刺梨GL2同源基因的克隆、系譜樹和表達分析
2020-03-02 07:46:59黃小龍陳婷婷張琴琴莫佳佳龔盼琴閆慧清
廣西植物 2020年1期
黃小龍 陳婷婷 張琴琴 莫佳佳 龔盼琴 閆慧清

摘 要:為了觀察刺梨果實的果刺細胞學發育過程,該研究以刺梨‘貴農5號的cDNA為模板,通過RACE克隆獲得刺梨中與擬南芥表皮毛形成GL2的同源基因RrGL2,并對該基因進行生物信息學分析和表達分析。結果表明:(1)刺結構在花芽形成早期基部內的細胞首先不斷分裂,向外繼續發育,中部的細胞變細、變長形成“針”狀結構,頂部的細胞逐漸木質化使刺變硬,形成果刺。(2)通過RACE擴增得到RrGL2的cDNA全長2 292 bp,編碼763 aa氨基酸。(3)RrGL2具有Homeodomain同源結構域和StAR磷脂酰膽堿轉移蛋白的結構域,RrGL2與其他物種編碼的GL2氨基酸同源性高度相似,并且系譜樹分析揭示刺梨RrGL2和野草莓的GL2密切相關。(4)qRT-PCR分析表明,RrGL2在莖和果實中的表達水平高于其他組織,在花后7周果刺中的表達最高,是3周和5周果刺中的7.87倍和2.10倍。綜上結果發現RrGL2的功能與果刺的形成發育密切相關,該研究為刺梨中刺形成的分子機制和育種提供了理論基礎。
關鍵詞:刺梨,表皮毛,果刺,RrGL2,基因表達
中圖分類號:Q949.45文獻標識碼:A
Abstract:In order to observe cytological development of prickles in fruits of Rosa roxburghii, R. roxburghii GLABROUS 2 (RrGL2),a prickle-development related AtGL2 homology gene,was isolated from ‘Guinong 5 and relative biological information and expression were analyzed in this paper. The cytological development of fruit thorn of Rosa roxburghii was observed by paraffin section. Leaves of Rosa roxburghii was used to synthesize cDNA based on the manufacturers instructions of RACE. Subsequently RrGL2 was made relative informatics analysis and the gene expression level was eva-luated. The results were as follows:(1) The base cells continuously divided at the early stage of flower bud,then outward developed. The middle cells continued to become thinner and longer to form a “needle” structure. In the early stage of flower bud formation,the cells in the base of the thorn structure first divided continuously and continued to develop outwards. The cells in the middle became thinner and longer,forming a “needle” structure. The lignification gradually was observed at the top cells to make the prickles hard. (2) The full lengths of RrGL2 was 2 292 bp by RACE,encoding 763 amino acids. (3) The RrGL2 had a structure of Homeodomain and StAR phosphatidylcholine transfer protein,which is likely to regulate the development of Rosa roxburghii prickles. Then,a search for homologous species in the NCBI databases revealed a high similarity of amino acid homology encoded by the RrGL2 with other Rosa species,and phylogenic analysis revealed a close relationship of structure domains between Rosa roxburghii and Fragaria vesca. (4) Finally,real-time PCR analysis showed that the relative expression value of RrGL2 in fruit prickles after seven weeks after flowering was the highest,almost respectively 7.87 times and 2.10 times than that during three weeks and during five weeks after flowering. RrGL2,a prickles-forming gene acted to regulate the morphology and development of prickles. Therefore,the function of RrGL2 is closely related to thorn formation. These results could provide theoretical basis for thorn formation and development.
Key words:Rosa roxburghii,trichomes,prickles,RrGL2,gene expression
刺梨(Rosa roxburghii)為薔薇科多年生落葉灌木繅絲花的果實,因具有良好的風味和較高的營養價值深受消費者喜愛,在中國西南地區尤其是貴州省大規模種植。果實中因含有一些酚類化合物、抗氧化物等(Van et al.,2008),可作為放射性保護劑和腫瘤抑制劑(Liu et al.,2012;Xu et al.,2014);同時也是果汁和干果的重要原料。然而,刺梨果實密披果刺,果刺對水果采摘、食品加工、田間和果園管理造成不便。近年來隨著園藝業的發展,植物果刺已經引起越來越多的關注。
刺是廣泛存在于許多植物中由表皮組織生長所形成的漸尖突起(Kellogg et al.,2011),可存在于葉、莖、果實和其他器官(Feng et al.,2015)。刺的存在增加植物表皮厚度,減少熱量和水分的散失,防止昆蟲和病原體的侵襲或機械損傷(Gomes et al.,2012)。覆盆子和玫瑰的刺來源于修飾的腺毛,它們繼續生長并最終硬化成最終的刺狀形態,作為表皮組織的外生物。因此,刺是表皮毛與少數皮層細胞的變形所形成的;表皮毛是植物器官表面一種特殊的單細胞結構;表皮毛發育所需基因的時空表達受一種三元激活復合物協調(An et al.,2011)。植物表皮毛發育的調節特別是在擬南芥中的研究取得了很大的進展。通過轉錄因子MYB[GLABRA 1(GL1)、WEREWOLF、CAPRICE、TRIPTYCHON];WD-40型[(TRANSPARENT TESTA GLABRA 1(TTG1];bHLH[GLABRA 3(GL3)];ENHANCER of GLABRA 3(EGL3);HD-zip[GLABRA2(GL2)]和WRKYl類轉錄因子[TRANSPARENT TESTA GLABRA2(TTG2)](Zhao et al.,2008; Gan et al.,2011)。酵母雙雜交實驗得到GL1和TTG1可以結合到GL3和EGL3的不同區域,表明MYB-bHLH-WD40復合物可形成三聚體轉錄激活成分,并調控表皮毛發育的下游基因和形態發生(Pesch et al.,2015; Ramsay & Glover,2015)。GL2可通過反饋機制激活參與表皮毛成熟的TTG1復合體來切換MYB-bHLH-WD40復合體的功能。在表皮毛和根毛的形成過程中,兩個相鄰的細胞能夠相互競爭GL2/TTG1等表達調控因子(Pu et al.,2003)。
GL2在表皮毛形態發生中發揮著重要作用,包括細胞分枝、擴張和細胞壁成熟(Szymanski et al.,1998)。GL2是一種同源框基因,編碼含有StAR(steroidogenic acute regulatory protein)結構域的HD-Zip轉錄因子。這種同源框蛋白可在植物不同的發育過程中協調靶基因的表達(Rerie et al.,1994; Di & Al,1996)。GL2在成熟表皮毛中持續表達,并且是在早期形態發生中非根毛細胞和表皮毛分化所必需(Fyvie et al.,2015)。前人研究表明,GL2是調節根毛發育所必需的,并且優先在根的分生組織和伸長區域內的非毛發表皮細胞中表達(Masucci et al.,1996)。GL2突變體和GL2/GL3雙突變體均影響表皮毛的形態發生,使葉片上的表皮毛降解和減少,同時GL2突變體還具有種皮粘液缺乏、形成異位根毛等表型(Gao et al.,2008; Shi et al.,2012)。此外GL2突變體不能形成圍繞表皮毛的毛狀輔助細胞。遺傳實驗表明,GL2是GL1和TTG1的下游調控基因 (Pesch & Hülskamp,2011)。
本研究利用貴州省廣泛種植的‘貴農5號分離克隆與果刺發育有關的RrGL2,對RrGL2基因進行了生物信息分析和時空表達檢測,可為進一步研究刺梨果刺形成和發育的分子機制以及通過基因工程培育刺梨無刺果實提供了遺傳資源和理論基礎。
1 材料與方法
1.1 植物材料和細胞學分析
采集刺梨‘貴農5號的葉片和果實后,一部分立即在液氮中冷凍并儲存在-80 ℃。選擇葉、幼果和成熟果實并收集花后3周、5周和7周的果刺以檢測RrGL2的表達水平。另一部分通過體視顯微鏡(SZX7,OLYMPUS,Japan)觀察并拍照。
采摘不同時期的幼芽用FAA固定液固定,先在離心管中裝入3 mL固定液,將樣品放入固定液中,封口膜封口,用解剖針在封口膜上扎幾個小孔,然后抽真空,抽過真空后再加入2 mL固定液(樣品與固定液之比約為1∶20)。若長期保存應將固定液換成70%酒精,4 ℃冰箱保存。第1天用70%酒精過夜,第2天用85%酒精、95%酒精、無水酒精、無水酒精、1/5二甲苯、2/5二甲苯、3/5二甲苯、4/5二甲苯、純二甲苯進行梯度洗脫,純二甲苯處理后加碎蠟(放入36 ℃烘箱,放置3 d以上)洗滌。將已固著和修好的蠟塊裝在樣品固定器上,并固定好。將切片刀裝在切片機上。調節刀片的厚度為8~15 μm。
將切出的蠟帶,平展于盒內以供展片。之后經顯微鏡(BX53,Olympus,Japan)觀察,SPOT FLEXTM CCD拍攝(Diagnostic Instrument,USA)。
1.2 RNA提取和純化
使用Trizol試劑(TaKaRa,Japan)提取莖、葉片、花芽、種子和花后3周、5周和7周的皮刺部位的總RNA,具體步驟參考試劑盒說明。先用DNA酶(TaKaRa,Japan)處理RNA樣品。然后根據試劑盒說明使用oligo dT-接頭引物的RT-PCR Kit(TaKaRa,Japan)將提取的RNA進行逆轉錄。
1.3 分離GL2 cDNA
1.3.1 3′RACE的合成 利用3′ RACE(TaKaRa,Japan)試劑盒以刺梨葉片提取的RNA進行第一鏈cDNA合成。根據NCBI(https://www.ncbi.nlm.nih.gov/)下載的其他生物所報道的與果刺發育有關的GL2同源序列設計引物。用引物進行兩輪PCR擴增基因的3′ 末端(表1)。第一輪PCR:首先94 ℃變性3 min,然后進行20個循環的擴增(94 ℃ 30 s,55 ℃ 30 s,72 ℃ 2 min)和最后72 ℃延伸10 min。然后以第一輪PCR產物作為第二次PCR擴增的模板,并且在與第一輪PCR相同的條件下運行35個循環。
1.3.2 5′RACE的合成 根據SMARTerTM RACE cDNA擴增試劑盒(No.634923,Clontech),以葉片總RNA為模板合成cDNA第一鏈。按照上述3′末端的序列合成基因特異性引物(表1)進行Touch-down PCR:首先(94 ℃,30 s;72 ℃,90 s)5個循環,然后(94 ℃,30 s;70 ℃,30 s;72 ℃,1 min)5個循環,最后(94 ℃,30 s;55 ℃,2 min;72 ℃,2 min)30個循環。
1.4 克隆和測序
PCR產物用1%瓊脂糖凝膠檢測,挖膠回收后以瓊脂糖凝膠DNA純化試劑盒(DV805A,TaKaRa,Janpan)純化,克隆到pMD18-T載體(TaKaRa,Janpan)中,最后熱激法轉化大腸桿菌DH5α菌株(Trans,China)。陽性克隆由上海生工生物工程技術服務有限公司(中國上海)測序。
1.5 序列分析
使用NCBI ORF finder對PCR擴增獲得的GL2開放閱讀框片段進行分析。通過NCBI BLASTp獲得RrGL2同源蛋白。使用ProtParam分析蛋白質的分子量、等電點和氨基酸組成。用ProtScale的Kyte & Doolittle方法預測這些蛋白質的親水性或疏水性。TMHMM 2.0服務器(http://www.cbs.dtu.dk/services/ TMHMM/)用于預測蛋白質的跨膜結構域。SMART在線預測蛋白質結構域。ESPript 3.0(http://espript.ibcp.fr/ESPript/ESPript/)預測蛋白質二級結構。Mega6.0軟件Neighbor-Joining法用于構建系譜樹。
1.6 基因表達檢測
使用FastStart DNA Master SYBR Green I試劑盒在LightCycler 480儀器(Roche,Switzerland)中進行RrGL2的qRT-PCR檢測。β-actin作為內參基因。引物為RrGL2,Forward (5′-3′):CGAGGCAGTGACAGTGAAGG; Reverse (5′-3′):GGCAGACTCAACAGACTCCATAG。β-actin Forward (5′-3′):CCGCCATGTA TGTTGCCATCC; Reverse (5′-3′):AGCCAGGTCAAGACGCAGAAT。
qRT-PCR程序根據SYBR說明書進行,擴增進行40個循環:在95 ℃變性30 s,在55 ℃退火30 s,并在72 ℃延伸1 min。相對于對照的表達水平通過計算△△Ct[(△△Ct=sample △Ct-control △Ct),△Ct= RrGL2 Ct -(β-actin)Ct],然后用2-△△Ct方法分析。
1.7 統計分析
本研究中的所有數據均為三次生物學重復的平均值和相應的標準偏差表示。使用SPSS單因素方差分析方法分析獲得的數據,Duncan多重檢驗以比較統計顯著性差異(P<0.05)。
2 結果與分析
2.1 刺梨不同組織的觀察和早期細胞學研究
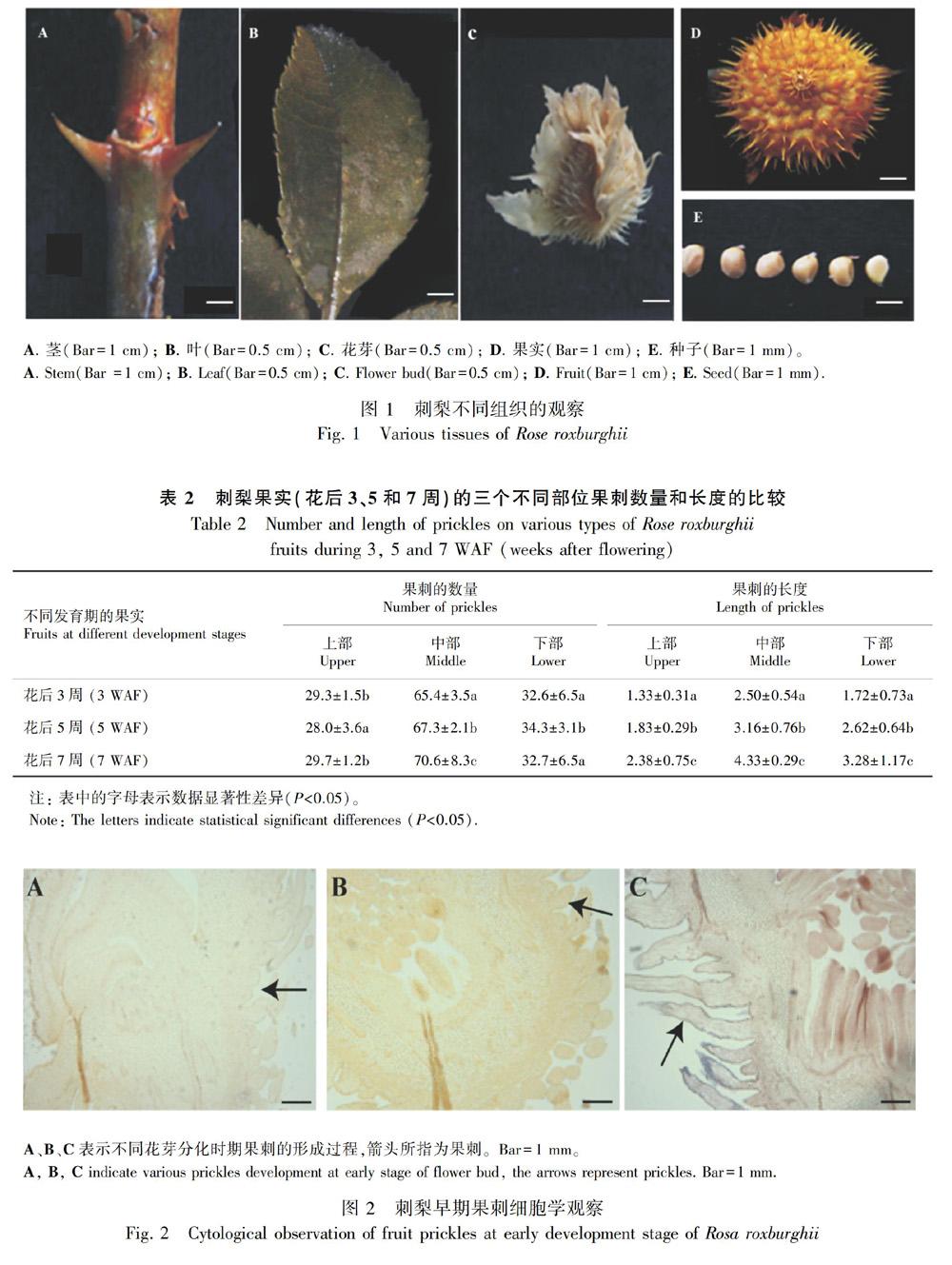
分別選取刺梨的不同部位觀察,如圖1所示,包括的組織有莖(圖1:A)、葉(圖1:B)、花芽(圖1:C)、果實(圖1:D)和種子(圖1:E)。在莖和果實的組織表面上觀察到有一些堅硬的刺,分別為枝刺和果刺。在花芽最外的部位也存在少量刺的形態,葉片和種子外表面沒有出現堅硬的刺。
進一步選取不同發育階段的果實進行果刺數量和長度的統計,如表2所示。選取花后3、5和7周形成的果實,此時刺梨果實的直徑分別是1.5、2.3和3.4 cm。將果實分為三個部位進行統計,分別為上部、中部和下部。從表2可以得到,在不同的發育時期,果刺的數量均主要集中在中部,上部和下部數量較少。隨著果實的發育,三個部位果刺的長度均發生顯著性的增加。
刺梨經過一定時間的營養生長后,在外界各因素達到一定要求時進入生殖生長階段,這是刺梨果實形成的必經階段。當刺梨進入生殖生長時期,刺梨莖端分生組織細胞開始分裂分化,在花的發育過程中刺梨萼片上的刺細胞也開始不斷發生。花的發育是一個動態的過程,最初觀察到花萼上有少量已經形成的刺結構,隨著花的發育,萼片上刺結構會成批次的產生,逐漸增多(圖2:A)。當花的結構慢慢向外展開時,在花瓣原基基部處,雄蕊原基開始慢慢發育形成一些粗而短的近似橢圓形的結構,刺原基突起內的基本分生組織細胞進行切向分裂,細胞數目開始增加,表皮上的細胞也進行分裂以適應刺的向外延伸(圖2:B)。基部內的細胞迅速分裂使基部擴展變大,中部細胞增多,形態也發生變化,橢圓形的細胞開始變細、變長,尖端呈“針”狀型結構,刺結構的整體外形初步形成(圖2:C)。
2.2 刺梨RrGL2的序列分析
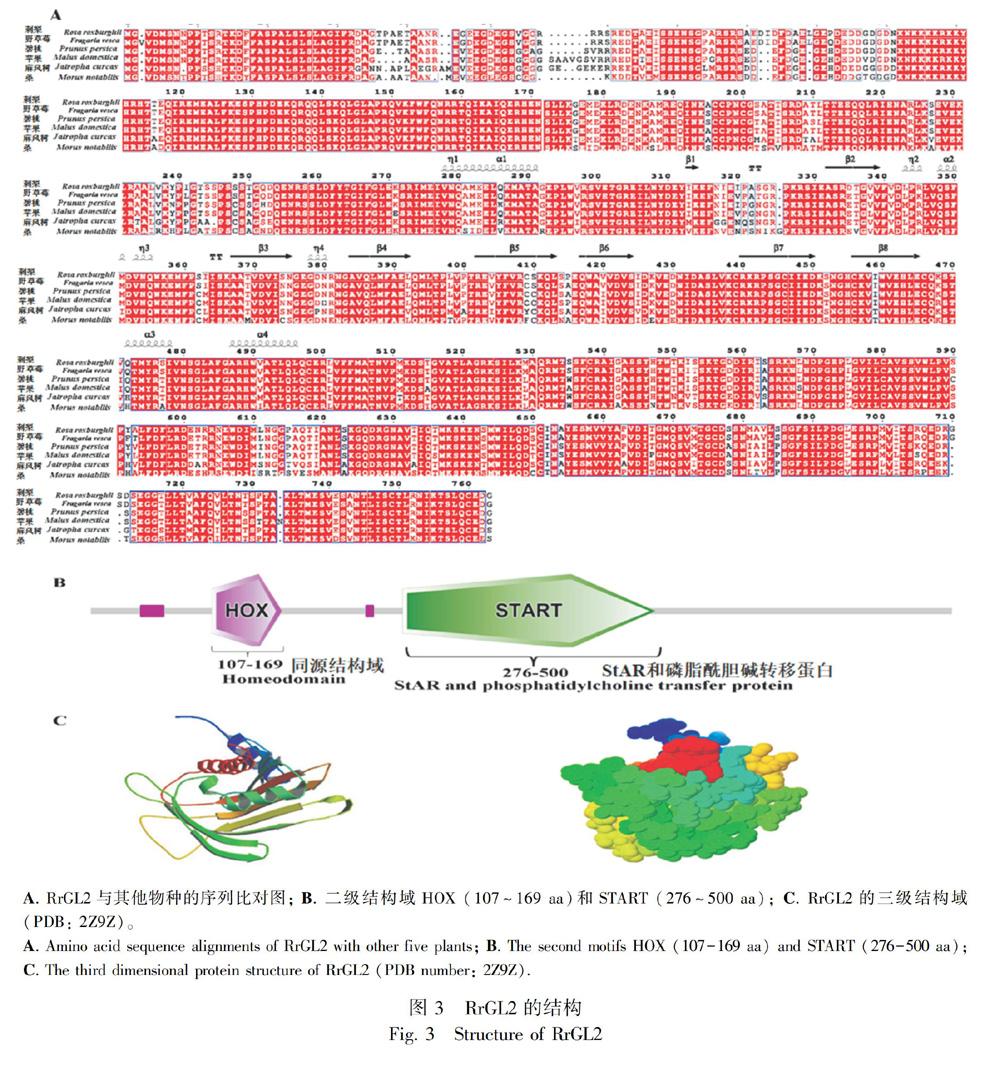
以‘貴農5號的RNA為模板,通過RACE方法得到RrGL2的全長cDNA。RrGL2(Genebank登錄號:MG386498)的全長為2 292 bp,編碼763個氨基酸。BLAST分析表明,刺梨的RrGL2序列和野草莓(Fragaria vesca)、碧桃(Prunus persica)、蘋果(Malus domestica)、麻風樹(Jatropha curcas)和桑(Morus notabilis)GL2具有較高的同源性,分別是96%、88%、86%、78%和80%的相似性。
ProtParam分析RrGL2得到其分子量為8.49 kDa。RrGL2的等電點是5.73。 RrGL2含有13.76%的酸性氨基酸、13.63%的堿性氨基酸、38.79%的疏水性氨基酸、27.39%的帶電荷氨基酸和61.07%的極性氨基酸。ProtScale分析RrGL2得到其親水性氨基酸的數量明顯大于疏水性氨基酸的數量,且親水性的平均值為-0.525,表明RrGL2編碼的蛋白質為可溶性蛋白質。TMHMM2.0預測表明RrGL2不含有跨膜結構域。
2.3 RrGL2的同源蛋白和系譜樹分析
將不同物種的GL2進行比對分析(圖3:A)。BLAST分析表明,RrGL2和野草莓(Fragaria vesca)、碧桃(Prunus persica)、蘋果(Malus domestica)、麻風樹(Jatropha curcas)和桑(Morus notabilis)的蛋白質序列相似性較高,分別是98%、92%、90%、83%和82%相似性。
基于RrGL2的三維結構(PDB編號:2Z9Z),利用ESPript 3.0得到蛋白質的二級結構和三級結構。如圖3:A所示,β-轉角是序列的主要二級結構。多序列比對的結果表明,RrGL2與其他物種的二級結構域也較為保守。SMART分析顯示RrGL2的結構域包括兩個主要結構域,分別是位于107~169氨基酸的同源結構域HOX(homeobox)和位于276~500氨基酸的START(StAR和磷酰膽堿轉移蛋白)(圖3:B)。圖3:C以兩種不同的方式顯示了RrGL2三維結構,在不同的折疊處,包括轉角和不規則的卷曲均可發生與DNA/RNA的結合。使用Mega 6.0軟件構建RrGL2的系統進化樹圖,得到刺梨與其他植物的親緣關系,如圖4所示。圖中顯示RrGL2與野草莓中的GL2序列最相似。除此之外,RrGL2與其他薔薇科植物的親緣關系也較近,如桃、櫻桃、梅和蘋果。
2.4 RrGL2在不同組織和果實發育的表達模式
利用Real-time PCR獲得RrGL2在不同組織中的表達水平,結果如圖5所示。RrGL2在種子中的表達量最低,莖和果實的RrGL2表達量分別是種子的59.95倍和33.52倍。葉和花中RrGL2的表達量分別是種子中的7.99倍和15.62倍。
從圖5:B中可以看出,花后3、5和7周的果刺中RrGL2表達量。結果表明隨著果實的發育和果刺的增加,RrGL2表達量逐漸增加,在花后7周果刺中RrGL2表達量最高,分別是花后3周和5周果刺中的7.87倍和2.10倍,可以推斷RrGL2在果刺的形成中發揮一定的作用。
3 討論
形態觀察結果發現刺梨莖上有一些堅硬的刺,果實外表面上密披果刺。通過細胞學觀察得到,果刺在花芽階段已經形成。同時,RrGL2在莖和果中的表達水平高于其他組織,說明RrGL2可能在果刺和表皮毛形態發生中發揮重要作用。研究表明RrGL2作用在果實發育早期的果刺中檢測到RrGL2的表達,證明它可能在果實發芽階段起作用(Ohashi et al.,2002; Vernoud et al.,2009)。隨著果實的成熟,果刺的數量和長度都增加。與形態學改變相對應,RrGL2的表達水平也隨之提高。這些結果暗示RrGL2可能與刺梨果刺發育起始密切相關(Rerie et al.,1994)。而在花后7周的果實果刺中RrGL2表達量達到最高,暗示與刺發育相關的RrGL2基因能夠增加果刺的形成。轉錄因子EGL3和TTG1參與GL2的表達調控,最終導致表皮毛形成(Song et al.,2015)。MYB類蛋白如GL1和WER,可通過與bHLH蛋白相互作用來調節GL2同源框基因在特定部位的表達。同時GL2的表達還受到CAPRICE MYB的抑制(Lee & Schiefelbein,1999)。RrGL2作為一個關鍵基因和其他參與調節刺梨果刺發育的基因功能還需要進一步的功能分析確定。
本研究通過RACE技術從刺梨中克隆得到RrGL2。在NCBI blast數據庫中搜索獲得RrGL2與其他物種的氨基酸結構域的高度相似性的GL2類蛋白成員,表明RrGL2可能與這些物種中的GL2具有類似功能,參與調節果刺的起始,形態發生和發育(Wang et al.,1999)。結構域分析顯示RrGL2是一個Homeodomain蛋白。Homeodomain蛋白可結合DNA并調節轉錄DNA模板,參與形成多蛋白復合物從而調控發育基因的表達(Foronda et al.,2009)。RrGL2的三級結構是DNA/RNA結合螺旋結構和orthogonal結構,這也暗示RrGL2可以與DNA相互作用并控制。同時,GL1和GL3轉錄因子的相互作用激活GL2的表達(Wang & Chen,2008)。據報道GL2以細胞位置依賴性方式在根毛分化形成過程中調節根毛發育,并影響種子含油量(Masucci et al.,1996; Shen et al.,2006)。
本研究通過生物信息學的分析方法得到RrGL2調節蛋白與其他物種中的直系同源物,展現GL2中正向進化選擇的潛力。利用時空表達的檢測獲得RrGL2在不同組織和果刺不同發育階段的相對表達水平。因此,本文為揭示和理解參與刺梨果刺發育的基因以及果刺發育分子機制提供了分子基礎,為通過基因工程培育更少或甚至無刺刺梨提高理論基礎。
參考文獻:
AN L,ZHOU Z,YAN A,et al.,2011. Progress on trichome deve-lopment regulated by phytohormone signaling[J]. Plant Signal Behav,6(12):1959-1962.
DI CM,AL E,1996. The Arabidopsis Athb-10 (GLABRA2) is an HD-Zip protein required for regulation of root hair deve-lopment[J]. Plant J Cell Mol Biol,10(3):393-402.
FENG LG,LUAN XF,WANG J,et al.,2015. Cloning and expression analysis of transcription factor Rrttg1 related to prickle development in rose (Rosa Rugosa)[J]. Arch Biol Sci,67(4):1219-225.
FORONDA D,NAVAS LF,GARAULET DL,et al.,2009. Function and specificity of Hox genes[J]. Int J Dev Biol,53(8-10):1404-1419.
FYVIE MJ,MURRAY JA,KILBY NJ,2015. Mosaic analysis of GL2 gene expression and cell layer autonomy during the specification of Arabidopsis leaf trichomes[J]. Genesis,28(2): 68-74.
GAN L,XIA K,CHEN JG,et al.,2011. Functional characte-rization of trichomeless 2,a new single-repeat R3 MYB transcription factor in the regulation of trichome patterning in Arabidopsis[J]. Bmc Plant Biol,11(1):176-187.
GAO Y,GONG X,CAO W,et al.,2008. SAD2 in Arabidopsis functions in trichome initiation through mediating GL3 function and regulating GL1,TTG1 and GL2 expression[J]. J Integr Plant Biol,50(7):906-917.
GOMES S,BACELAR E,MARTINSLOPES P,et al.,2012. Infection process of olive fruits by Colletotrichum acutatum and the protective role of the cuticle and epidermis[J]. J Agric Sci,4(2):101-110.
KELLOGG AA,BRANAMAN TJ,JONES NM,et al.,2011. Morphological studies of developing rubus prickles suggest that they are modified glandular trichomes[J]. Botany,89(4):217-226.
LEE MM,SCHIEFELBEIN J,1999. Werewolf,a MYB-related protein in Arabidopsis,is a position-dependent regulator of epidermal cell patterning[J]. Cell,99(5):473-477.
LIU W,LI S Y,HUANG XE,et al.,2012. Inhibition of tumor growth in vitro by a combination of extracts from Rosa roxburghii Tratt and Fagopyrum cymosum[J]. Asian Pac J Cancer Prev,13(5):2409-2414.
MASUCCI JD,RERIE WG,FOREMAN DR,et al.,1996. The homeobox gene glabra 2 is required for position-dependent cell differentiation in the root epidermis of Arabidopsis tha-liana[J]. Development,122(4):1253-1260.
OHASHI Y,OKA A,RUBERTI I,et al.,2002. Entopically additive expression of glabra2 alters the frequency and spa-cing of trichome initiation[J]. Plant J,29(3):359-369.
PESCH M,HLSKAMP M,2011. Role of triptychon in trichome patterning in Arabidopsis[J]. Bmc Plant Biol,11(1): 130-138.
PESCH M,SCHULTHEI I,KLOPFFLEISCH K,et al.,2015. Transparent testa glabra1 and glabra1 compete for binding to glabra3 in Arabidopsis[J]. Plant Physiol,168(2): 584-597.
PU L,SUO J,XUE Y,2003. Molecular control of plant trichome development[J]. Acta Genet Sin,30 (11):1078-1084.
RAMSAY NA,GLOVER BJ,2005. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity[J]. Trends Plant Sci,10(2):63-70.
RERIE WG,FELDMANN KA,MARKS MD,1994. The glabra2 gene encodes a homeo domain protein required for normal trichome development in Arabidopsis[J]. Gene Dev,8 (12):1388-1399.
SHEN B,SINKEVICIUS KW,SELINGER DA,et al.,2006. The homeobox gene glabra2 affects seed oil content in Arabidopsis[J]. Plant Mol Biol,60(3):377-387.
SHI L,KATAVIC V,YU Y,et al.,2012. Arabidopsis glabra2 mutant seeds deficient in mucilage biosynthesis produce more oil[J]. Plant J Cell Mol Biol,69(1):37-46.
SONG SK,KWAK SH,CHANG SC,et al.,2015. Werewolf and enhancer of glabra3 are interdependent regulators of the spatial expression pattern of glabra2 in Arabidopsis[J]. Biochem Biophys Res Comm,467(1):94-100.
SZYMANSKI DB,JILK RA,POLLOCK SM,et al.,1998. Control of GL2 expression in Arabidopsis leaves and trichomes[J]. Development,125(7):1161-1171.
VAN FH,VAN CS,RAUTENBACH GS,et al.,2008. In vitro antioxidant,antimutagenic and genoprotective activity of Rosa roxburghii fruit extract[J]. Phyto Res,22(3):376-383.
VERNOUD V,LAIGLE G,ROZIER F,et al.,2009. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize[J]. Plant J,59(6): 883-894.
WANG S,CHEN JG,2008. Arabidopsis transient expression analysis reveals that activation of glabra2 may require concurrent binding of glabra1 and glabra3 to the promoter of glabra2[J]. Plant Cell Physiol,49(12):1792-1804.
WANG YL,NIE JT,CHEN HM,et al.,2016. Identification and mapping of Tril,a homeodomain-leucine zipper gene involved in multicellular trichome initiation in Cucumis sativus[J]. Theor Appl Genetics,129(2):305-316.
XU P,ZHANG WB,CAI XH,et al.,2014. Flavonoids of Rosa roxburghii Tratt act as radioprotectors[J]. Asian Pac J Cancer Prev,15(23):8171-175.
ZHAO M,MOROHASHI K,HATLESTAD G,et al.,2008. The TTG1-bHLH-MYB complex controls trichome cell fate and patterning through direct targeting of regulatory loci[J]. Development,135(11):1991-1999.
(責任編輯 周翠鳴)
