長(zhǎng)林4號(hào)油茶幼苗對(duì)低溫脅迫的生理響應(yīng)
2020-03-31 12:23:41胡倩裴云霞倪天虹耿若楠張娟娟余金金杜克兵周忠誠(chéng)
湖北林業(yè)科技 2020年6期
胡倩 裴云霞 倪天虹 耿若楠 張娟娟 余金金 杜克兵 周忠誠(chéng)

摘 要: 為探究油茶幼苗對(duì)低溫脅迫的生理生化響應(yīng)機(jī)制,本研究以湖北省主栽油茶品種長(zhǎng)林4號(hào)的2 a生幼苗為試驗(yàn)材料,研究了其表型、葉片氣體交換參數(shù)以及相關(guān)生理指標(biāo)對(duì)低溫脅迫(4 ℃和-4 ℃)的響應(yīng)。結(jié)果表明:①低溫脅迫顯著影響了長(zhǎng)林4號(hào)油茶幼苗的形態(tài)特征,使嫩莖和葉片出現(xiàn)不同程度的萎蔫和失水癥狀,并且-4 ℃處理比4 ℃處理植株受損程度更嚴(yán)重。②低溫脅迫顯著降低了油茶葉片的氣體交換參數(shù)(Pn、Gs、Tr、RWUEi)值,恢復(fù)期間其值仍持續(xù)下降,且-4 ℃處理的抑制作用顯著強(qiáng)于4 ℃處理。③低溫脅迫雖然降低了油茶葉片的SPAD值,但未達(dá)到顯著水平。④低溫脅迫顯著提高了油茶葉片中的MDA、Pro和可溶性糖含量。恢復(fù)期間,4 ℃處理的MDA和Pro含量略有下降,而可溶性糖含量仍繼續(xù)增加;-4 ℃處理的MDA和可溶性糖含量略有下降,Pro含量仍繼續(xù)增加。(5)低溫脅迫顯著影響了油茶葉片中的抗氧化酶活性。隨著脅迫時(shí)間的延長(zhǎng),SOD和POD活性持續(xù)上升,而CAT活性則先上升后下降,并且-4 ℃處理的上升幅度明顯高于4 ℃處理;恢復(fù)期間,抗氧化酶活性均有不同程度的降低。
關(guān)鍵詞: 油茶;低溫脅迫;表型;生理生態(tài)特征
中圖分類號(hào):S794.4 ? 文獻(xiàn)標(biāo)識(shí)碼:A ? 文章編號(hào):1004-3020(2020)06-0005-08
Abstract: In order to explore the physiological and biochemical response mechanism of C.oleifera seedlings to low temperature stress,this experiment took the 2-year-old seedings of C.oleifera‘Changlin4 as the experimental material to study the response of its phenotype、leaf gas exchange parameters and related physiological indicators under cold stress (4 ℃and -4 ℃).The results showed that: ① Cold stress significantly affected the morphological characteristics of C.oleifera ‘Changlin4,causing wilting and water loss symptoms of the young stems and leaves in different degrees,and the degree of plant damage was more serious under -4 ℃ treatment than under 4℃ treatment.② Cold stress significantly reduced the values of gas exchange parameters (Pn、Gs、Tr、RWUEi) in the leaves of C.oleifera,which continued to decline during the recovery period,and the inhibition effect of -4 ℃ treatment was significantly stronger than that of 4 ℃.③ Although cold stress reduced the SPAD value of C.oleifera,it did not reach a significant level.(4) Cold stress significantly increased the content of MDA,Pro and soluble sugar in the leaves of C.oleifera.During the recovery period,the MDA and Pro content of 4 ℃ treatment decreased slightly,while the soluble sugar content continued to increase;the MDA and soluble sugar content of -4 ℃ treatment decreased slightly,while the Pro content continued to increase.(5) Cold stress significantly affected the activity of antioxidant enzymes in the leaves of C.oleifera.With the extension of stress time,SOD and POD activities continued to increase,while CAT activities first increased and then decreased,and the increase of -4 ℃ treatment was significantly higher than that of 4 ℃;during the recovery period,the activity of antioxidant enzymes decreased in varying degrees.
Key words: Camellia oleifera;cold stress;phenotype;eco-physiological characteristics
油茶Camellia oleifera是山茶科Theaceae山茶屬Camellia中油脂含量較高且具有經(jīng)濟(jì)價(jià)值的一類植物的總稱,被譽(yù)為“東方橄欖油”[1]。油茶在中國(guó)已經(jīng)有2000多年的栽培歷史,其適應(yīng)能力強(qiáng),經(jīng)濟(jì)價(jià)值高,是一種常綠闊葉的經(jīng)濟(jì)樹種[2]。截止2018年,中國(guó)現(xiàn)有油茶林已達(dá)到436.67萬hm2,主要分布在江西、湖南、浙江、福建、廣西等省區(qū)[3-4]。湖北省是中國(guó)油茶北部中心產(chǎn)區(qū),也是油茶生長(zhǎng)的邊緣地區(qū),主要栽培的油茶有長(zhǎng)林4號(hào)C.oleifera ‘Changlin4、長(zhǎng)林18號(hào)C.oleifera ‘Changlin18、長(zhǎng)林23號(hào)C.oleifera ‘Changlin23、長(zhǎng)林40號(hào)C.oleifera ‘Changlin 40、谷城大紅果8號(hào)C.oleifera ‘Guchengdahongguo8等品種[5]。近年來,湖北省油茶產(chǎn)業(yè)發(fā)展迅速,但是低溫對(duì)油茶栽培造成了許多不利影響,常常導(dǎo)致植株幼苗出現(xiàn)不同程度的凍害現(xiàn)象,造成巨大的經(jīng)濟(jì)損失。
目前,中國(guó)科研工作者在油茶良種選育、分子育種、栽培技術(shù)以及茶油提取等方面做出了大量的研究[6-8]。關(guān)于油茶低溫脅迫的研究雖有報(bào)道,但針對(duì)長(zhǎng)林4號(hào)油茶品種抗寒性生理的研究卻較少。為了探究油茶對(duì)低溫逆境的生理響應(yīng),本研究以長(zhǎng)林4號(hào)油茶為試驗(yàn)材料,針對(duì)低溫脅迫對(duì)其表型、葉片氣體交換參數(shù)以及相關(guān)生理指標(biāo)的影響進(jìn)行了研究,以期為湖北省油茶栽培提供參考。
1 材料和方法
1.1 試驗(yàn)材料
試驗(yàn)材料為2 a生長(zhǎng)林4號(hào)C.oleifera‘Changlin4的嫁接苗(以油茶實(shí)生苗為砧木)。
1.2 試驗(yàn)方法
1.2.1 植株培養(yǎng)
于2017年秋季,將1 a生油茶嫁接苗移栽于規(guī)格為10.5 cm×8.5 cm×20.0 cm的營(yíng)養(yǎng)缽中,置于室內(nèi)培養(yǎng),室內(nèi)溫度為(25±2)℃,光照強(qiáng)度為500 μmol·(m2·s)-1,光照時(shí)間為16 h·d-1,光源為白色冷光燈,室內(nèi)空氣相對(duì)濕度為70%~80%。苗木培養(yǎng)基質(zhì)為蛭石和營(yíng)養(yǎng)土(1︰3,v/v)的混合物,其pH值為5.5,基質(zhì)中含有大量的N、P2O5、K2O(2%~5%)和有機(jī)質(zhì)(>20%)。植株隨機(jī)排列,培養(yǎng)期間進(jìn)行常規(guī)管理。2018年9月,當(dāng)植株高度達(dá)到30~35 cm、地徑為3.0~3.5 cm時(shí),選取大小、長(zhǎng)勢(shì)基本一致的苗木進(jìn)行低溫脅迫試驗(yàn)。
1.2.2 低溫處理
試驗(yàn)共設(shè)3個(gè)溫度處理(25、4、-4 ℃),采用LED智能光照培養(yǎng)箱(武漢瑞華儀器設(shè)備有限公司)進(jìn)行試驗(yàn)。將植株分別置于3個(gè)不同的培養(yǎng)箱內(nèi),隨機(jī)排列,溫度分別設(shè)為25 ℃、4 ℃和-4 ℃,培養(yǎng)箱內(nèi)的其他環(huán)境條件與培養(yǎng)室保持一致。在低溫分別為4 ℃和-4 ℃的處理過程中,將箱內(nèi)溫度進(jìn)行每分鐘降低2 ℃的降溫處理,直至達(dá)到所設(shè)定的溫度后開始計(jì)算低溫脅迫的時(shí)間。
1.2.3 測(cè)定方法
將每個(gè)溫度處理下的植株隨機(jī)分為2組:一組用于表型、葉片氣體交換參數(shù)、葉綠素含量(SPAD值)等無損傷性指標(biāo)的觀測(cè);另一組用于丙二醛(MDA)、脯氨酸(Pro)、可溶性糖、超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)等生理指標(biāo)的測(cè)定。具體試驗(yàn)和觀測(cè)方法如下:
4 ℃和-4 ℃處理:先將植株分別置于4 ℃和-4 ℃條件下處理72 h,隨后移至25 ℃的培養(yǎng)箱內(nèi)恢復(fù)24 h,共處理96 h。期間,分別于4 ℃和-4 ℃處理的前1天(處理0 h)、處理6、24、48、72 h和恢復(fù)24 h(處理96 h)時(shí)觀測(cè)植株的表型變化、葉片氣體交換參數(shù)和SPAD值。同時(shí)采集植株葉片,于液氮中速凍后于-80 ℃下保存,用于MDA、Pro、可溶性糖含量以及SOD、POD、CAT活性等生理指標(biāo)的測(cè)定,3次重復(fù)/處理。
25 ℃處理:將植株置于25 ℃培養(yǎng)箱下培養(yǎng),然后分別于處理0、6、24、48、72 h和恢復(fù)24 h(處理96 h)時(shí),進(jìn)行上述指標(biāo)的觀測(cè)與樣品采集,3次重復(fù)/處理。
相關(guān)指標(biāo)的觀測(cè)方法為:
(1)葉片氣體交換參數(shù)與SPAD值的測(cè)定:固定選取植株頂端完全展開的第4~5片嫩葉進(jìn)行測(cè)定。葉片氣體交換參數(shù)測(cè)定時(shí)間為上午10∶00~12∶00,采用LI-6400光合測(cè)定儀(LI-COR Inc.,Lincoln,NE,USA)測(cè)定。測(cè)定時(shí),采用標(biāo)準(zhǔn)LI-COR葉室,紅藍(lán)光源(6400-02 LED光源),設(shè)定光照強(qiáng)度為1500 μmol/(m2·s),空氣流速為500 μmol/s,測(cè)定指標(biāo)主要包括凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導(dǎo)度(Gs)、胞間CO2濃度(Ci)和環(huán)境CO2濃度(Ca),以及相應(yīng)的周圍環(huán)境條件,如葉面溫度、大氣相對(duì)濕度、光合有效輻射等。瞬時(shí)水分利用效率(RWUEi)的計(jì)算公式[9]為:RWUEi=Pn/Gs。采用SPAD-502plus葉綠素測(cè)定儀(Konica Minolta Inc,Japan)測(cè)定葉片相同部位的SPAD值。每個(gè)葉片測(cè)定5次,取其平均值作為該葉片的SPAD值。
(2)生理指標(biāo)測(cè)定:采用TBA法(硫代巴比妥酸)測(cè)定MDA含量;采用酸性茚三酮法測(cè)定Pro含量;采用蒽酮比色法測(cè)定可溶性糖含量;采用NBT(氮藍(lán)四唑)光化學(xué)還原法測(cè)定SOD活性;采用愈創(chuàng)木酚法測(cè)定POD活性;采用高錳酸鉀滴定法測(cè)定CAT活性[10]。
1.3 數(shù)據(jù)分析
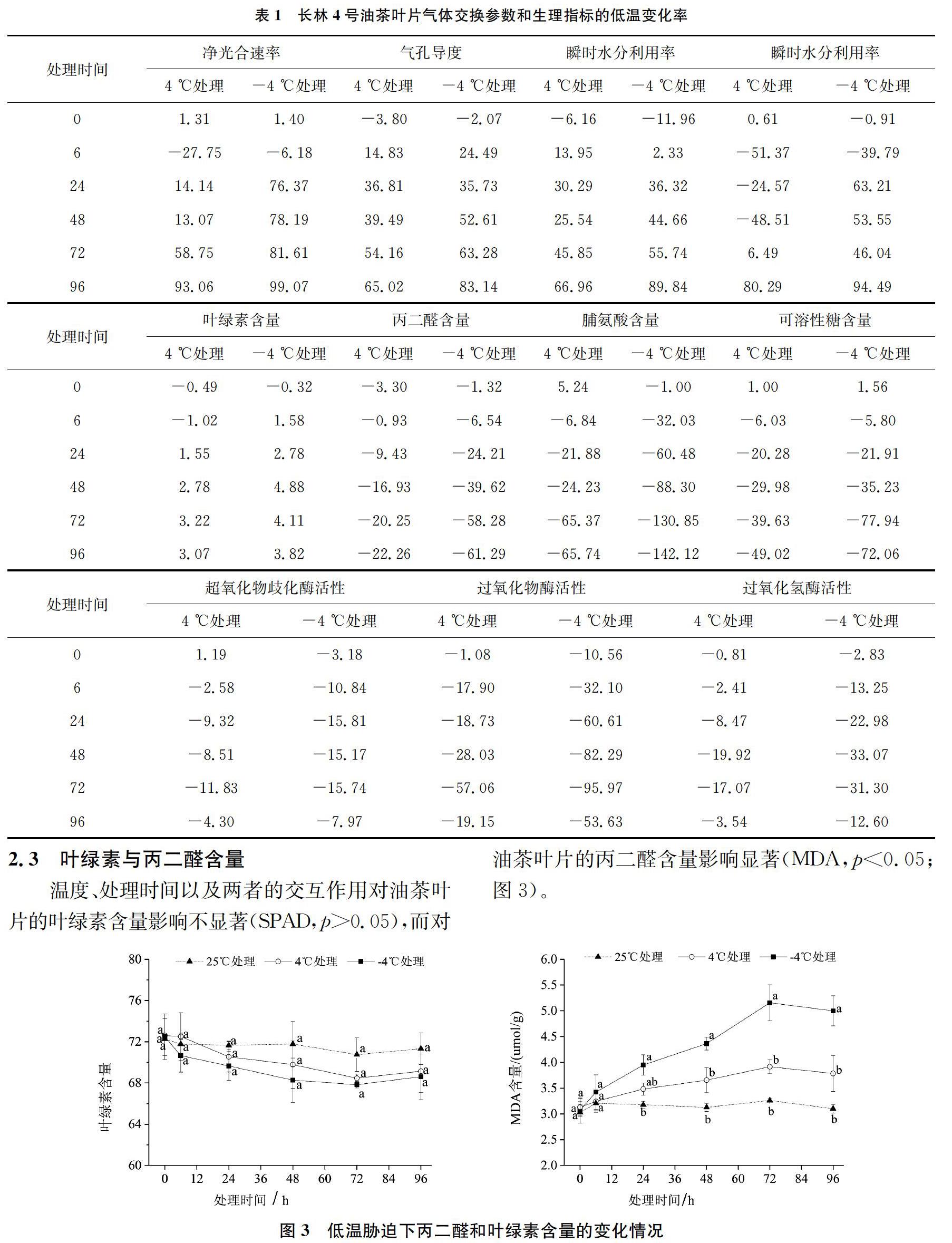
圖中數(shù)據(jù)均為平均值±標(biāo)準(zhǔn)誤(n=3)。方差分析、多重比較(Duncans)采用SAS8.1軟件(SAS Institute Inc.,Cary,NC,USA)進(jìn)行。采用低溫變化率評(píng)價(jià)低溫處理對(duì)各個(gè)指標(biāo)的影響程度。低溫變化率=(25 ℃對(duì)照值-低溫處理值)/25 ℃對(duì)照值×100%。
2 結(jié)果與分析
2.1 植株形態(tài)
低溫脅迫下植株的表型變化特征如圖1所示。25 ℃處理下,植株葉片和嫩莖的生長(zhǎng)都正常,均未出現(xiàn)受害癥狀。4 ℃處理下,處理前期植株均未出現(xiàn)受害癥狀;至72 h時(shí),所有植株下部成熟葉片未出現(xiàn)明顯受害癥狀,而其上部嫩葉和嫩莖有不同程度的受害癥狀,主要表現(xiàn)為葉片萎蔫和失水,恢復(fù)期間略有好轉(zhuǎn)。與4 ℃處理相比,-4 ℃處理下植株的受害程度更加嚴(yán)重。在處理48 h時(shí),植株上部嫩葉開始出現(xiàn)萎蔫的癥狀;至72 h時(shí),其嫩莖、嫩葉和下部成熟葉均出現(xiàn)明顯的變褐、萎蔫和失水癥狀,恢復(fù)期間未出現(xiàn)明顯好轉(zhuǎn)。可見,-4 ℃處理72 h已經(jīng)對(duì)長(zhǎng)林4號(hào)油茶幼苗造成了較為嚴(yán)重的傷害。
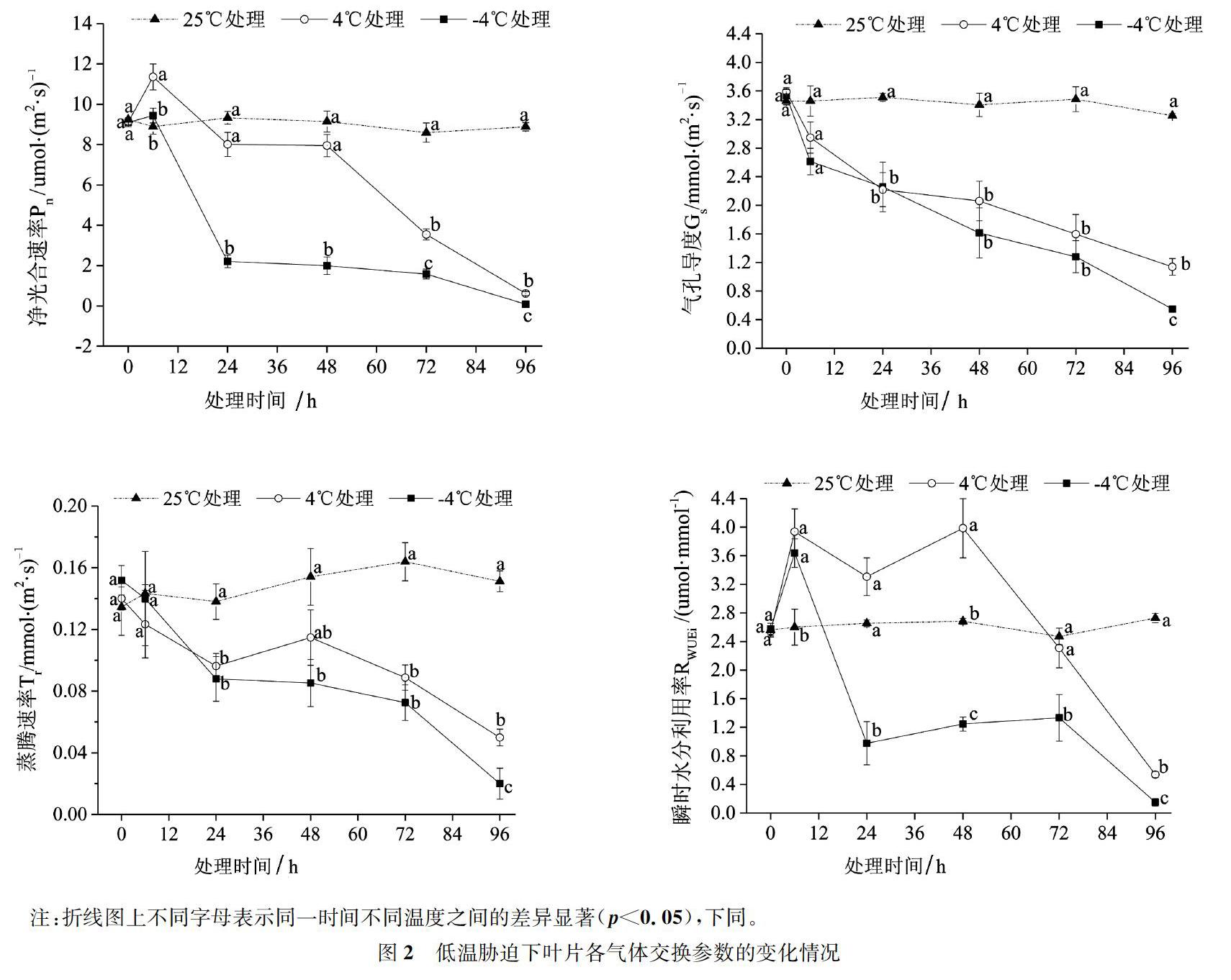
2.2 葉片氣體交換參數(shù)
溫度、處理時(shí)間以及兩者的交互作用顯著影響了油茶葉片的凈光合速率(Pn,p<0.05;圖2)、氣孔導(dǎo)度(Gs,p<0.05)、蒸騰速率(Tr,p<0.05)和瞬時(shí)水分利用率(RWUEi,p<0.05)。
對(duì)于Pn,25 ℃條件下,Pn值比較穩(wěn)定,無明顯變化(圖2)。4 ℃和-4 ℃條件下,Pn在6 h時(shí)略有上升,隨后開始下降,恢復(fù)期間仍持續(xù)降低,并于96 h時(shí)達(dá)到最低值,在6 h-24 h期間Pn值下降速度最快。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)下降的幅度更大。至72 h時(shí),-4℃和4℃處理的低溫變化率分別達(dá)到81.61%和58.75%;至96 h時(shí),分別達(dá)到99.07%和93.06%(表1)。
對(duì)于Gs,25 ℃條件下,其值比較穩(wěn)定,無明顯變化(圖2)。4 ℃和-4 ℃條件下,Gs呈下降趨勢(shì),恢復(fù)期間仍持續(xù)降低,并于96 h時(shí)達(dá)到最低值,其中0~6 h期間Gs值下降速度最快。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)下降的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到63.28%和54.16%;至96 h時(shí),分別達(dá)到83.14%和65.02%(表1)。
Tr的變化趨勢(shì)與Gs相似,在25 ℃條件下,其值比較穩(wěn)定,變化幅度不大(圖2)。4 ℃條件下,Tr值除了在48 h時(shí)有小幅上升之外,試驗(yàn)期間持續(xù)降低,并于96 h時(shí)達(dá)到最低值。-4 ℃條件下,Tr在整個(gè)試驗(yàn)期間持續(xù)降低,并于96 h時(shí)達(dá)到最低值,在6~24 h期間Tr下降速度最快。與4 ℃相比,-4 ℃處理隨著脅迫時(shí)間的延長(zhǎng)下降的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到55.74%和45.85%;至96 h時(shí),分別達(dá)到了89.84%和66.96%(表1)。
對(duì)于RWUEi,其值在25 ℃條件下比較穩(wěn)定,無明顯變化(圖2)。4 ℃條件下,RWUEi在6 h時(shí)明顯上升,隨后下降,在48 h時(shí)又有小幅上升,隨后持續(xù)下降,并于96 h時(shí)達(dá)到最低值,在48~72 h期間RWUEi值下降速度最快。-4 ℃條件下,RWUEi在6 h時(shí)明顯上升,隨后下降,在48~72 h時(shí)又有小幅上升,恢復(fù)期間開始下降,并于96 h時(shí)達(dá)到最低值,其中6~24 h期間RWUEi值下降速度最快。與4 ℃相比,-4 ℃處理對(duì)RWUEi的抑制作用更加顯著。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到46.04%和6.49%;至96 h時(shí),分別達(dá)到94.49%和80.29%(表1)。
2.3 葉綠素與丙二醛含量
溫度、處理時(shí)間以及兩者的交互作用對(duì)油茶葉片的葉綠素含量影響不顯著(SPAD,p>0.05),而對(duì)油茶葉片的丙二醛含量影響顯著(MDA,p<0.05;圖3)。
對(duì)于SPAD,25 ℃條件下,SPAD值比較穩(wěn)定,無明顯變化(圖3)。4 ℃和-4 ℃條件下,SPAD值均呈持續(xù)下降趨勢(shì),但是下降幅度不大,在72 h時(shí)達(dá)到最低值,恢復(fù)期間略有上升。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)下降的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到4.11%和3.22%;至96 h時(shí),分別達(dá)到3.82%和3.07%(表1)。
就MDA而言,25 ℃條件下,其值比較穩(wěn)定,變化幅度不大(圖3)。4 ℃和-4 ℃條件下,MDA含量均呈上升趨勢(shì),并于72 h時(shí)達(dá)到最大值,恢復(fù)期間略有下降,其中48~72 h期間MDA含量上升速度最快。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)上升的幅度更大。至72 h時(shí),-4℃和4℃處理的低溫變化率分別達(dá)到58.28%和20.25%;至96 h時(shí),分別達(dá)到61.29%和22.26%(表1)。
2.4 脯氨酸與可溶性糖含量
溫度、處理時(shí)間以及兩者的交互作用顯著影響了油茶葉片的脯氨酸(Pro,p<0.05;圖4)和可溶性糖含量(p<0.05)。
Pro的變化趨勢(shì)與MDA相似,25 ℃條件下,Pro含量比較穩(wěn)定,無明顯變化(圖4)。4 ℃條件下,Pro含量呈緩慢上升趨勢(shì),并于72 h時(shí)達(dá)到最大值,恢復(fù)期間略有下降,在48~72 h期間Pro含量上升速度最快。-4 ℃條件下,Pro含量上升速度較快,恢復(fù)期間仍在上升,并于96 h時(shí)達(dá)到最大值,其中0~6 h期間Pro含量上升速度最快。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)上升的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到130.85%和65.37%;至96 h時(shí),分別達(dá)到了142.12%和65.74%(表1)。
對(duì)于可溶性糖含量,25 ℃條件下,其值比較穩(wěn)定,無明顯變化(圖4)。4 ℃和-4 ℃條件下,可溶性糖含量在整個(gè)試驗(yàn)期間均呈持續(xù)上升趨勢(shì),恢復(fù)期間仍在上升,并于96 h時(shí)達(dá)到最大值,其中0~6 h期間可溶性糖含量上升速度最快。與4 ℃相比,-4 ℃處理隨著脅迫時(shí)間的延長(zhǎng)上升的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到77.94%和39.63%;至96 h時(shí),分別達(dá)到72.06%和49.02%(表1)。
2.5 抗氧化酶活性
溫度、處理時(shí)間以及兩者的交互作用顯著影響了油茶葉片的超氧化物歧化酶活性(SOD,p<0.05;圖5)、過氧化物酶活性(POD,p<0.05)和過氧化氫酶活性(CAT,p<0.05)。
對(duì)于SOD和POD活性,兩者的變化趨勢(shì)相似,25 ℃條件下,其活性均比較穩(wěn)定,變化幅度不大(圖5)。4 ℃和-4 ℃條件下,其活性均呈持續(xù)上升趨勢(shì),并于72 h時(shí)達(dá)到最大值,恢復(fù)期間略有下降,在0~6 h期間兩者活性上升速度最快。與4 ℃相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)上升的幅度更大。至72 h時(shí),-4 ℃和4 ℃處理下SOD的低溫變化率分別達(dá)到15.74%和11.83%,POD的低溫變化率分別達(dá)到95.97%和57.06%;至96 h時(shí),-4 ℃和4 ℃處理下SOD的低溫變化率分別達(dá)到7.97%和4.30%,POD的低溫變化率分別達(dá)到53.63%和19.15%(表1)。
對(duì)于CAT活性,25 ℃條件下,CAT活性比較穩(wěn)定,無明顯變化(圖5)。4 ℃和-4 ℃條件下,CAT活性均呈先上升后下降的趨勢(shì),在48 h時(shí)達(dá)到最大值,隨后下降,恢復(fù)期間仍持續(xù)降低,并于96 h時(shí)達(dá)到最低值。其中,4 ℃條件下在24~48 h期間CAT活性上升速度最快,-4 ℃條件下0~6 h期間上升速度最快。與4 ℃處理相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)上升的幅度更大。至72h時(shí),-4 ℃和4 ℃處理的低溫變化率分別達(dá)到31.30%和17.07%;至96 h時(shí),分別達(dá)到12.60%和3.54%(表1)。
3 結(jié)論與討論
3.1 結(jié)論
本試驗(yàn)研究表明,低溫脅迫顯著影響了長(zhǎng)林4號(hào)油茶的形態(tài)特征,使嫩莖和葉片出現(xiàn)不同程度的萎蔫和失水癥狀,并且-4 ℃處理比4 ℃處理植株受損程度更嚴(yán)重。對(duì)于葉片各氣體交換參數(shù),低溫脅迫使Pn、Gs、Tr值均有不同程度的降低,恢復(fù)期間,Pn、Gs、Tr值仍持續(xù)下降,且-4 ℃處理的抑制作用顯著強(qiáng)于4 ℃處理。RWUEi在4 ℃低溫處理的前48 h明顯高于對(duì)照,-4 ℃處理的前6 h略高于對(duì)照,之后均低于對(duì)照,恢復(fù)期間,RWUEi均略有下降。對(duì)于葉綠素含量,低溫脅迫雖然降低了油茶葉片的SPAD值,但未達(dá)到顯著水平。對(duì)于MDA、Pro和可溶性糖含量,低溫脅迫顯著提高了其含量,恢復(fù)期間,4 ℃處理的MDA和Pro含量略有下降,而可溶性糖含量仍繼續(xù)增加;-4 ℃處理的MDA和可溶性糖含量略有下降,Pro含量仍繼續(xù)增加。對(duì)于抗氧化酶活性,低溫脅迫顯著提高了油茶葉片中的抗氧化酶活性,隨著脅迫時(shí)間的延長(zhǎng),SOD和POD活性持續(xù)上升,而CAT活性則先上升后下降,并且-4 ℃處理的上升幅度明顯高于4 ℃處理;恢復(fù)期間,抗氧化酶活性均有不同程度的降低。
3.2 討論
3.2.1 低溫脅迫對(duì)油茶幼苗形態(tài)及光合性能的影響
表型指標(biāo)是顯示植物在低溫脅迫下受害程度最直接的指標(biāo)。本研究中,4 ℃處理的植株僅有嫩葉與嫩莖出現(xiàn)不同程度的表型損傷,而-4 ℃處理的植株其嫩葉、嫩莖和成熟葉片均出現(xiàn)了明顯的表型損傷,說明-4 ℃低溫對(duì)2年生油茶幼苗的傷害更大,且處理72 h便出現(xiàn)不可逆的傷害,這與王永紅等[11]采用油茶離體葉片在低溫脅迫下的研究結(jié)果一致。光合作用是決定植物發(fā)育的主要因素,往往會(huì)受到各種環(huán)境因子的影響,其中溫度是一個(gè)較為重要的影響因子。低溫對(duì)光合作用的影響體現(xiàn)在光合器官的結(jié)構(gòu)和活性、葉片氣孔導(dǎo)度、參與光合作用的酶活性等方面[12]。葉片氣體交換參數(shù)、葉綠素含量均為顯示逆境脅迫下植物光合性能的重要指標(biāo)[13]。低溫脅迫下,相關(guān)指標(biāo)的變化率常常可以反映植物的受害程度,變化率越大,說明植物受害越嚴(yán)重[14]。本試驗(yàn)中,低溫脅迫導(dǎo)致長(zhǎng)林4號(hào)油茶葉片的Pn、Gs、Tr、RWUEi等氣體交換參數(shù)值均有不同程度的降低。與4 ℃處理相比,-4 ℃處理下降的幅度更大;恢復(fù)期間,這些參數(shù)值均未升高,甚至仍持續(xù)降低,這一結(jié)果與王雨水等[15]對(duì)‘雜優(yōu)閩1油茶的研究結(jié)果相似,說明4 ℃和-4 ℃低溫對(duì)油茶幼苗有較大的傷害,且溫度越低傷害越大,并難以在短時(shí)間內(nèi)恢復(fù),與植株的表型結(jié)果基本一致。試驗(yàn)中發(fā)現(xiàn),低溫脅迫下葉片的SPAD值與對(duì)照相比雖然略有降低,但是差異不顯著,這表明低溫造成光合性能降低的主要因素是生理代謝或者氣孔因素,而不是葉綠素含量的降低。
3.2.2 低溫脅迫對(duì)油茶幼苗丙二醛、脯氨酸、可溶性糖含量的影響
植物在低溫脅迫的環(huán)境下,細(xì)胞膜會(huì)發(fā)生膜脂過氧化反應(yīng),膜脂過氧化物分解產(chǎn)生丙二醛,其含量高低與植物抗逆性強(qiáng)弱呈負(fù)相關(guān)關(guān)系[16],脯氨酸和可溶性糖是重要的滲透調(diào)節(jié)物質(zhì),能夠保持細(xì)胞內(nèi)環(huán)境的穩(wěn)定,維持細(xì)胞代謝平衡,降低植物受害程度[17]。因此,丙二醛、脯氨酸和可溶性糖含量是顯示植物抗寒性重要指標(biāo)。本試驗(yàn)中,這些指標(biāo)值均有不同程度的升高。在4 ℃條件下,這些指標(biāo)上升的幅度比較緩慢,在0~24 h期間增幅不大,至48 h時(shí)開始迅速增加,恢復(fù)期間略有下降,這與王國(guó)霞等[18]對(duì)‘大別山1號(hào)、‘豫油茶1號(hào)、‘豫油茶3號(hào)油茶的研究結(jié)果相似。說明在低溫處理前期植物受傷害程度較小,細(xì)胞液滲透率不大,細(xì)胞膜結(jié)構(gòu)完整,能夠維持正常的生理代謝活動(dòng)。隨著脅迫時(shí)間的延長(zhǎng),植物受傷害程度逐漸增大,細(xì)胞膜脂過氧化程度加劇,細(xì)胞內(nèi)環(huán)境失去平衡,不能維持正常的生理代謝。恢復(fù)期間,丙二醛和脯氨酸含量均有不同程度降低,說明油茶能夠耐受一定程度的低溫,且通過生理調(diào)控可以維持一定的時(shí)間。與4 ℃處理相比,-4 ℃處理下這些指標(biāo)在每個(gè)時(shí)間點(diǎn)上升的速度更快,這與文佳等[19]對(duì)‘大別山1號(hào)油茶的研究結(jié)果相似,說明油茶幼苗雖然能夠形成低溫適應(yīng)機(jī)制,表現(xiàn)出一定的抗寒性,但是超出其抗寒性所能抵御的范圍時(shí),就會(huì)導(dǎo)致油茶幼苗代謝功能嚴(yán)重失衡,膜脂過氧化程度加劇,細(xì)胞膜結(jié)構(gòu)嚴(yán)重受損。
3.2.3 低溫脅迫對(duì)油茶幼苗抗氧化酶活性的影響
一般來說,活性氧自由基在植物體內(nèi)都有分布,但是含量不高,處于產(chǎn)生和清除的平衡狀態(tài),對(duì)植物的新陳代謝不產(chǎn)生影響。當(dāng)植物受到低溫脅迫時(shí),細(xì)胞的防御能力會(huì)下降,植物體內(nèi)會(huì)產(chǎn)生大量的活性氧,造成活性氧自由基等物質(zhì)的積累。SOD和POD可以清除細(xì)胞內(nèi)的活性氧和自由基,降解植物代謝產(chǎn)生的H2O2,降低膜脂過氧化程度,保持細(xì)胞內(nèi)自由基的平衡,CAT能夠清除線粒體和膜脂過氧化產(chǎn)生的活性氧,提高植物對(duì)低溫脅迫的適應(yīng)性[19-21]。這3種抗氧化酶與植物的抗寒性密切相關(guān),已經(jīng)被大量研究所證明[22-24]。梁大偉[22]等研究了自然降溫對(duì)紅花玉蘭抗寒性生理指標(biāo)的影響,結(jié)果表明隨著溫度的降低,SOD、POD和CAT這3種酶活性逐漸增大,但達(dá)到一定脅迫程度后會(huì)逐漸下降。蔣景龍等[25]對(duì)漢中主栽柑橘品種在低溫脅迫下的生理響應(yīng)進(jìn)行了研究,結(jié)果表明SOD、POD和CAT這3種酶活性均表現(xiàn)出先升高后降低的趨勢(shì)。本研究表明,在25 ℃條件下,這3種酶活性均比較穩(wěn)定,變化幅度不大。4 ℃和-4 ℃條件下,這3種酶活性會(huì)出現(xiàn)不同程度的上升,其中SOD和POD活性呈持續(xù)上升趨勢(shì),CAT活性呈先上升后下降趨勢(shì),恢復(fù)期間,這3種酶活性均有不同程度的下降。并且與4 ℃處理相比,-4 ℃處理在每個(gè)時(shí)間點(diǎn)上升的幅度更大,這與已有的研究結(jié)果相似。說明低溫脅迫可以誘導(dǎo)抗氧化物酶活性的增加,減少細(xì)胞內(nèi)自由基和活性氧的積累,從而提高油茶幼苗的抗寒性,但是隨著處理溫度的降低,脅迫時(shí)間的延長(zhǎng),植株受傷害程度逐漸加深,其體內(nèi)抗氧化系統(tǒng)會(huì)遭到破壞,導(dǎo)致酶活性逐漸下降。
參 考 文 獻(xiàn)
[1]莊瑞林.中國(guó)油茶[M].北京:中國(guó)林業(yè)出版社,2008.
[2]石鵬皋,羅治建.湖北省油茶產(chǎn)業(yè)發(fā)展對(duì)策[J].湖北林業(yè)科技,2009(01):48-51.
[3]王斌,王開良,童杰潔,等.我國(guó)油茶產(chǎn)業(yè)現(xiàn)狀及發(fā)展對(duì)策[J].林業(yè)科技開發(fā),2011,25(02):11-15.
[4]肖玉霞.我國(guó)油茶產(chǎn)業(yè)發(fā)展現(xiàn)狀及其高效發(fā)展措施探討[J].南方農(nóng)業(yè),2019,13(29):109-110.
[5]石林曌,倪天虹,裴云霞,等.湖北省主栽油茶品種苗期抗寒性評(píng)價(jià)[J].經(jīng)濟(jì)林研究,2019,37(04):10-19.
[6]侯陽(yáng).油茶種植優(yōu)勢(shì)及豐產(chǎn)栽培技術(shù)探討[J].南方農(nóng)業(yè),2019,13(29):5-6.
[7]張琳,譚曉風(fēng),胡姣,等.油茶乙酰CoA酰基轉(zhuǎn)移酶基因cDNA克隆及序列特征分析[J].中南林業(yè)科技大學(xué)學(xué)報(bào),2011,31(08):108-112.
[8]張日清,王承南,李建安,等.關(guān)于油茶現(xiàn)代產(chǎn)業(yè)化體系建設(shè)的戰(zhàn)略思考[J].經(jīng)濟(jì)林研究,2010,28(02):146-150.
[9]Mielke MS,Dealmeida AAF,Gomes FP,et al.Effects of soil flooding on leaf gas exchange and growth of two neotropical pioneer tree species.New Forest,2005,29(02): 161-168.
[10]王學(xué)奎,黃見良.植物生理生化實(shí)驗(yàn)原理與技術(shù)[M].北京:高等教育出版社,2015.
[11]王永紅.山茶屬主要物種及部分園藝品種耐寒性研究[D].北京:中國(guó)林業(yè)科學(xué)研究院,2005.
[12]Lambers HL,Chapin FS,Pons TL.Plant Physiological Ecology[M].New York: Springer New York,2008.
[13]Zooaoui R,Ammari Y,Abassi M,et al.Physiological and biochemicall responses of Rhus tripartite (ucria) grande under water stress[J].Pakistan Journal of Botany,2019,51(4): 1215-1221.
[14]Du KB,Xu L,Wu H,et al.Eco-physiological and morphological adaption to soil flooding of two poplar clones differing in flood-tolerance[J].Flora,2012,207(2): 96-106.
[15]王雨水.低溫鍛煉對(duì)冷脅迫下油茶幼苗光合速率與抗氧化酶活性的影響[J].福建林業(yè)科技,2011,38(1):41-46.
[16]李曉靖,崔海軍.低溫脅迫下植物光合生理研究進(jìn)展[J].山東林業(yè)科技,2018,48(06):90-94.
[17]任軍,黃志霖,曾立雄,等.低溫脅迫下植物生理反應(yīng)機(jī)理研究進(jìn)展[J].世界林業(yè)研究,2013,26(06):15-20.
[18]王國(guó)霞,關(guān)欣,楊玉珍,等.變溫低溫脅迫對(duì)油茶生理生化特性的影響[J].貴州農(nóng)業(yè)科學(xué),2017,45(9):12-16.
[19]文佳,張誠(chéng)誠(chéng),曹志華,等.低溫脅迫對(duì)油茶苗抗寒性的影響[J].湖北農(nóng)業(yè)科學(xué),2014,53(03):609-612.
[20]林鄭和,鐘秋生,游小妹,等.低溫脅迫對(duì)茶樹抗氧化酶活性的影響[J].茶葉科學(xué),2018,38(04):363-371.
[21]洪文泓,董斌,黃永芳,等.三種油茶幼苗對(duì)低溫脅迫的生理響應(yīng)及其抗寒性綜合評(píng)價(jià)[J].亞熱帶植物科學(xué),2016,45(02):117-121.
[22]梁大偉,馬履一,賈忠奎,等.自然降溫對(duì)紅花玉蘭抗寒生理指標(biāo)的影響[J].林業(yè)科技開發(fā),2010,24(2):23-26.
[23]張婷,車?guó)P斌,潘儼,等.低溫脅迫對(duì)核桃枝條幾個(gè)抗寒生理指標(biāo)的影響[J].新疆農(nóng)業(yè)科學(xué),2011,48(8):1428-1433.
[24]逯明輝,宋慧,李曉明,等.冷害過程中黃瓜葉片SOD、CAT和POD活性的變化[J].西北植物學(xué)報(bào),2005,25(8):1570-1573.
[25]蔣景龍.漢中主栽柑橘品種對(duì)低溫脅迫的生理響應(yīng)[J].河南農(nóng)業(yè)科學(xué),2016,45(3):106-111,115.
(責(zé)任編輯:唐 嵐)
