生物修復劑TF3對鉛污染土壤的修復效果研究
2020-04-09 01:22:08郜雅靜李建華盧晉晶靳東升郜春花
生態科學 2020年2期
郜雅靜, 李建華, 盧晉晶, 靳東升, 郜春花*
生物修復劑TF3對鉛污染土壤的修復效果研究
郜雅靜1, 李建華2, 盧晉晶2, 靳東升2, 郜春花2*
1. 山西大學生物工程學院, 太原 030006 2. 山西省農業科學院農業環境與資源研究所, 太原 030031
為研究生物修復劑TF3(0.5 g·kg-1腸桿菌干菌體+20 g·kg-1生物炭+20 g·kg-1有機肥)對鉛污染土壤的修復效果, 選取5種鉛濃度土壤為研究對象, 采用盆栽實驗方法, 研究施用修復劑TF3后, 鉛在油菜體內的富集、轉運, 以及土壤有效態鉛、酶活性、微生物的響應。結果表明: TF3能夠促進油菜生長、顯著降低油菜地上部、地下部鉛含量。與CK組相比, TF3組油菜地上部、地下部及總富集系數分別降低20.12%—71.91%、 37.75%—60.13%、22.45%—68.77%。TF3組顯著降低土壤有效態鉛含量, 降低23.10%—39.84%。添加TF3, 土壤四種酶活性和微生物數量都有不同程度的增加, 真菌、放線菌、細菌分別較CK組增加11.76%—40.00%、6.45%—25.61%、120.20%—290.24%。研究表明, TF3降低了污染土壤鉛對油菜的脅迫作用, 可以作為修復鉛污染的一種有效生物修復劑。
生物修復劑; 鉛含量; 污染土壤; 油菜; 富集系數
0 前言
鉛作為農業環境中主要重金屬污染元素之一, 主要通過工業廢物排放、汽車廢氣沉降、生活垃圾隨意堆放等途徑, 沉積到土壤中, 不易被降解, 最終隨著土壤-植物-動物逐級累積, 最終進入人體[1], 對人類的健康和生存構成威脅。據有關統計, 全世界每年鉛用量大約有4×106t, 但只有1/4的鉛實現了回收再利用, 剩余大部分造成了環境污染[2]。鉛在重金屬污染物中, 屬于毒性最強之一, 含量最高的一類[3]。因此, 治理鉛污染土壤迫在眉睫。
目前, 重金屬污染土壤修復常用的處理方法多為物理、化學方法, 這些方法都會不同程度地產生二次污染, 而且處理成本相對較高, 過程操作復雜[4]。而生物修復具有費用低、效果好等優勢, 因此具有良好的發展前景[5]。目前針對重金屬生物修復主要以單接微生物為主, 而直接施加微生物會導致微生物的流失或吞噬, 而且微生物生長代謝能力不佳, 對重金屬的固定能力有限[6]。因此本文通過前期實驗研發了一種生物修復劑TF3由0.5 g·kg-1腸桿菌干菌體、20 g·kg-1生物炭和20 g·kg-1有機肥組成。腸桿菌通過在鉛污染土壤篩選而得, 具有高耐鉛性且在水溶液中對Pb2+吸附率可以達到98%以上[7]。生物炭具有孔隙結構較大、表面活性大、吸附能力很強等優勢, 另外它還具有性質及穩定的碳結構, 能夠增加有機質含量、改良土壤性質[8], 使其在固定重金屬方面有很大的潛力。有機肥能夠提供土壤養分、疏松土壤、促進土壤養分的轉換, 改善土壤微環境。因此, 該實驗通過在不同濃度鉛污染土壤上種植油菜, 來驗證TF3對鉛污染土壤的修復效果。
1 材料與方法
1.1 供試材料
實驗于2018年5月—7月在山西省農業科學院溫室內進行。試驗所在地海拔790 m, 屬于大陸性暖溫帶半干旱季風氣候, 無霜期平均149—175 d, 年均降水量為468.4 mm。
供試修復劑TF3: 0.5 g·kg-1腸桿菌干菌體+ 20 g·kg-1生物炭+20 g·kg-1有機肥(TF3: 土壤=1:25)。
菌株: 自主篩選的菌株腸桿菌屬() GDYX03, 形態呈擴展形, 表面光滑凸起, 易挑起, 乳白色且半透明, 為革蘭氏陰性菌。
市售的生物質炭、商品有機肥基礎性質見表1、表2。
供試植物為青美油菜(L), 購買于山西瑞豐種業。
供試污染土壤制備: 添加不同濃度的外源鉛溶液, 用Pb(NO3)2分析純試劑配置, 加入去離子水, 使所有處理的土壤水分達到田間持水量的70%, 充分混勻裝入塑料框中, 干濕交替, 老化10個月, 獲得一系列鉛濃度的實驗土壤[9], 具體性質見表3。

表1 生物炭理化性質

表2 有機肥的養分含量
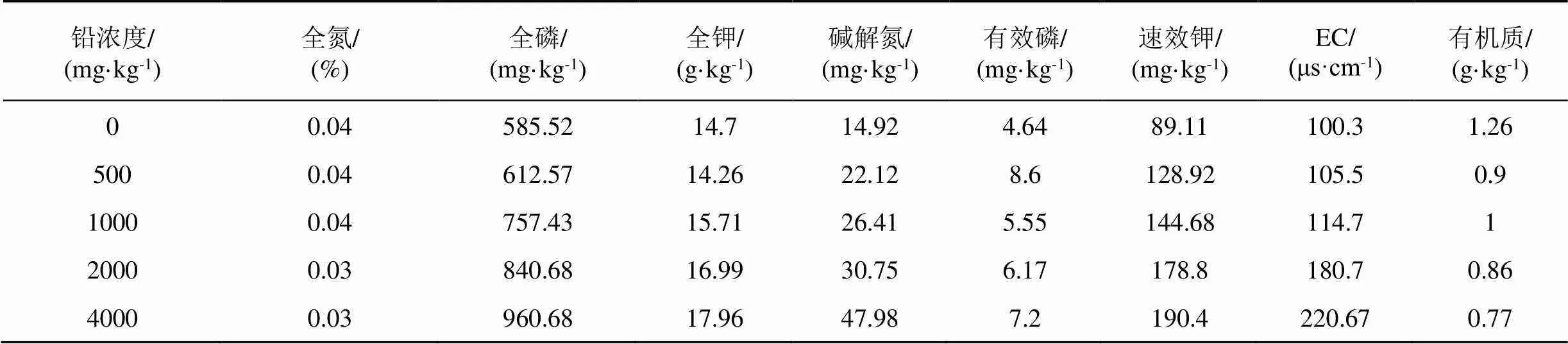
表3 土壤基礎性質
1.2 實驗設計
選取5個鉛濃度(0、500、1000、2000、4000mg·kg-1)進行修復實驗, 采用裂區設計, 一組中不添加修復劑為CK組, 另一組中添加修復劑為TF3組, 種植油菜, 生長時間55 d(成熟期), 每個處理4次重復。所有處理基施無機肥20 g (N:P2O5:K2O為4:3:3), 與土壤混勻后裝盆, 每盆裝土量為2.5 kg, 每盆從育苗盤挑選長勢均勻的3株移入, 以稱重法保持土壤含水量在田間持水量的75%左右。
1.3 樣品采集
植株樣品的采集: 收獲時, 將植株完整的從盆中拿小鏟子輕輕挖出, 分離地上、地下部, 分別先用自來水沖洗干凈泥土, 再用去離子水沖洗干凈, 105 ℃殺青30 min, 然后65 ℃烘干, 粉碎、裝入自封袋中儲存, 用于測定植株鉛含量。
土壤樣品的采集: 挖起植株, 采用抖落法收集根區土壤, 用滅菌的鑷子去除土壤中的雜質, 一部分裝入已編號的無菌自封袋, 置于4 ℃冰箱, 盡快進行微生物數量及多樣性的分析; 一部分風干、磨碎、過篩, 用于土壤中鉛含量的測定和土壤酶活性的分析。
1.4 實驗指標測定方法
(1)鉛含量的測定
植株中鉛含量: 加入硝酸, 高氯酸酸化, 使用微波消解儀消解, 采用火焰原子吸收分光光度計(240FSAA)測定[10], 單位: mg·kg-1。
土壤有效態鉛含量: 用DTPA(二乙三胺五乙酸)浸提劑(0.005 mol·L-1DTPA+0.1mol·L-1TEA(三乙醇胺)+0.01 mol·L-1CaCl2)浸提, 0.45 um微孔濾膜過濾, 采用火焰原子吸收分光光度計(240FSAA)測定[11], 單位: mg·kg-1。
土壤殘留鉛含量: 加入鹽酸-硝酸-氫氟酸-高氯酸酸化, 微波消解儀消解, 采用火焰原子吸收分光光度計(240FSAA)測定[11], 單位: mg·kg-1。
(2)土壤酶的測定
脲酶活性: 苯酚鈉-次氯酸鈉比色法; 磷酸酶活性: 磷酸苯二鈉法; 蔗糖酶活性: 3, 5-二硝基水楊酸比色法[12]。過氧化氫酶活性: 采用紫外分光光度法[13]。
(3)土壤微生物數量的測定
稀釋平板計數法: 細菌培養使用牛肉膏蛋白胨培養基, 放線菌培養使用改良高氏Ⅰ號培養基, 真菌培養使用孟加拉紅培養基[14]。
微生物多樣性采用Biolog Eco板法測定[15]。
1.5 數據統計
盆栽實驗數據用四個重復的平均數±標準偏差表示(X±SD,=4), 差異顯著性用 SPSS 18.0軟件進行Duncan 法檢驗, 分析在<0.05水平(表示差異達顯著水平)。Microsoft Excel 2007軟件作圖。
1.6 計算公式

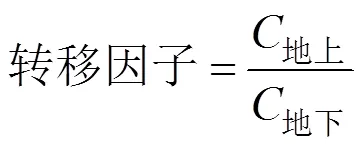
(3)
(4)

式中: 各處理值為TF3組各測量指標數值; CK處理值為對照組測量指標的具體數值; C地上為油菜地上部鉛含量, mg·kg-1;地下為油菜地下部鉛含量, mg·kg-1;土壤為土壤殘留鉛含量, mg·kg-1。
2 結果與分析
2.1 TF3對污染土壤中油菜生長的影響
2.1.1 TF3對污染土壤中油菜生長指標的影響
修復劑TF3對油菜生長的影響見表4, 在鉛濃度為0—4000 mg·kg-1范圍內添加TF3, 油菜葉片數、株高均顯著增加, 葉綠素在鉛濃度為4000 mg·kg-1, 增加幅度最大, 增率達5.78%。在鉛濃度為2000 mg·kg-1時, 油菜葉片數、株高增率最大, 較CK分別增加39.77%、34.54%、85.27%。
隨土壤中鉛濃度的增加, 添加TF3和CK兩組中, 葉片數、株高均表現為先增加后降低, 在鉛濃度為500 mg·kg-1時, 達到最大值; 而油菜葉綠素隨著鉛濃度的增加, 一直減小, 說明了鉛濃度的增大, 降低了葉綠素合成作用, 分解作用增強, 降低了葉綠素含量。
2.1.2 TF3對污染土壤中油菜生物量的影響
由表5可知, 隨土壤中鉛濃度的增加, TF3和CK兩組中, 油菜生物量表現為先增加后降低趨勢, 在鉛濃度為500 mg·kg-1時, 達到最大量。與CK組相比, 土壤中添加TF3后, 油菜生物量均顯著增加(<0.05)。在鉛濃度為2 000 mg·kg-1時, 地上部鮮重、地下部干重增率最大, 分別增加120.36%、66.96%; 地下部鮮重在鉛濃度4000 mg·kg-1時, 增率最大為60.06%; 地上部干重在鉛濃度1000 mg·kg-1時增率最大, 較CK增加81.99%。
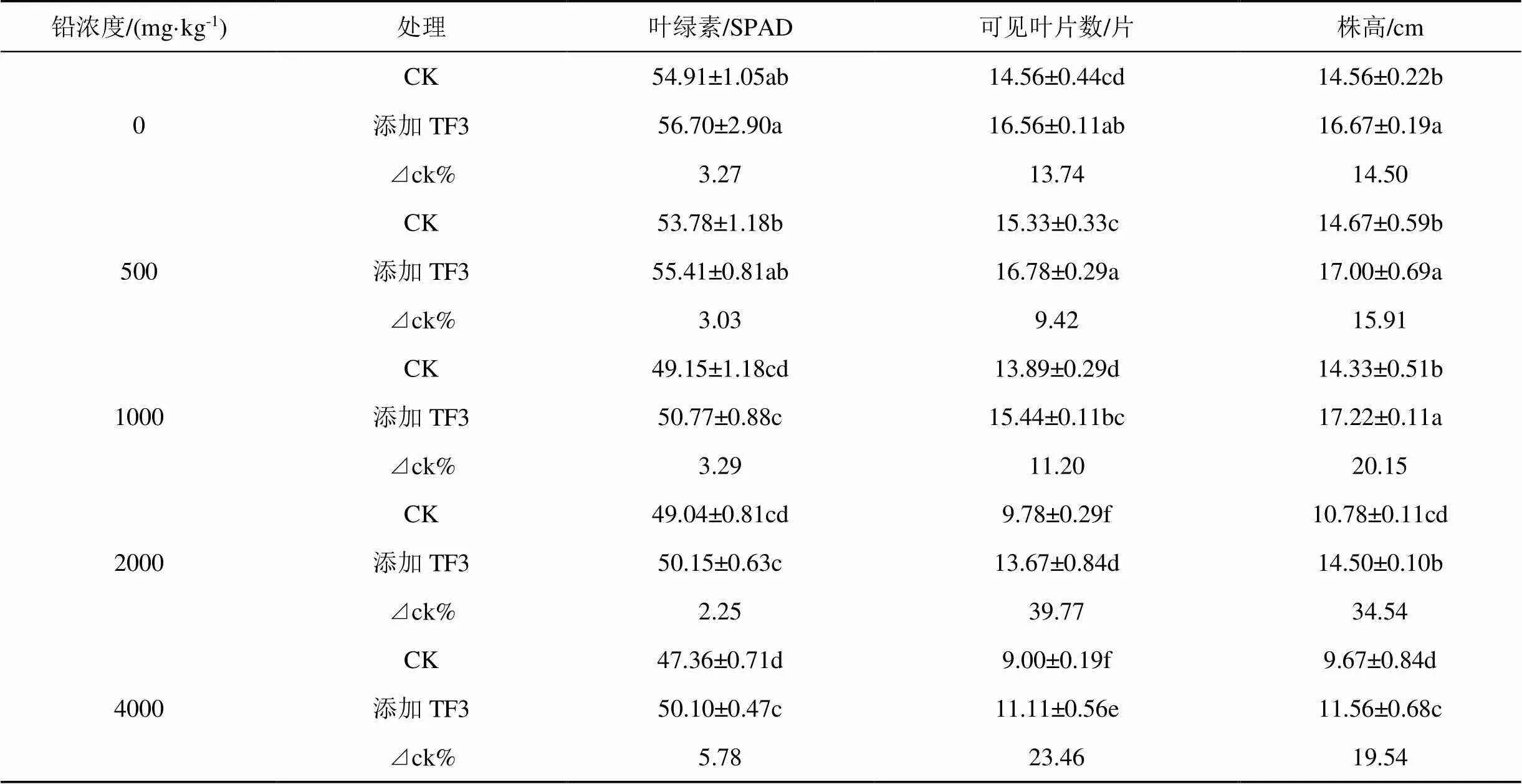
表4 TF3對污染土壤中油菜生長指標的影響
注:表中數據形式為: 平均值±標準差, 重復數=4; 同列不同的小寫字母表示同指標之間有顯著性差異(<0.05), 相同的小寫字母表示同指標間差異不顯著,下表相同。
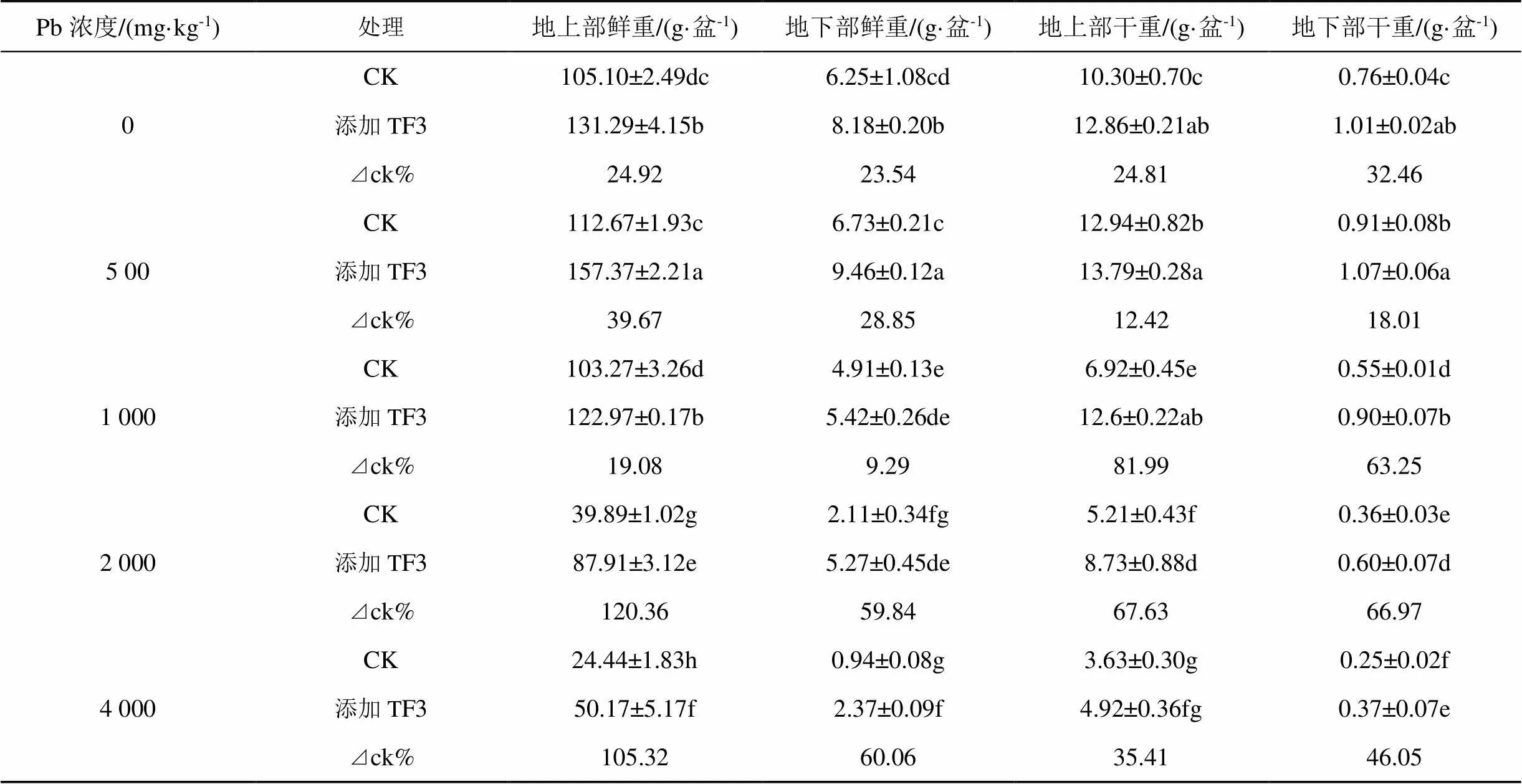
表5 TF3對污染土壤油菜生物量的影響
2.2 TF3對污染土壤油菜地上部、地下部鉛含量的影響
2.2.1 TF3對污染土壤中油菜地上部鉛含量的影響
由圖1可知, 在污染土壤中CK組中油菜地上部鉛含量為0.44—129.99 mg·kg-1, TF3組中油菜地上部鉛含量為0.22—90.63 mg·kg-1, 添加TF3后使油菜地上部的鉛含量顯著降低30%—50%。在土壤鉛濃度為 0、500、1000、2000、4000 mg·kg-1時, 地上部鉛含量分別降低50.01%、47.62%、42.36%、41.28%、30.28%。添加TF3后均降低了油菜地上部的鉛含量。
鉆戒若在許沁手上,許沁大可不必驚慌,大不了退了鉆戒——可鉆戒不在她手里;如果鉆戒真的寄國外朋友了,她也不必驚慌,讓朋友再寄回來也不很難——事實上她并沒有寄國外朋友;她的確把鉆戒送人了,且送的人非親非友,而是個當官的——給當官送禮,一般皆有求于官,焉能再索討回來呢?東西要回來了,求人的事就泡湯了。
2.2.2 TF3對污染土壤油菜地下部鉛含量的影響
由圖2可知, 在污染土壤中, 油菜地下部鉛含量遠遠大于地上部鉛含量, 油菜地下部鉛含量與地上部鉛含量規律一致, 均隨著土壤鉛濃度的增加, 油菜地下部鉛含量逐漸增加。CK組中的油菜地下部鉛含量為1.22—851.36 mg·kg-1, 添加TF3組油菜地下部鉛含量為0.45—763.01 mg·kg-1, TF3的施用顯著降低了油菜地下部鉛含量, 使油菜地下部的鉛含量降低10%—30%。在鉛濃度為 0、500、1000、2000、4000 mg·kg-1時, 地下部鉛含量分別降低50.01%、47.62%、42.36%、41.28%、30.28%。
2.3 TF3對油菜中鉛轉移因子、富集系數的影響
TF3對油菜中鉛轉移因子和富集系數的影響見表6。TF3對油菜中鉛的轉移因子影響不明顯, 在土壤鉛濃度大于500 mg·kg-1, 添加TF3后, 油菜中鉛的轉移因子減小, 阻礙了油菜將鉛從地下部向地上部分轉移; 在土壤鉛濃度小于500 mg·kg-1, 添加TF3后油菜中鉛轉移因子增大。
在土壤鉛濃度0—4000 mg·kg-1范圍內加入TF3后顯著降低了油菜中鉛地上部、地下部、總富集系數。地下部及總富集系數在鉛濃度1000 mg·kg-1最大, 在鉛濃度1000 mg·kg-1時, CK組地下部富集系數、總富集系數分別為0.397、0.454, TF3組分別為0.250、0.278, TF3組顯著降低37.75%— 60.13%、22.45%—68.77%。地上部富集系數在鉛濃度500 mg·kg-1時達到最大, CK處理為0.063、添加TF3處理為0.028, TF3組顯著降低20.12%— 71.91%。
2.4 TF3對污染土壤鉛含量的影響
2.4.1 TF3對污染土壤有效態鉛含量的影響
從圖3可以看出, 添加TF3后均能夠降低土壤有效態鉛含量, 在鉛濃度大于500 mg·kg-1達到顯著降低(<0.05)。在土壤鉛濃度0—4000 mg·kg-1范圍內添加TF3, 有效態鉛含量較CK組降低幅度均達到20%以上, 且隨著鉛濃度的增大, 降低的幅度逐漸減小。在鉛濃度0—4000 mg·kg-1范圍內, CK組有效態鉛含量為1.68—2705.21 mg·kg-1; TF3組有效態鉛含量為1.02—2080.41 mg·kg-1, TF3組較CK組有效態鉛含量分別降低39.84%、28.31%、27.21%、26.56%、23.10%。
2.4.2 TF3對污染土壤殘留鉛含量的影響
由圖4可以看出, 在不同鉛濃度中添加TF3土壤殘留鉛含量均有所增加。在土壤鉛濃度0—4000 mg·kg-1范圍內, CK組殘留鉛含量為20.33—3470.03 mg·kg-1; TF3組殘留鉛含量為26.4—3889.83 mg·kg-1。在5個土壤濃度下, TF3組較CK組殘留鉛含量分別增加6.07、122.50、151.00、226.83、419.80 mg·kg-1, 增加幅度分別達到29.84%、18.28%、15.15%、12.15%、12.10%。隨著土壤鉛濃度的增大, TF3對土壤中殘留鉛增加幅度在逐漸減小。通過添加TF3對土壤重金屬的固定增加, 導致土壤有效態鉛含量降低, 土壤殘留鉛含量增加。
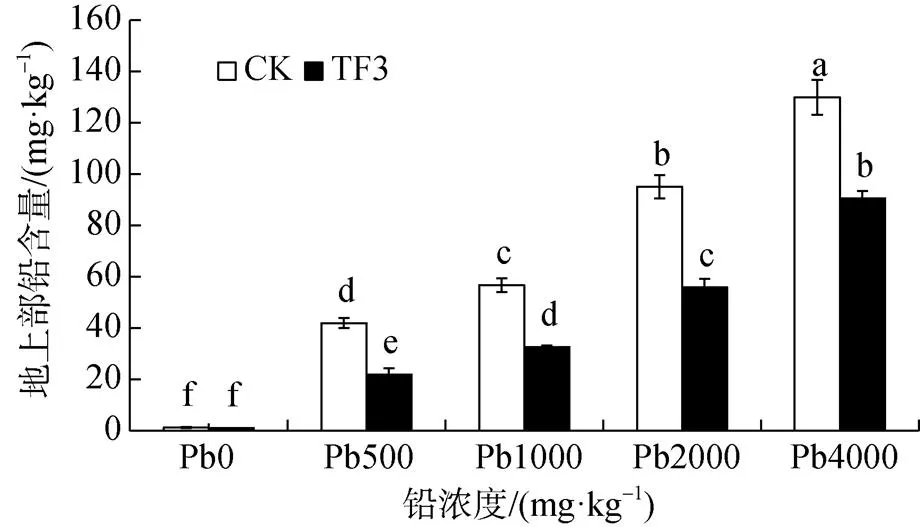
圖1 TF3對污染土壤油菜地上部鉛含量的影響
Figure 1 Effect of TF3 on lead content in the above ground part of rapeseed in contaminated soil
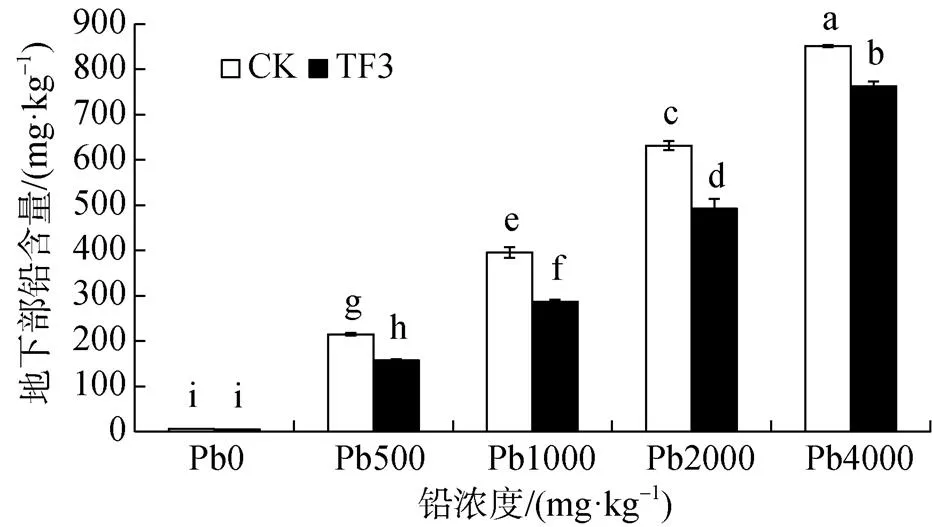
圖2 TF3對污染土壤油菜地下部鉛含量的影響
Figure 2 Effect of TF3 on lead content in the under part of rapeseed in contaminated soil
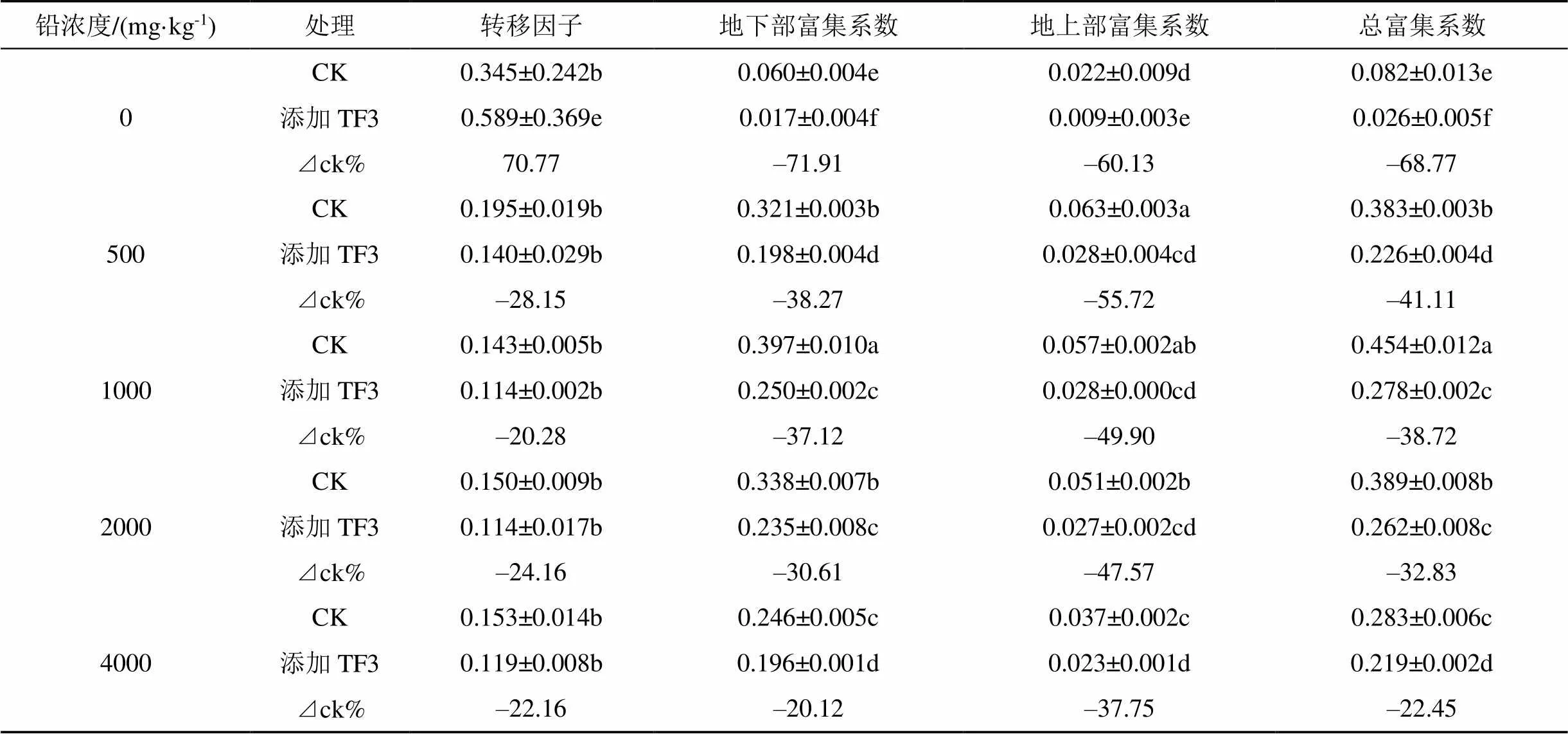
表6 TF3對油菜中鉛轉移因子和富集系數的影響
2.5 TF3對污染土壤酶活性的影響
由圖5可知, 鉛可刺激油菜根區土壤過氧化氫酶活性, 隨著鉛濃度增加土壤過氧化氫酶活性增加。添加TF3后土壤氧化氫酶活性增加, 增率為3.54%—7.30%。在鉛濃度0、500、1000、2000 mg·kg-1, 添加TF3過氧化氫酶活性增加達到顯著性差異(<0.05)。在鉛濃度4000 mg·kg-1, 添加TF3增加沒有達到顯著性差異。
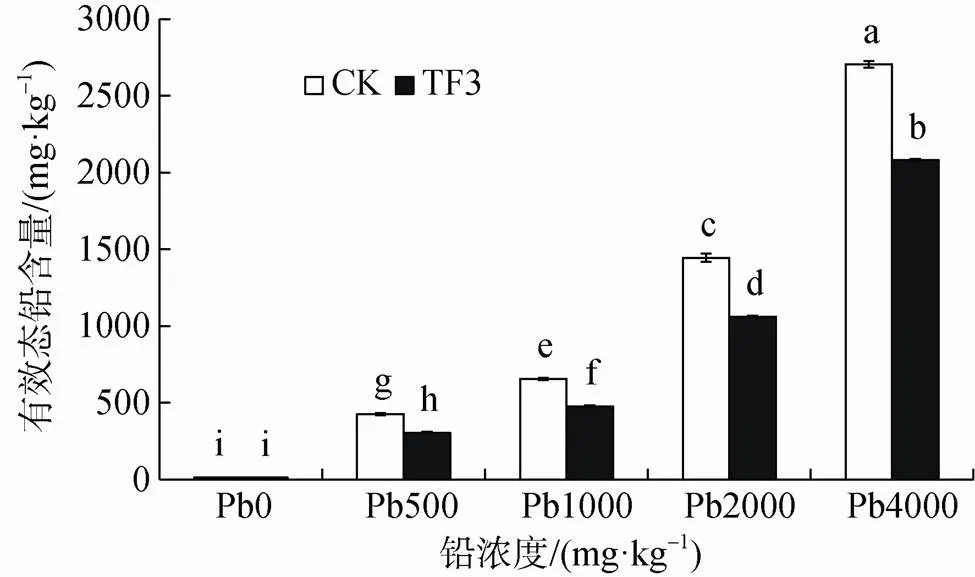
圖3 TF3對污染土壤有效態鉛含量的影響
Figure 3 Effect of TF3 on available lead content in contaminated soil
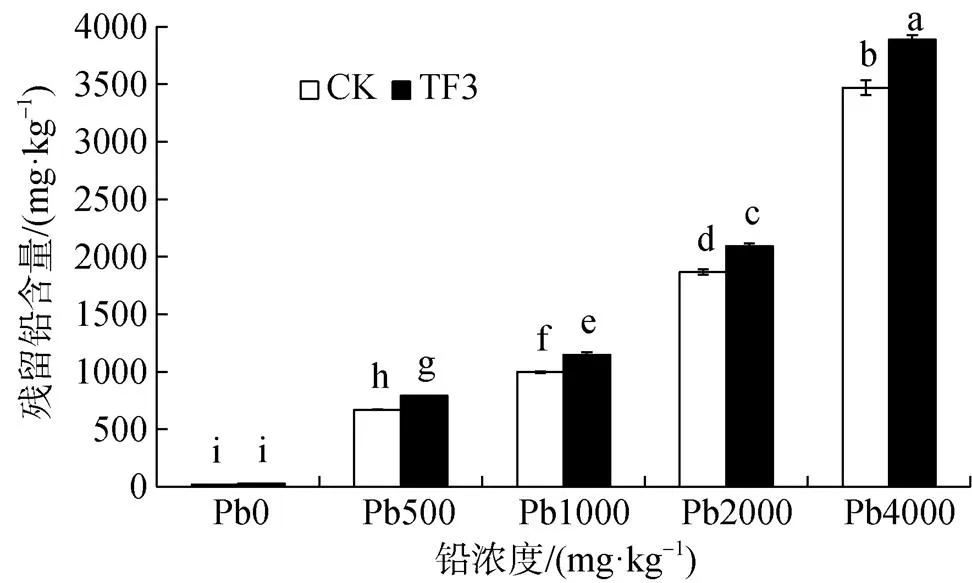
圖4 TF3對污染土壤殘留鉛含量的影響
Figure 4 Effect of TF3 on residual lead content in contaminated soil
由圖6可知, 在污染土壤中添加TF3后能夠增加土壤蔗糖酶活性, 其增率隨土壤鉛濃度增大逐漸減小。在鉛濃度為 0、500、1000、2000、4000 mg·kg-1時, 增率分別為23.98%、19.21%、15.38%、8.62%、7.49%。在鉛濃度為500、1000 mg·kg-1時, 增加達顯著性差異。說明添加TF3對低濃度鉛脅迫根區土壤蔗糖酶活性促進作用明顯。土壤蔗糖酶活性在鉛濃度為500 mg·kg-1時達到最強, 說明一定量的鉛濃度對土壤蔗糖酶活性有刺激作用, 在濃度超過2000 mg·kg-1對蔗糖酶有抑制作用。

圖5 TF3對污染土壤過氧化氫酶活性的影響
Figure 5 Effect of TF3 on catalase activity in contaminated soil
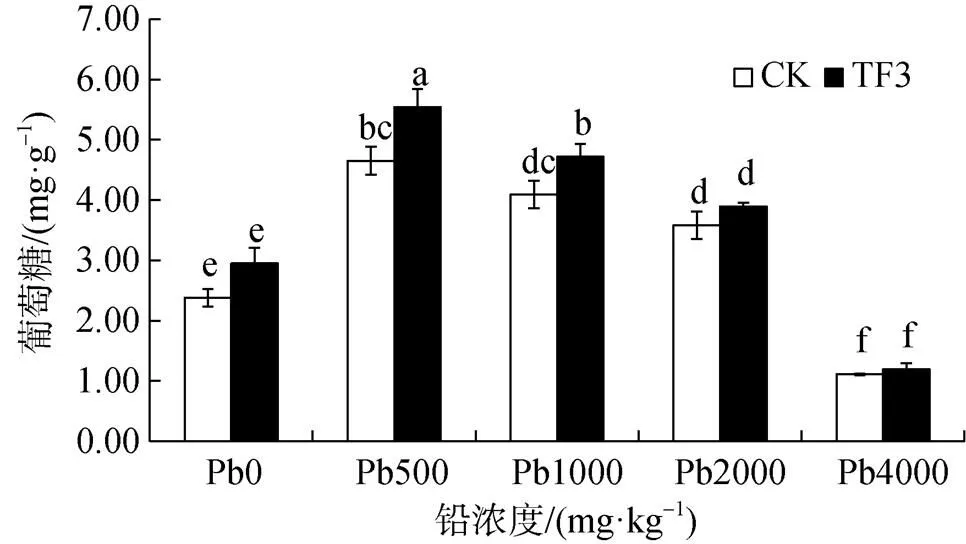
圖6 TF3對污染土壤蔗糖酶活性的影響
Figure 6 Effect of TF3 on the activity of invertase in contaminated soil
由圖7可知, 磷酸酶活性變化與蔗糖酶活性變化趨勢一致, 添加TF3后能夠增加油菜根區土壤中磷酸酶活性, 其增率為11.38%—21.90%, 在鉛濃度為500 mg·kg-1時增率最大, 在鉛濃度為4000 mg·kg-1時增加達顯著性差異。說明加入TF3鉛脅迫根區土壤磷酸酶活性促進作用不明顯。隨土壤鉛濃度增加磷酸酶活性先增加后降低, 在鉛濃度4000 mg·kg-1時活性最低, 在鉛濃度為500 mg·kg-1最高, 說明在一定鉛濃度下對土壤磷酸酶活性有一定的刺激作用。
由圖8可知, 添加TF3增加油菜根區土壤中脲酶活性, 其增率為20.25%—41.51%, 在鉛濃度為0、500、1000 mg·kg-1時增加達顯著性差異。說明添加TF3在小于鉛濃度1000 mg·kg-1對土壤磷酸酶活性起促進作用。脲酶活性同樣在鉛濃度為500 mg·kg-1最高, 在鉛濃度為4000 mg·kg-1最低, 說明一定含量的鉛對油菜根區土壤中脲酶活性有一定的刺激作用。
2.6 TF3對污染土壤微生物的影響
2.6.1 TF3對污染土壤微生物區系的影響
由表7可看出, 油菜根區土壤微生物數量: 細菌>放線菌>真菌。在污染土壤中添加TF3能夠增加土壤真菌數量, 在土壤鉛濃度0、2000 mg·kg-1增加顯著, 分別較CK增加40.00%、31.82%。在鉛濃度500、1000、4000 mg·kg-1增加不顯著, 分別較CK增加11.76%、11.48%、39.39%。隨土壤鉛濃度的增加, 土壤真菌在鉛濃度500 mg·kg-1時數量最多。
添加TF3后對土壤放線菌增加不顯著, 在鉛濃度0—4000 mg·kg-1范圍內增率為6.45%—25.61%, 在鉛濃度為4000 mg·kg-1增率最大。隨著鉛濃度的增加, 兩組處理油菜根區土壤放線菌數量變化同真菌變化一致, 先增加后減少, 在鉛濃度為500 mg·kg-1時數量最多。
在鉛污染土壤中添加TF3組均顯著增加土壤細菌數量, 且TF3組比CK組細菌數量增加達到1.2— 2.9倍。對土壤真菌、細菌、放線菌, 添加TF3后顯著增加根區土壤細菌的數量, 這可能與TF3組中加入的耐鉛菌有關。
2.6.2 TF3對污染土壤微生物多樣性的影響
多樣性指數會盡可能詳細的反應微生物群落物種組成和個體數量的分布情況, 反應微生物功能多樣性的不同側面[17]。由表8可看出, McIntosh指數越大, 說明群落均勻性越好。不同土壤鉛濃度下, TF3組McIntosh指數均比CK組大, 說明添加TF3后, 增加了土壤微生物群落的均勻性。Simpson指數越大, 說明群落多樣性越好[18]。TF3組較CK組, 隨著鉛濃度的增大Simpson指數分別增加2.37%、2.89%、5.81%、-1.36%、-1.74%。在鉛濃度大于2000 mg·kg-1時, 添加TF3, Simpson指數減小。Shannon均勻度指數越大, 說明群落越均一, 種類之間個體分布越均勻。添加TF3組, Shannon均勻度數值低于CK組, 說明相同鉛濃度, 添加TF3組降低了土壤微生物的均勻度, 使土壤微生物不均勻, 突出了TF3中耐鉛菌的作用。碳源利用豐富度指數S越大, 說明微生物可利用的碳源越多, 多樣性越好。添加TF3組, 碳源豐富度指數分別較CK組增加15.69%、10.93%、10.13%、1.55%、3.25%。隨著鉛含量的增加, 添加TF3后對碳源豐富度指數增加的幅度越來越小。
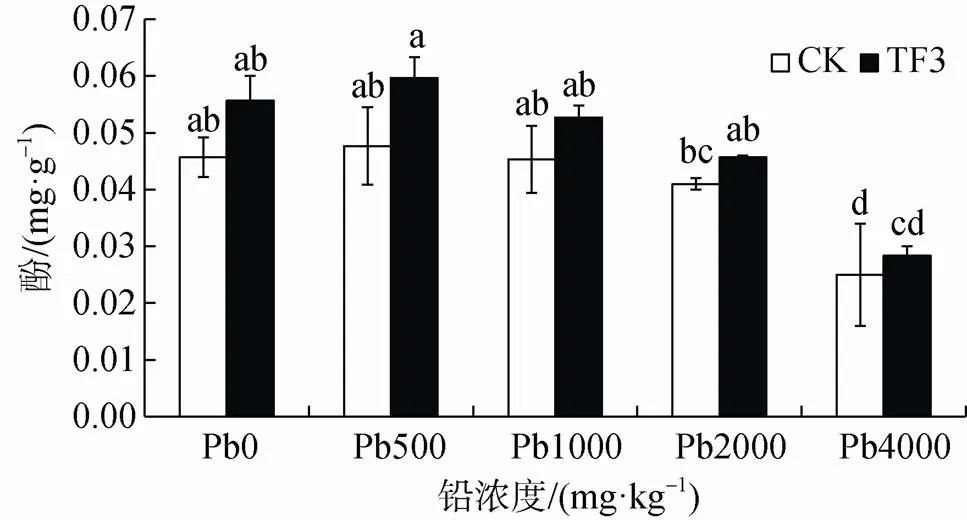
圖7 TF3對污染土壤磷酸酶活性的影響
Figure 7 Effect of TF3 on phosphatase activity in conta-minated soil
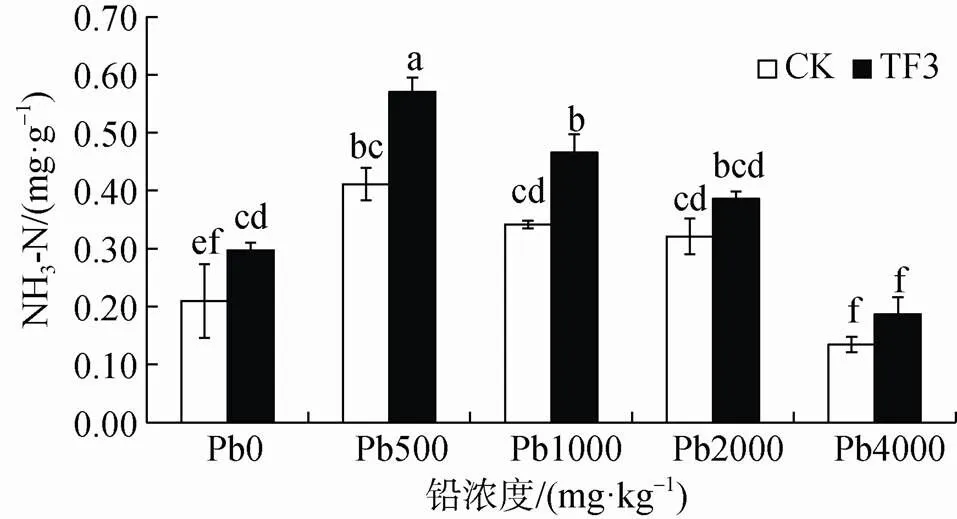
圖8 TF3對污染土壤脲酶活性的影響
Figure 8 Effect of TF3 on urease activity in contaminated soil
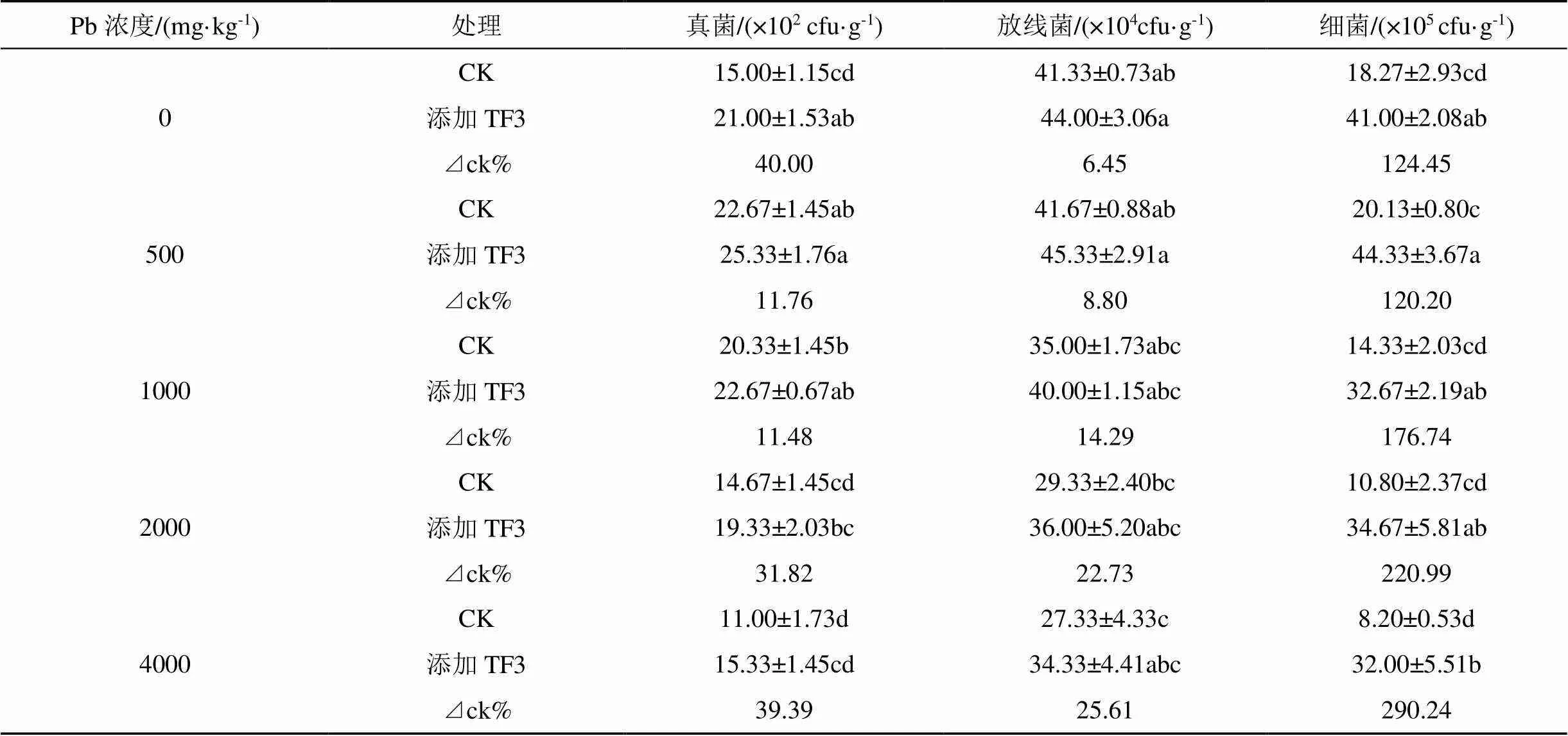
表7 TF3對污染土壤微生物區系的影響

表8 TF3對污染土壤微生物多樣性的影響
3 討論
種植植物是評估原位修復效果最有效的方法。通過添加修復劑來固定土壤重金屬含量的變化可以通過植物中重金屬含量的變化及植物生物量體現。大多數金屬離子通過與修復劑結合, 從而被固定在修復劑結構中, 修復劑極大地限制了重金屬離子在土壤中的遷移和富集。選擇合適的修復劑對鉛污染進行修復效果評估, 能更好地研究鉛離子在土壤-植物系統中的相互作用。
本實驗結果表明, 修復劑的施用能夠增加油菜生物量, 馮佳蓓也發現在鉛污染土壤中施加修復劑可以有效增加小白菜生物量; 但李紅等[19]通過在鉛污染土壤中加入蒙黏土與含磷材料作為修復劑, 發現其對小白菜生長無顯著影響。該實驗中油菜顯著增加, 可能是因為添加的TF3, 為油菜生長提供了一個相對良好的環境; 同時生物質炭、有機肥的添加能提高土壤碳、磷等營養元素含量, 促進作物對養分的吸收。而且, TF3能夠促進土壤對鉛的固定, 緩解對油菜的脅迫作用。
本研究表明, 加入修復劑TF3能顯著降低油菜地上部、地下部鉛含量。這可能是因為植物對金屬的吸收量主要取決于土壤有效態重金屬濃度。添加TF3后降低土壤中有效態鉛含量, 減少了植物對鉛吸附作用; 此外, 修復劑可以調節土壤微生態環境, 促進養分活化, 增加植物生物量。董同喜等[20]發現畜禽糞便可降低水稻土中重金屬的生物有效性, 進而降低水稻體內重金屬鉛含量; 曹書苗等[16]也發現加入生物肥可降低黑麥草土壤有效態鉛含量, 導致黑麥草地上和根部鉛含量降低; 這些研究與本研究有相似的結果。
研究發現, 鉛主要積累于油菜根系中, 油菜地下部分對鉛的富集能力遠高于地上部分, 導致地下部分鉛含量高于地上部分。說明鉛不易在植株體內轉運, 由轉運因子也可以看出。加入TF3增加了油菜生物量, 降低了油菜地上部、地下部的鉛含量, 這有利于控制植株體內的鉛殘留量[21]。同時TF3有效降低了油菜地下部對地上部的轉運能力, 轉運因子很低, 轉運能力弱。油菜對Pb的富集能力高于轉運能力, 尤其是地下部富集能力, 因此應該更關注Pb的富集作用。石汝杰等[22]發現鉛更易于被黑麥草地下部吸收, 不同處理地下部累積鉛含量均大于地上部, 這與本實驗研究結果一致。植株受鉛脅迫生長后, 隨著土壤中鉛濃度增加, 油菜地上部和地下部的鉛含量也增加, 土壤中鉛含量與植株內部鉛含量有一定的相關性。曹書苗等[16]研究黑麥草對鉛的耐受性和積累時也有相似的結果。
重金屬有效態可反映重金屬移動性、毒性和生物有效性, 受土壤重金屬總量、有機質含量等多種因素影響[23]。本文研究中可知, 添加TF3可顯著降低土壤有效態鉛含量。張敏[6]通過透射電鏡發現該菌株對 Pb2+的吸附主要是細胞表面的吸附, 也可以通過胞外某陰離子與Pb2+發生絡合反應形成沉淀。生物炭富含有較大的比表面積和大量官能團, 是重金屬離子的重要載體, 可以吸附大量重金屬元素[24]。有機肥在多數文獻[24-25]報道中表現促進作物生長, 具有增產作用, 被廣泛認為是影響土壤肥力的重要參數, 是增加作物產量、改善土壤性質的重要因子。通過耐鉛菌、生物炭與有機肥對土壤重金屬的固定增加, 導致土壤有效態鉛含量降低, 增加了土壤殘留鉛含量。鄭少玲等[26]通過土培試驗和盆栽試驗研究發現施用生物修復劑可以降低芥藍體內重金屬的含量, 增加生物量。Farfel 等人[27]發現在鉛污染土壤中添加污泥堆肥可提高草坪覆蓋面積, 降低土壤有效態鉛; 這些研究與本研究有相似的結果。
土壤酶活性一定程度上能夠反應土壤的養分含量和受污染程度。本研究中, 加入TF3對四種酶活性都表現為增加作用。可能是因為TF3對土壤重金屬含量影響較大, 有利于植物的生長, 增強了土壤酶活性。楊海征[28]通過在污染土壤中施加雞糞堆肥, 發現可以提高土壤脲酶、磷酸酶、蔗糖酶和過氧化氫酶的活性。楊繼飛[29]研究發現在鉛污染土壤上施用菌肥、腐殖酸, 能降低重金屬鉛的毒害, 提高玉米、高粱、蓖麻、向日葵含量, 增加土壤酶活性, 施用菌肥和腐殖酸可以達到修復鉛污染土壤的目的。
土壤微生物數量反應了土壤養分含量和肥力水平, 添加TF3能顯著提高鉛脅迫下油菜土壤微生物活性。這可能是因為一方面加入的耐鉛菌可以在土壤中大量繁殖, 促進微生物數量的增加; 一方面加入的有機肥含有豐富礦質元素和碳水化合物, 能夠疏松土壤、增加土壤孔隙度、提高土壤透氣性、改善土壤微環境, 為微生物生長提供一定的養分, 增強土壤生物活性[30]。牛旭[31]發現添加牛糞堆肥所制成的生物有機肥, 可提高種植春小麥的礦區復墾土壤微生物數量。張連忠等[32]也發現有機肥施用可提高果園根區土壤細菌、放線菌、真菌的數量, 明顯減輕重金屬對土壤微生物的危害, 改善重金屬污染土壤, 這與本研究有相似的結果。
4 結論
實驗結果表明, 在土壤鉛濃度為0—4000 mg·kg-1范圍內添加修復劑TF3, 均可以顯著促進油菜生長, 減少油菜體內鉛含量; 降低土壤有效態鉛含量23.10%以上、增加土壤酶活性和微生物多樣性。TF3降低了污染土壤鉛對油菜的脅迫作用, 可以作為修復鉛污染的一種有效生物修復劑。今后的研究中, 需要將生物修復劑與市售修復劑的效果進行比較, 為進一步應用TF3對重金屬污染土壤進行治理提供一定理論依據和實踐指導。
[1] 郜雅靜, 郜春花, 李建華, 等. 重金屬污染土壤的微生物修復技術探討[J]. 山西農業科學, 2018, 46(1): 150–154.
[2] 向捷, 陳永華, 向敏, 等. 土壤重金屬污染修復技術比較研究[J]. 安徽農業科學, 2014, 42(22): 7367–7369.
[3] 靳治國. 耐鉛鎘菌株的篩選及其在污染土壤修復中的應用[D]. 重慶: 西南大學, 2010.
[4] DONMEZ G and KOCBERBER N. Bioaccumulation of hexavalent chromium by enriched microbial cultures obtained from molasses and NaCl containing media[J]. Process Biochemistry, 2005, 40(7): 2493–2498.
[5] 趙素麗, 譙華. 生物修復技術的發展現狀[J]. 重慶工業高等專科學校學報, 2004, 19(5): 14–16.
[6] 王婷. 高效誘變菌與生物炭復合修復重金屬污染土壤的研究[D]. 天津: 南開大學, 2013.
[7] 張敏, 郜春花, 李建華,等. 一株耐鉛土著微生物的吸附特性及機制研究[J]. 山西農業科學, 2018, 46(8): 1321– 1328.
[8] SIGUA G C, STONE K C, HUNTP G, et al. Increasing biomass of winter wheat using sorghum biochars[J]. Agronomy for Sustainable Development, 2015, 35(2): 739–748.
[9] 周宇杰, 趙文, 羅春巖, 等. 有機肥對鉛在土壤中形態分配的影響[J]. 環境化學, 2018, 37(3): 534–543.
[10] DENG Zujun, ZHANG Renduo, SHI Yang, et al. Enhancement of phytoremediation of Cd-and Pb-contaminated soils by self-fusion of protoplasts from endophytic fungus Mucor sp. CBRF[J]. Chemosphere, 2013, 91(1): 41–47.
[11] UDOVIC M, DROBNE D, LESTAN D. An in vivo invertebrate bioassay of Pb, Zn and Cd stabili-zation in contaminated Soil[J]. Chemosphere, 2013, 92(9): 1105–1110.
[12] 關松蔭, 張德生, 張志明. 土壤酶及其研究方法[M]. 北京: 中國農業出版社, 1986.
[13] 楊蘭芳, 曾巧, 李海波, 等. 紫外分光光度法測定土壤過氧化氫酶活性[J]. 土壤通報, 2011, 42(1): 207–210.
[14] NASEEM H, BANO A. Role of plant growth- promoting rhizobacteria and their exopolysaccharide in drought tolerance of maize[J]. Journal of Plant Interactions, 2014, 9(1): 689–701.
[15] SCHUTTER M, Dick R. Shifts in substrate utilization potential and structure of soil microbial communities in response to carbon substrates[J]. Soil Biology& Bioche-mistry, 2001, 33(11): 1481–1491.
[16] 曹書苗. 放線菌強化植物修復土壤鉛鎘污染的效應及機理[D]. 西安: 長安大學, 2016.
[17] 馬克平, 黃建輝, 于順利, 等. 北京東靈山地區植物群落多樣性的研究Ⅱ.豐富度、均勻度和物種多樣性指數[J]. 生態學報, 1995, 15(3): 268–277.
[18] RAUNKAIER C. The life form of plant and statistical plant geography[M]. Oxford: Clarendon Press, 1934: 623.
[19] 李紅, 區杰泳, 顏增光, 等. 牛骨炭與伊/蒙黏土組配改良劑對土壤中Cd 的鈍化效果[J]. 環境科學研究, 2018, 31(4):725–731.
[20] 董同喜, 張濤, 李洋, 等. 畜禽糞便有機肥中重金屬在水稻土中生物有效性動態變化[J]. 環境科學學報, 2016, 36(2): 621–629.
[21] 江春玉, 盛下放, 何琳燕, 等. 一株鉛鎘抗性菌株WS34的生物學特性及其對植物修復鉛鎘污染土壤的強化作用[J]. 環境科學學報, 2008, 28(10): 1961–1968.
[22] 石汝杰. 草坪植物對鉛的吸收積累及其根際效應[D]. 貴陽: 貴州大學, 2005.
[23] 鐘曉蘭, 周生路, 李江濤, 等. 土壤有效態Cd、Cu、Pb的分布特征及影響因素研究[J]. 地理科學, 2010, 30(2): 254–260.
[24] 王秀梅, 安毅, 秦莉, 等. 對比施用生物炭和肥料對土壤有效鎘及酶活性的影響[J]. 環境化學, 2018, 37(1): 67– 74.
[25] 王期凱, 郭文娟, 孫國紅, 等. 生物炭與肥料復配對土壤重金屬鎘污染鈍化修復效應及酶活性的影響[J]. 農業資源與環境學報, 2015, 32(6): 583–589.
[26] 鄭少玲, 陳瓊賢, 馬磊, 等. 施用生物有機肥對芥藍及土壤重金屬含量影響的研究[J]. 農業環境科學學報, 2005, 24(增刊): 62–66.
[27] FARFEL M R, ORLOVA A O, CHANEY R L, et al. Biosolids compost amendment for reducing soil lead hazards: a pilot study of Orgro amendment and grass seeding in urban yards[J]. Science of the Total Environment, 2005, 340(1): 81–95.
[28] 楊海征. 雞糞堆肥對重金屬污染土壤茼蒿品質,土壤 Cu、Cd 形態和酶活性影響[D]. 武漢: 華中農業大學, 2009.
[29] 楊繼飛. 菌肥對鉛污染土壤中玉米生物效應的研究[D]. 太原: 山西農業大學, 2015.
[30] 李建華. 微生物菌劑對礦區復墾土壤的生態效應研究[D]. 太原: 山西大學, 2009.
[31] 牛旭. 生物有機肥對礦區復墾土壤微生物多樣性及小麥生長的影響[D]. 太原: 山西大學, 2014.
[32] 張連忠, 路克國, 王宏偉, 等. 重金屬和生物有機肥對蘋果根區土壤微生物的影響[J]. 水土保持學報, 2005, 19(2): 92–95.
Bioremediation agent reduces the stress of lead in polluted soil on rapeseed
Gao Yajing1, Li Jianhua2, Lu Jinjing2, Jin Dongsheng2, Gao Chunhua2*
1. College of Biological Engineering, Shanxi University, Taiyuan 030006, China 2. Institute of Agricultural Environment & Resources, Shanxi Academy of Agricultural Sciences, Taiyuan 030031, China
In order to study the repairing effect of bio-remediation agent TF3(0.5 g·kg-1enterobacteriaceae and 20 g·kg-1bio-char and 20 g·kg-1organic fertilizer) on lead-contaminated soil, five lead concentrationswere selected. The pot experiment method was used to study the enrichment and transport of lead in rapeseed and the changes of soil available lead, the enzyme activity and microbial response after application of repair agent TF3. The results showed that TF3 could promote the growth of rapeseed and significantly reduce the lead content in the above ground and under-ground parts of rapeseed. Compared with CK group, the above ground, under-ground and total enrichment factors of TF3 group were decreased by 20.12% to 71.91%, 37.75% to 60.13% and 22.45% to 68.77%, respectively. The TF3 group significantly reduced the effective lead concentration in the soilby 23.10% to 39.84%. With the addition of TF3, the four enzyme activities and the number of microorganisms in the soil were increased to different degrees. The fungi, actinomycetes and bacteria were increased by 11.76% to 40.00%, 6.45% to 25.61% and 120.20% to 290.24%, respectively. Our results indicated that TF3 reduced the stress of lead-contaminated soil on rapeseed and could be used as an effective bio-remediation agent for controlling lead pollution.
bio-remediation agent; lead content; contaminated soil; rape; enrichment factor
10.14108/j.cnki.1008-8873.2020.02.015
X53
A
1008-8873(2020)02-114-10
2019-06-01;
2019-10-10
國家重點聯合基金項目“復墾土壤功能微生物演變特征及定向調控 ”(U1710255); 山西省農科院農業科技創新研究課題“礦區復墾土 壤功能微生物定向調控技術研究”(YCX2018DZYS02);山西省科技成果轉化引導專項項目"礦區復墾土壤快速培肥技術示范推廣"(201804D131049)
郜雅靜(1993—), 女, 在讀研究生, 主要從事污染土壤生物修復研究, E-mail: 884076352@qq.com
郜春花(1963—), 女, 本科, 研究員, 主要從事農業微生物研究, E-mail: chunhuagao@163.com
郜雅靜, 李建華, 盧晉晶, 等. 生物修復劑TF3對鉛污染土壤的修復效果研究[J]. 生態科學, 2020, 39(2): 114–123.
Gao Yajing, Li Jianhua, Lu Jinjing, et al. Bioremediation agent reduces the stress of lead in polluted soil on rapeseed[J]. Ecological Science, 2020, 39(2): 114–123.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
小學科學(學生版)(2021年7期)2021-07-28 06:44:32
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
小學科學(學生版)(2020年6期)2020-08-13 07:47:12
航空世界(2020年10期)2020-01-19 14:36:20
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
