basma、VAM、紅花大金元、塘蓬、Beinhart1000-1、Florida301的開花時間很相近,分別在138、139、140、140、140.5、140.5d。供試群體開花期的最小值為124.5d,而最大值有152d,開花期的峰度和偏度的絕對值均小于1,說明開花期具有正態分布特征,呈現數量性狀遺傳特征。
2.2花期與SNP的全基因組關聯分析(GWAS)
采用TASSELV5.0軟件MLM模型,對519份MAGIC群體材料進行開花期的全基因組關聯分析,如圖2、3所示。其中共檢測到13個與開花期顯著關聯的位點p《000)(表1),分別位于煙草第4、5、6、8、12、13、15、17、19號染色體上,根據值排序,在煙草全基因組染色體上的物理位置分別為:12號染色體,109616410bp;8號染色體,2266918bp8號染色體,135192920bp;17號染色體,43342018bp6號染色體,74138868p:4號染色體,35423572bp:4號染色體,91394779bp2號染色體,57703873bp:13號染色體,76563713bp:15號染色體,21104329bp;5號染色體,138720068bp;6號染色體,146052300bp:19號染色體,40502891bp其中,4、6、8號染色體分別檢測出2個顯著性位點,12號染色體上109616410bp處的SNP位點顯著性最高,p值為4.79;單個位點表型変異貢獻率(R)范圍為3.38~4.74%,其中最顯著位點的表型貢獻率為4.49%。
2.3預測候選基因
根據關聯分析結果,位于12號染色體上的SNP位點AX-117626364最可能與開花期性狀緊密關聯。
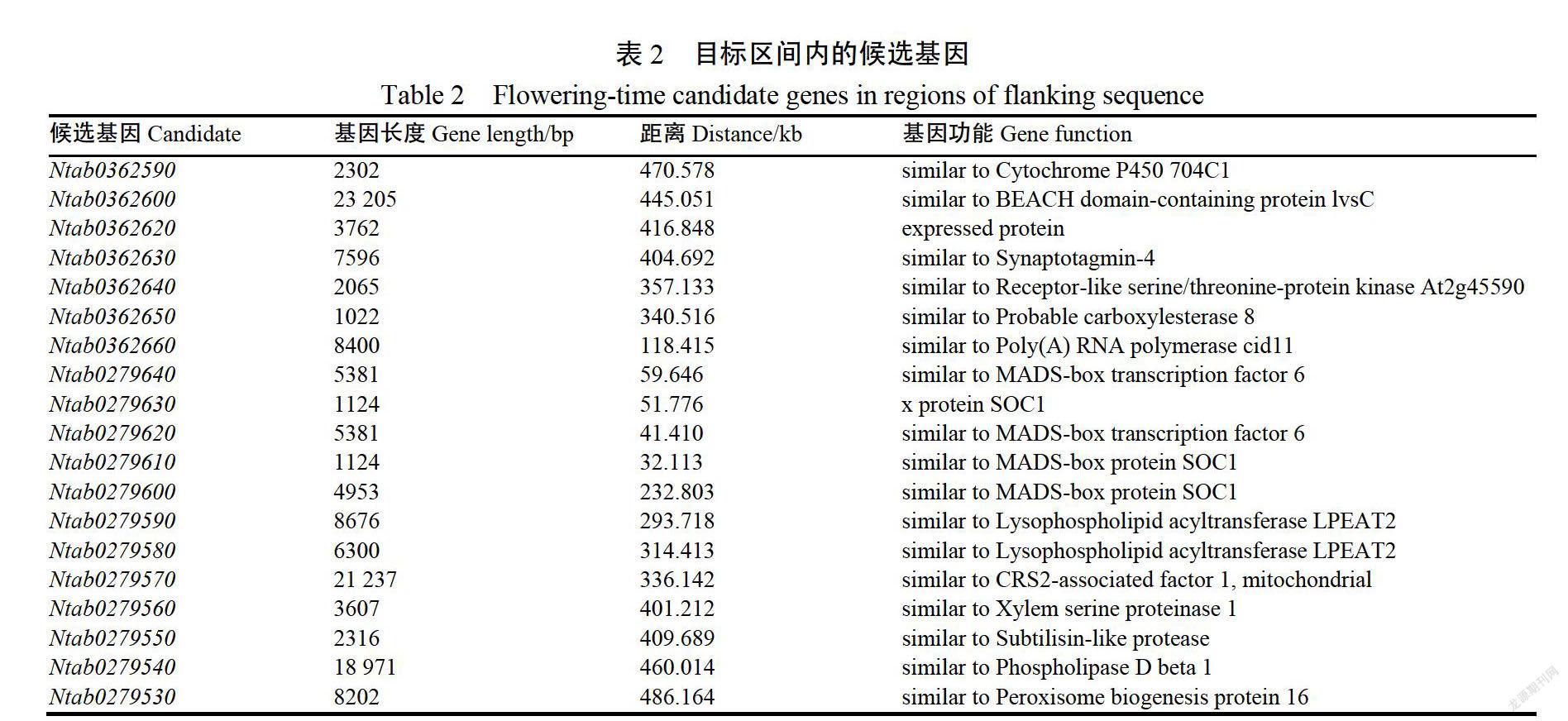
因此,以QFT1上下500kb區間范圍內進行候選基因篩選。如表2所示,目標區間內總共有19個編碼基因,注釋功能為細胞色素P450、突觸結合、絲氨酸/蘇氨酸蛋白激酶活性、羧酸酯酶活性MADS-box蛋白、溶血磷脂酰基轉移酶、木質部絲氨酸蛋白酶、類草菌素蛋白酶、磷脂酶β1、過氧化物酶體生物發生蛋白等。其中Mab0279640和Mab0279620編碼與擬南芥MADS-box轉錄因子功能相似蛋白,控制著花器官形成、發育等重要生理過程1Mab0279630、Mab02790610和Mab0279600與擬南芥(Arabidopsis thaliana)中的花卉整合子SOC1功能相似,調控上游CONSTANS基因(CO)影響開花時間2。其中Mab0279610距離最顯著的SNP位點的物理距離僅為32.11kb,因此初步將其確定為控制烤煙開花期關鍵候選基因。
3討論
3.1Magic群體
國際上早在2002年就已經提出包含有8個親本的互交設計,又稱為多親本高世代互交(Multiparent Advanced Generation Inter-crossing,MAGIC)。2008年,CAVANAGEI等2向植物界引進了多親本高代互交(Muli-parent advanced generation Inter-cross)群體,簡稱為MAGIC群體。
相比雙親群體和自然群體,MAGIC群體具有顯著優勢。本研究中以8個優質特色品種構建的煙草上第一套MAGIC群體,其中包含烤煙(紅花大金元)、晾曬煙(小花青、塘蓬)、香料煙(Samsun、Basma)、雪茄煙(Beinhart1000-1、Florida301)和白肋煙(VAM)。與自然群體相比,MAGC群體沒有群體結構問題,避免了關聯分析結果出現假陽性。全基因組分析結果共有13個與花期顯著相關的位點,群體遺傳豐富性得到很好的體現參考煙草全基因功能注釋,由最顯著位點初步篩選到Mab0279610,該基因與擬南芥中控制開花關鍵基因SOC同源,也極大可能是控制烤煙開花時間的關鍵基因。由此看出,MAGIC群體的連續雜交,極大地提高了重組概率,加了定位的準確性。
結果表明,MAGIC群體相比雙親群體和自然群體,不僅在數量性狀的關聯分析及精細定位方面等具有極大的優勢,而且群體規模越大,定位越準確。總而言之,由8個不同種質構建的MAGIC群體既可以彌補雙親群體重組發生有限、作圖分辨率低的不足,又能控制群體結構,非常適合當前的遺傳和育種研究,是理想的遺傳和育種研究材料。
3.2SNP標記
單核苷酸多態性(Single Nucleotide Polymorphism,SNP)標記作為目前最具發展潛力的分子標記,包含有單堿基的轉換、顛倒、插入及缺失等多種形式,在基因組中具有遺傳穩定、數量多、分布廣、易于檢測等特點,適合于MAGIC群體的檢測分析,滿足全基因組關聯分析對于大樣本高密度標記的要求,可以大大提高關聯分析的統計效率。
基于SNP標記的全基因組關聯分析所定位的QTL,在MAS中可提高選擇的目的性和準確性,進而提高育種效率,已成功應用于人類2、果蠅等2多種生物的遺傳研究。但是在煙草中目前報道的關聯分析研究大多采用SSR標記27-,分析群體的分子標記數目一般小于300個,如此少的標記數目很難實現全基因組分析。本研究利用高密度SNP芯片對MAGIC群體進行分析,總共獲得了124153個多態性位點的基因型數據,多態性標記覆蓋了煙草整個基因組,為開展煙草重要性狀全基因組關聯分析定了堅實的數據基礎。
3.3SOCI
利用MAGIC群體和高密度SNP分子標記,我門對控制開花的關鍵位點進行了全基因組關聯分析。研究結果表明,距離最顯著的SNP位點32.113kb位置上有個與擬南芥SOC基因同源的基因。植 物經過長期的發育進化,形成了一套復雜而精細的基因調控網絡,以確保植株能在最佳時間開花。MADS-box蛋白是一種轉錄調控因子,在控制植物開花時間、花器官形成等方面的信號轉導過程中起著關鍵作用SOC1作為一個集成多個開花信號的整合子,已經被深入研究了十幾年。SOC1作為一個MADS轉錄調控因子,在控制植物開花時間、花器官形成和分生組織特異性等方面的信號轉導過程中起著關鍵作用,是構成花卉時序和花卉發育基礎調控網絡的主要樞紐。因此,根據我們的定位結果及候選基因分析,將Nab0279610初步確定為候選基因,相關功能驗證正在開展中。
4結論
利用8個不同類型煙草種質為親本構建了煙草MAGIC群體,結合124153個SNP標記,進行基因型分型結果和全基因組關聯分析,共挖掘到10個與煙草花期相關聯的位點(p《0.0001)。根據煙草全基因組數據庫,初步將基因ab0279610確定為控制烤姻開花時間的關鍵候選基因,為進一步開展煙草開花期分子機制研究及分子育種奠定了基礎。
參考文獻
[1]張素芝,左建儒.擬南芥開花時間調控的研究進展[J].生物化學與生物物理進展,2006(4):301-309
[2]解敏敏,龔達平,孫榕,等.煙草成花素FT基因及其調控機制研究進展中國煙草科學,2018,39(3):98-102
[3]佟道儒.煙草育種學[M]北京:中國農業出版社,1997
[4] YOO S K, CHUNG K S, KIM J, et al. CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS I through FLOWERING LOCUS T to promote flowering inarabidopsisj Plant Physiology, 2005, 139(2): 770-778
[5] LEE J, OH M, PARK H, et al. SOCI translocated to the nucleus by interaction with AGL24 directly regulates leafy. Plant Journal2008,55(5):832-843
[5] TURCK F, FORNARA F, COUPLAND G, et al. Regulation and identity of florigen: flowering locus T moves center stage [J]. Annual eview of Plant Biology, 2008, 59: 573-594
[7] KOORNNEEF M, HANHART C J, VEEN JH, et al. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana [J]. Molecular and General Genetics MGG, 1991, 229(1)
[8] HARIG L, BEINECKE F A, OLTMANNS J, et al. Proteins from the FLOWERING LOCUS T-like subclade of the PEBP family act antagonistically to regulate floral initiation in tobacco [J]. Plant ournal,2012,72(6):908-921
[9] SMYKAL P, GENNEN J, D et al. Flowering of hotoperiodic Nicotiana varieties in non-inductive conditions by transgenic approaches[J]. Plant Molecular Biology, 2007, 65(3)23-242
[10]李元元低溫誘導煙草早花研究與煙草MADS-box基因的同源克隆[D]北京:中國農業科學院,2011
[11]白戈,楊大海,姚恒,等.煙草耐低溫脅迫早花MB5的克隆及功能分析[J]分子植物育種,2018,16(1):3504-3508
[12]]楊永銀,胞外脫氫抗壞血酸在低溫誘導煙草早花過程中的參與機制[D]鄭州:河南農業大學,2015
[13] DRAKE K, BAKAHER N, GOEPFERT S, et al. Multiple disease resistance loci affect soilborne disease resistance in tobacco (icotiana tabacum L.). Phytopathology, 2017, 107(9)1055-1061
[14]焦天雷.煙草(Nicotiana tabacum L.)赤星病抗性QL的定位分析[D]杭州:浙江大學,201
[15]牟建英,錢玉梅,任民,等煙草白粉病抗性基因的QL定位中國煙草學報,2013,19(4):105-108
[16]李茜.煙草遺傳圖譜構建與農藝性狀和化學成分的QTL定位[D]北碚:西南大學,2015
[17]蔡長春,鄧環,馮吉,等.烤煙生育期相關性狀的初步遺傳分析[J].中國煙草學報,2014,20(6):70-7
[18]張劍鋒,羅朝鵬,何聲寶,等.應用SNP標記分析24份煙草品種的遺傳多樣性[J].煙草科技,201750(11):1-8
[19]李元元,楊愛國,王魯,等.煙草MADS-box家族基因保守片段的克隆與序列分析[J].中國煙草科學,201,32(3):21-27
[20]陸瑩,劉艷華,任民,等.煙草CONSTANS同源基因的克隆與分析[J].中國煙草科學,2013,34(3):60-64.
[21] CAVANAGH C, MORELL M, MACKAY I, et al. From mutations to MAGIC: resources for gene discovery, validation and delivery in crop plants [J]. Current Opinion in Plant Biology, 2008, 11(2)215-221
[22] MARTIN W G, THOMAS A, MARION S, et al. SNP identification in crop plants [J]. Current Opinion in Plant Biology, 2009, 12(2)211-217
[23]陳秋玲,高建明,羅峰,等分子標記技術在禾本科作物基因定位上的研究進展[J]中國農學通報,2010,26(9):42-48CHENQL,GAOJM,
[24] PALAISA K A, MORGANTE M, WILLIAMS M, et al. Contrasting effects of selection on sequence diversity and linkage disequilibrium at two phytoene synthaseloc[J] The Plant Cell, 2003, 15(8)1795-1806.
[25] MICK E, MCGOUGH JJ, MIDDLETON F A, et al. Genome-wide association study of blood pressure response to methylphenidate treatment of attention-deficit/hyperactivity disorder[J]. Progress in Neuro-psychopharmacology and Biological Psychiatry, 2011, 35(2)466-472
[26] GERRARD D T, FRICKE C, EDWARD D A, et al. Genome-wide responses of female fruit flies subjected to divergent mating regimes. PLOS ONE, 2013, 8(6): e68136
[27]任民,張長靜,蔣彩虹,等.基于高密度SSR連鎖群的煙草致香物質關聯分析[J]中國煙草學報,2014,20(4):88-93
[28]樊文強,孫鑫,楊愛國,等.基于關聯分析發掘煙草高鉀優異等位變異[J]中國煙草學報,2016,22(2):100-107
[29]童治軍,陳學軍,方敦煌,等.231份烤煙種質資源SSR標記遺傳多樣性及其與農藝性狀和化學成分的關聯分析[J]中國煙草學報,2017,23(5):31-61
[30]朱承廣,任民,蔣彩虹,等以關聯分析發掘煙草抗赤星病基因分子標記[J]中國煙草科學,2017,38(1):68-72
[31]姚琦園,李紛芬,張林成,等.植物MADS-bx轉錄因子參與調控非生物脅迫的研究進展[J].江西農業學報,2018,30(5):73-79YAOQY,LI
[32]陳嬌.芥菜AGL24基因的啟動子克隆與其互作因子SOC1遺傳轉化研究[D].北碚:西南大學,2016
[33] SIMPSON GG, DEAN C. Arabidopsis, the rosetta stone of flowering time?[J]. Science, 2002, 296(5566): 285-289
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
