基于18S rRNA序列的云南大葉千斤拔與細葉千斤拔根內叢枝菌根真菌鑒定
2020-04-19 10:08:13周麗思郭順星
世界中醫藥 2020年5期
周麗思 郭順星

摘要?目的:調查云南省大葉千斤拔(Flemingia macrophylla)與細葉千斤拔(Flemingia lineata)根內叢枝菌根真菌(Arbuscular Mycorrhizal Fungi,AMF)群落結構多樣性。方法:使用巢式-PCR、克隆、限制性片段長度多態性(Restriction Fragment Length Polymorphism,RFLP)分析及測序技術。結果:共獲得558個含有叢枝菌根真菌18S rRNA片段的克隆子,經RFLP分析后得到83個RFLP類型,DNA序列分析可將其劃分為23個可操作分類單元(Operational Taxonomic Units,OTUs),分屬于5個科,Glomeraceae為優勢類群。在MaarjAM數據庫中進行比對后,23個OTUs可鑒定為18個虛擬分類分子種,分布于13種不同的生境。經統計分析大葉千斤拔與細葉千斤拔根內叢枝菌根真菌群落組成比較,差異有統計學意義(P<0.001)。結論:比較大葉千斤拔與細葉千斤拔根內叢枝菌根真菌群落差異,結合叢枝菌根真菌在生態系統中的分布特點,為種植千斤拔屬植物的選址提供有力地環境指標數據,并為篩選促生菌株提供依據。
關鍵詞?千斤拔屬;叢枝菌根真菌;多樣性;分布;分子鑒定;巢式-PCR;限制性片段長度多態性分析;可操作分類單元
Identification of Arbuscular Mycorrhizal Fungi of Flemingia macrophylla and Flemingia lineata in Yunnan, Based on 18S rRNA Sequences
ZHOU Lisi, GUO Shunxing
( Institute of Medicinal Plant Development, Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing 100193, China )
Abstract?Objective:To investigate the community diversify of arbuscular mycorrhizal fungi(AMF) of Flemingia macrophylla and Flemingia lineata.Methods:The nested-PCR, cloning, RFLP analyzing and sequencing techniques were used.Results:A total of 558 clones containing 18S rRNA genes from AMF were obtained.After RFLP analysis, 83 different RFLP types were obtained, which can be divided into 23 Operational Taxonomic Units (OTUs) by DNA sequence analysis.They belonged to 5 subjects, and Glomeraceae was the dominant group.After they were compared in MaarjAM database, 23 OTUs could be identified as 18 molecular virtual taxon, scattered in 13 different habitats.By statistical analysis, AMF communities were significantly different between F.macrophylla and F.lineata roots, the difference was statistically significant (P<0.001).Conclusion:Compared with the difference of AMF community in F.macrophylla and F.lineata roots, combining with the distribution features of AMF species in ecosystem, valuable environmental indicator data to choose site for the Flemingia plantation is provided.
Keywords?Flemingia; Arbuscular mycorrhiza; Diversity; Distribution; Molecular identification; Nest-PCR; RFLP; Operational taxonomic unit
中圖分類號:R282.2文獻標識碼:Adoi:10.3969/j.issn.1673-7202.2020.05.002
千斤拔屬(Flemingia)植物多為灌木或亞灌木,全球有40余種,主要分布于亞洲、非洲、大洋洲的熱帶地區,我國有16種及1變種[1]。2015版《中華人民共和國藥典》收載千斤拔藥材來源為豆科植物蔓性千斤拔Flemingia pholippinensis、大葉千斤拔Flemingia macrophylla和繡毛千斤拔Flemingia ferruginea的干燥根。此外,《中國植物志》《現代中藥學大辭典》和《藥用植物詞典》中指出大部分千斤拔屬植物均可供藥用。千斤拔及相關中成藥產品主要用于婦科疾病、風濕痹痛、體虛貧血、坐骨神經痛、咽喉腫痛、跌打損傷等方面的治療[2-3]。分離自千斤拔屬植物的化合物主要包括黃酮類、酚類、甾體類、蒽醌類、揮發油類等[4],其中異戊烯基取代黃酮類成分是千斤拔屬植物的主要成分和特征成分[5]。相關藥理研究表明黃酮類化合物具有顯著的抗氧化、抗腫瘤、保護肝臟及心血管系統的活性[6-7];萜類與甾類等化合物具有抗菌驅蟲、抗氧化等作用[8]。隨著千斤拔相關產品的開發,市場需求量日益增大,千斤拔屬植物的人工繁殖勢在必行,我國已相繼開展了種子繁殖、離體繁殖、扦插繁殖等千斤拔人工栽培技術研究。有研究表明,千斤拔屬植物有效成分的積累與種質資源和環境條件都息息相關[9-10]。叢枝菌根真菌作為與植物關系最為密切的土壤微生物,可與絕大多數植物形成共生體,從而影響植株的生長微環境[11]。
叢枝菌根真菌作為“生物肥料”已廣泛應用于農作物生產[12],其不僅改善植物營養狀況、促進植物對養分元素的吸收和代謝、同時提高植物對不良環境和病蟲害的抵抗能力。因此,叢枝菌根真菌在藥用植物栽培,尤其是根、莖為主要藥用部位的藥用植物人工繁殖方面的應用研究也逐漸受到關注。相關研究表明,接種叢枝菌根真菌之后,可提高半夏塊莖中鳥苷、生物堿等化學成分含量[13];可以顯著提高川黃柏幼苗的小檗堿含量[14];可增加北柴胡皂苷A和地上部分黃酮含量[15]。明確原生地藥用植物根內叢枝菌根真菌群落組成,將有助于了解該植物與土壤微生物處于最佳關系時的微生物群落結構,為優化人工栽培環境提供有力地理論依據。
得益于分子生物學技術日益成熟,對不可純培養的叢枝菌根真菌群落結構的鑒定方法也不僅限于對根外孢子的傳統形態學顯微觀察法,根內叢枝菌根真菌群落組成亦成為關注焦點[16]。前者可獲得優質的叢枝菌根真菌種質資源便于開展應用研究,后者可直接獲得根內叢枝菌根真菌的群落結構信息,真實反應與植物直接發生作用的叢枝菌根真菌種類[17]。本文分別以2015版《中華人民共和國藥典》收載物種大葉千斤拔和非藥典收載的藥用物種細葉千斤拔為研究對象,通過根內叢枝菌根真菌18s rRNA克隆文庫構建及RFLP分析,考察二者根系中叢枝菌根真菌群落結構及其種屬間的差異,旨在為解決千斤拔栽培中的相關問題提供理論依據。
1?材料與方法
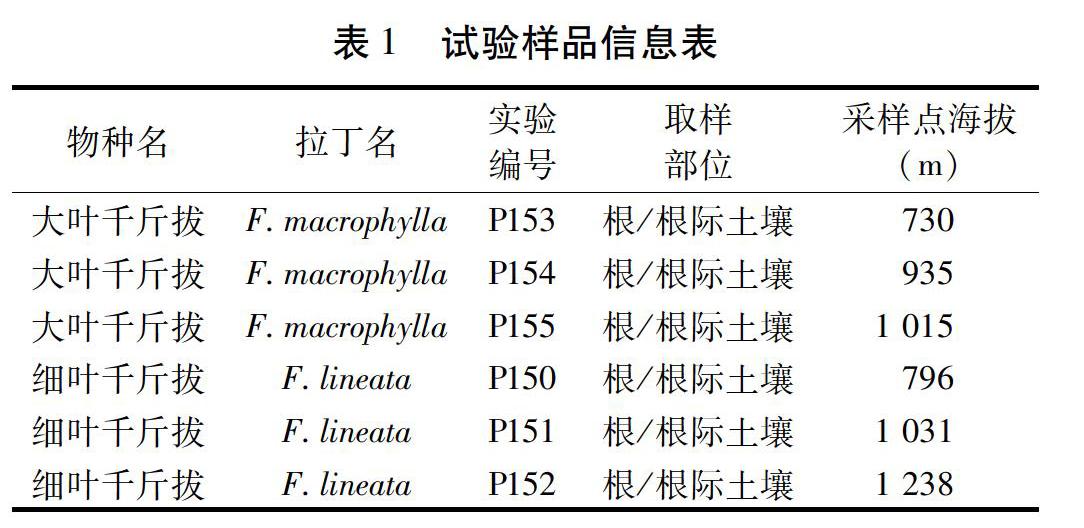
1.1?樣品采集及處理?大葉千斤拔和細葉千斤拔樣品均于2012年1月采自景洪至勐海縣的G214國道沿途。見表1。采樣地位于云南省西雙版納傣族自治州,地處北回歸線以南的熱帶北部邊緣,全年干濕季分明,降雨主要集中在5月下旬至10月下旬,全年平均降雨量為1 557 mm,全年平均氣溫為21.5 ℃,屬熱帶季風氣候[18]。隨機選取植株生長的3個地點,在根圍選擇5個采樣點,每個采樣點采集一份根系樣本,將5個采樣點的根系樣品混合后作為該生長地點的一份根系樣品。在距植株30~50 cm處采集20~30 cm深處的土樣和細根樣,編號、裝入自封袋后置入冰盒帶回。將根系樣品清洗干凈后于-80 ℃冰箱中保存備用。土樣風干保存。
1.2?叢枝菌根侵染相關指標測定
1.2.1?侵染強度的測定?每份根系樣品選取30個1 cm左右的幼嫩根系,經0.05%曲利苯藍染色后,在顯微鏡下測定侵染率。根據菌根侵染分級標準和叢枝泡囊豐度分級標準對根段進行侵染率級別判斷,使用“MYCOCALC”軟件,計算侵染強度(M%),該指標是真菌侵入植物根系的頻度與強度的綜合反應[19]。
1.2.2?菌絲密度的測定?稱取5 g過1 mm篩的土壤樣品,采用雙層篩過濾法提取根外菌絲至微孔濾膜上,0.05%曲利苯藍染色后觀察菌絲[20]。每張微孔濾膜上選取均勻分布的25個樣點進行觀察、統計[19]。
1.2.3?孢子密度的測定?稱取10 g風干土樣,濕篩傾析-蔗糖離心法分離叢枝菌根真菌孢子,在體視顯微鏡下計數[20]。
1.3?根內叢枝菌根真菌分子多樣性
1.3.1?根系樣品DNA的提取?取約200 mg根系樣品,經表面消毒后(75%乙醇30 s、1%次氯酸鈉3 min、無菌水沖洗3次),用CTAB法提取植物基因組試劑盒(北京艾德萊生物科技有限公司,中國,批號:181953)并按照操作手冊進行根系樣品DNA提取,于-20 ℃保存備用。
1.3.2?巢式PCR擴增?為更全面地獲得根內叢枝菌根真菌群落組成,第1次PCR所用引物為叢枝菌根真菌特異引物AML1/AML2[22]表2、擴增模版為根系樣品總DNA,第2次PCR所用引物為叢枝菌根真菌特異引物NS31/AM1[23-24]、擴增模版為稀釋1 000倍后的第1次PCR產物。按照EasyTaq KIT說明書(北京全式金生物技術有限公司,中國,批號:G21017)依次進行PCR反應。2次PCR產物均用1%瓊脂糖凝膠電泳進行檢測。每份DNA樣品進行3次技術重復。
1.3.3?RFLP分析與克隆文庫構建?合并每份樣品的3個技術重復的PCR產物,并使用膠回收試劑盒進行純化(北京艾德萊生物科技有限公司,中國),純化產物用于構建叢枝菌根真菌18S rRNA克隆文庫(共6個文庫)。經藍白斑篩選,將陽性轉化子進行擴增后,用HinfI(Fermentas,立陶宛)進行RFLP分析。使用2%瓊脂糖凝膠電泳對酶切產物進行檢測并劃分RFLP譜型,RFLP譜型僅在同一克隆文庫內比較,不合并不同克隆文庫間的RFLP譜型。每一RFLP譜型隨機挑取一個克隆子進行DNA序列測定(蘇州金唯智生物科技有限公司,中國),若測序結果不好或不能測出的克隆,則選取相同RFLP類型的另一克隆子進行測序。
1.3.4?序列分析與系統發育樹構建?使用BLAST軟件與GenBank數據庫進行在線比對,去除非叢枝菌根真菌序列。利用DUTOR軟件按序列相似水平進行序列類群劃分,相似性大于97%的序列劃分為同一相似水平的OTUs。按所劃分的可操作分類單元對Shannon多樣性指數(H′)、相對多度(Relative Abundance)、重要值(Important Values)進行統計[20]。采用ClusterX軟件對代表序列與參考序列處理后,用Mega7.0軟件進行Neighbor-Joining系統發育分析。隨后,將本研究所獲得的代表序列與MaarjAM數據庫[25](http://maarjam.botany.ut.ee)進行比對,序列相似度大于97%鑒定為一個分子虛擬分類種(Molecular Virtual Taxon)。
1.4?數據處理?采用Microsoft Excel 2017軟件對數據進行處理和繪圖。使用SPSS 22.0統計分析軟件對大葉千斤拔和細葉千斤拔根內叢枝菌根真菌侵染率、孢子密度、菌絲密度等數據進行差異顯著性檢驗(LSD法),使用Fisher確切概率法對大葉千斤拔與細葉千斤拔根內叢枝菌根真菌序列類群對應克隆數及不同科叢枝菌根真菌的差異顯著性進行檢驗[26-27]。
2?結果與分析
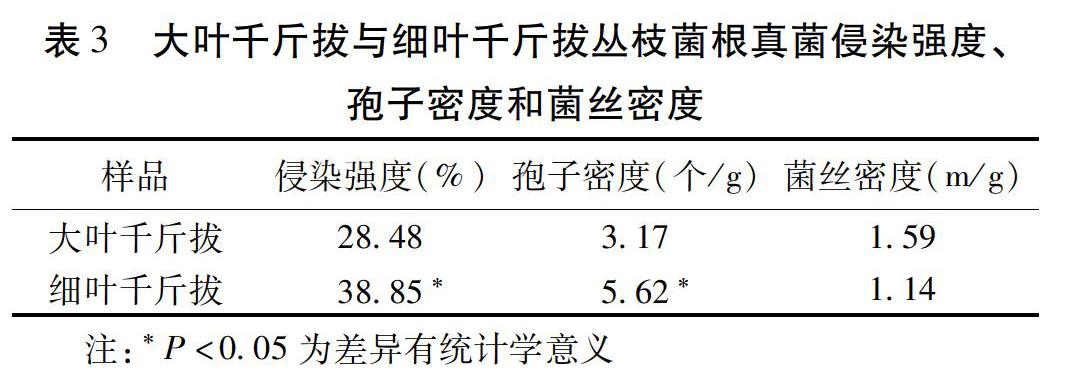
2.1?根際叢枝菌根真菌概況?大葉千斤拔與細葉千斤拔根際叢枝菌根真菌的侵染強度、孢子密度和菌絲密度等數據見表3。大葉千斤拔除了菌絲密度與細葉千斤拔比較差異無統計學意義,侵染強度和孢子密度均顯著低于細葉千斤拔。
2.2?根內叢枝菌根真菌克隆文庫的構建及其群落多樣性分析?全部大葉千斤拔與細葉千斤拔根系樣品均成功擴增出根內叢枝菌根真菌18S rRNA序列。采樣點分別建立克隆文庫。見表4。每個物種有3個采樣點樣本,每個樣本選出94個陽性克隆,2個物種3個采樣點樣本的總克隆數均為282個,測序后舍去非叢枝菌根真菌序列。大葉千斤拔樣品克隆文庫獲得276個克隆,細葉千斤拔樣品獲得282個克隆。克隆文庫的覆蓋度均在97%以上,可以較好地代表各樣本中叢枝菌根真菌的多樣性。
通過RFLP條帶類型,進一步劃分克隆子類型;大葉千斤拔樣本有48個RFLP條帶類型,細葉千斤拔樣本有35個,共獲得83條測序結果。按97%相似度劃分可操作分類單元(OTU),共劃分23個OTUs,每個OTU中隨機選取一條序列為代表序列(GenBank號:MN904800-MN904822);大葉千斤拔根系樣本含16個OTUs,細葉千斤拔根系樣本含12個OTUs。綜合每個物種的3個采樣點結果,大葉千斤拔的Shannon多樣性指數為1.80,細葉千斤拔則為1.13。以上結果表明大葉千斤拔根內叢枝菌根真菌種類較為豐富,但與細葉千斤拔比較差異無統計學意義(P>0.05)。
2.3?系統發育分析與叢枝菌根真菌群落組成?對已獲得23個序列類型中的代表序列進行系統發育分析,見圖1,大葉千斤拔與細葉千斤拔根內叢枝菌根真菌種類分屬于5個科,19個屬;經Blast序列比對,僅6個可以鑒定到種(Bootstrap>85)。6個可鑒定到種的可操作分類單元中見表5,有1個優勢類群(Rhizophagus fasciculatus),2個常見類群(Acaulospora spinosa、Rhizophagus intraradices),1個偶見類群(Glomus indicum),2個稀有類群(Claroideoglomus lamellosum、Glomus viscosum)。
與MaarjAM數據庫比對,18個序列類群均可以劃分為虛擬分子分類種(相似度>97%,見表5),且無分屬于同一虛擬分子分類種的現象。通過構建系統發育樹以及與虛擬分類分子種比對2種方法均未能對可操作分類單元成功鑒定的有5個OTUs。通過虛擬分子種的劃分,將未能通過構建系統發育樹鑒定到種的4個OTUs、且在大葉千斤拔與細葉千斤拔根內為常見類群的可操作分類單元進行虛擬分類(OTU5、OTU6、OTU9、OTU12)。通過MaarjAM數據庫還可以獲得對應虛擬分子分類種所處生態系統及生境的信息。在18個比對到虛擬分子分類種的序列類群中,6個序列類群所對應虛擬分子分類種所處生境為熱帶生態系統,包含7.89%的克隆子;7個虛擬分子分類種分布在亞熱帶生態系統,包含32.97%的克隆子;4個虛擬分子分類種分布在亞熱帶生態系統,包含41.40%的克隆子。
2.4?大葉千斤拔與細葉千斤拔根內叢枝菌根真菌分布的差異?大葉千斤拔與細葉千斤拔根內叢枝菌根真菌分布相對多度見圖2所示。所獲得的23個序列類型在大葉千斤拔和細葉千斤拔根內的分布比較,差異有顯著統計學意義(P<0.001),在分布種類和相對多度均有很大差別。在大葉千斤拔根內16個序列類型中,11個序列類型僅在大葉千斤拔根系樣品中出現,相對多度最高的3個序列類型依次為OTU22(9.06%)、OTU12(8.33%)、OTU4(7.61%);在細葉千斤拔根內12個序列類型中,7個序列類型僅在細葉千斤拔根系樣品中出現,相對多度最高的3個序列類型依次為OTU3(15.25%)、OTU6(7.45%)、OTU11(2.48%);OTU7(大葉千斤拔15.94%、細葉千斤拔48.23%)、OTU8(大葉千斤拔14.86%、細葉千斤拔1.06%)、OTU9(大葉千斤拔1.81%、細葉千斤拔18.09%)、OTU5(大葉千斤拔9.78%、細葉千斤拔3.90%)在2種樣品中均有出現。
本文所獲得的23個序列類群分屬于5個科,Glomeraceae在大葉千斤拔和細葉千斤拔根內的相對多度均大于90%,是二者根內共有種類,占絕對優勢。大葉千斤拔根內特有種類為Diversisporaceae,是偶見類群;細葉千斤拔根內特有種類為Acaulosporaceae、Ambisporaceae、Claroideoglomeraceae,前者為常見類群,后二者為稀有類群。
3?討論
通過Nested-PCR、RFLP與克隆測序技術相結合,使用叢枝菌根真菌特異性引物進行兩次擴增,盡可能全面地獲得根內不可培養叢枝菌根真菌的群落組成情況,并從大葉千斤拔和細葉千斤拔根內獲得23個OTUs,分屬于5個科,其中Glomeraceae在大葉千斤拔和細葉千斤拔根內占絕對優勢。由于Glomeraceae科叢枝菌根真菌可通過在根內產生泡囊結構儲存脂類物質幫助菌絲生長及再侵染[28],因此Glomeraceae科的繁殖策略更適合在經常被擾動的生態系統中存活[29]。已有研究表明,寬葉千斤拔在接種Glomeraceae科Funneliformis mosseae和Glomus versiforme可顯著促進植株生長和藥用部位的產量[30]。23個序列類型中6個序列類型鑒定到種,其中5種叢枝菌根真菌也廣泛存在于我國其他藥用植物根際土壤中:A.spinosa在人參[31]、三七[32]、三角葉黃連[33]等9種藥用植物根圍土壤中獲得;C.lamellosum則在石菖蒲[34]、薄荷[34]、喜樹[35]、蒙古扁桃[36]等藥用植物根圍土壤中發現;R.intraradices則在半夏[37]、糯米團[34]、紫茉莉[38]、連香樹[39]等13種藥用植物根圍土壤中獲得;R.fasciculatus則在人參、五味子、地黃、盾葉薯蕷等21種藥用植物根圍土壤中獲得[40];G.viscosum則在薯蕷、狗脊、南方紅豆杉等5中藥用植物根圍土壤中獲得[41];而G.indicum則未在我國已報道、分離叢枝菌根真菌的171中藥用植物根圍土壤中獲得,但其在大葉千斤拔根內的相對多度較高,在篩選大葉千斤拔促生菌株時應給予重視。另外,接種R.intraradices在高溫脅迫下可促進蒼術根莖生物量的積累并顯著影響根莖揮發油總量和揮發油主要成分[42],且可顯著提高庫頁紅景天的凈光合速率、根莖生物量及成活率[43]。同時該菌株為大葉千斤拔和細葉千斤拔根內的常見類群,應積極考察其對千斤拔屬植物的促生長和有效成分積累的作用,并關注該屬其他菌株與千斤拔屬植物的互作情況。
不可純培養的叢枝菌根真菌的鑒定早期主要是從根際土壤分離孢子進行形態學種類劃分,導致產孢數量少或不產孢的種類并沒有被發現。隨著分子生物學的發展,越來越多研究表明植物根系中分布的叢枝菌根真菌群落與土壤中分布的群落有很大差異[44]。由此,基于分子生物學技術對根內叢枝菌根真菌群落組成進行考察,所獲的優勢類群往往無法被鑒定到種的水平。利用MaarjAM數據庫、基于18S rRNA基因序列將叢枝菌根真菌的序列類型進行虛擬分類,可以更充分地挖掘分子鑒定的數據,使未能鑒定到種的序列類型也能找到與之相對應的虛擬分類分子種,并發現其在全球范圍內的分布特點和生態學功能。在所獲得的23個OTUs中,僅有6個OTUs可以比對到已獲得孢子的形態鑒定物種,剩下17個OTUs的分類信息則無法確認。但通過MaarjAM數據庫比對,又獲得13個OTUs對應的虛擬分類分子種,該數據庫還提供虛擬分類分子種的各種相關信息,包括采樣點的生態系統類型、植被類型、相關已發表文獻等信息,可對分子種進行后續研究、分析。因此,大葉千斤拔和細葉千斤拔根系樣品中的絕大多數真菌群落得以鑒定,并對其棲息環境的偏好性及其作用有了一定的了解。大葉千斤拔根內叢枝菌根真菌種類更傾向于分布在人為擾動較小的野生環境中,而細葉千斤拔根內的叢枝菌根真菌則傾向于分布在受人類活動影響的生態系統中。分子種OTU6(VTX00093)分離自亞熱帶重金屬污染土壤,僅在細葉千斤拔根系中發現,表明該地土壤人為擾動因素較多。所處生境會直接影響根內叢枝菌根真菌群落組成,真菌種類又會直接影響與植物的互作結果[45]。喜樹幼苗分別接種6種叢枝菌根真菌后[46],發現均對喜樹堿的代謝有影響。與無菌根幼苗對比,除F.mosseae會引起喜樹堿含量降低外,其他5種叢枝菌根真菌種類都顯著提高了喜樹堿的含量,且物種之間是存在顯著差異的,還有叢枝菌根真菌對喜樹幼苗喜樹堿含量的影響有器官差異。因此,在千斤拔屬植物種植時,應根據與之互作的根內菌落分布特征,選取具有相應棲息偏好的種類作為篩選促生菌株的候選。對根內叢枝菌根真菌群落組成的鑒定,有助于全面地了解該植物所處生境,解讀更多利于植物生長的微環境變化。
綜上所述,本研究基于18S rRNA分子鑒定技術及MaarjAM數據庫,全面地對大葉千斤拔與細葉千斤拔根內叢枝菌根真菌組成進行分析,結合叢枝菌根真菌在生態系統中的分布特點,有助于對這些缺乏形態學研究的叢枝菌根真菌種類的相應生態功能進行總結分析,為種植大葉千斤拔的選址提供有力地環境指標數據,并為篩選促生菌株提供依據。
參考文獻
[1]中國科學院中國植物志編輯委員會.中國植物志[M].41卷.北京:科學出版社,1995:313.
[2]陳一,李開雙,黃峰嬌.千斤拔的鎮痛和抗炎作用[J].廣西醫學,1993,15(2):77-79.
[3]任朝琴,戴先芝,劉圓.千斤拔藥材資源開發與利用的調查報告[J].西南民族大學學報:自然科學版,2011,37(4):610-613.
[4]Cardillo G,Merlini L,Mondelli R.Natural chromenes-III:colouring matters of wars:the structure of flemingins A,B,C and homoflemingin[J].Tetrahedron,1968,24(1):497-510.
[5]Cheng S Y,Xie Y,Feng X L,et al.Study of the volatile constituents in Radix Flemingia macrophyllae and a substitute by gas chromatography-mass spectrometry and chemometric methods[J].Molecules,2012,17(12):14111-14125.
[6]Ko Y J,Lu T C,Kitanaka S,et al.Analgesic and anti-inflammatory activities of the aqueous extracts from three Flemingia species[J].Am J Chin Med,2010,38(3):625-638.
[7]Wang Y,Curtis-Long M J,Yuk H J,et al.Bacterial neuraminidase inhibitory effects of prenylated isoflavones from roots of Flemingia philippinensis[J].Bioorg Med Chem,2013,21(21):6398-6404.
[8]Ho H Y,Wu J B,Lin W C.Flemingia macrophylla extract ameliorates experimental osteoporosis in ovariectomized rats[J].Evidence-based Complementary and Alternative Medicine,2009,2011(18):752302.
[9]李昌松,張麗霞,趙俊凌,等.云南地區大葉千斤拔不同種質的異黃酮含量比較[J].中國現代中藥,2011,13(10):30-32,42.
[10]管志斌,張麗霞,高微微.大葉千斤拔種子萌發特性研究[J].中國農學通報,2011,27(13):116-120.
[11]Jansa J,Smith F A,Smith S E.Are there benefits of simultaneous root colonization by different arbuscular mycorrhizal fungi?[J].New Phytologist,2008,177(3):779-789.
[12]姜攀,王明元.廈門市七種藥用植物根圍AM真菌的侵染率和多樣性[J].生態學報,2012,32(13):4043-4051.
[13]郭巧生,程俐陶,劉作易.叢枝菌根真菌對半夏產量及化學成分的影響[J].中國中藥雜志,2010,35(3):333-338.
[14]周加海,范繼紅.AM真菌對川黃柏幼苗生長及小檗堿含量的影響[J].北方園藝,2007,31(12):25-27.
[15]滕華容.AM真菌與施磷量對柴胡生長和化學成分交互效應的研究[D].咸陽:西北農林科技大學,2005.
[16]Hijri I,Zuzana Sykorová,Oehl F,et al.Communities of arbuscular mycorrhizal fungi in arable soils are not necessarily low in diversity[J].Molecular Ecology,2006,15(8):2277-2289.
[17]趙婧,賀學禮.河北省安國市藥用植物AM真菌資源和分布研究[J].河北農業大學學報,2010,33(1),43-48.
[18]盧華正,沙麗清,王君,等.西雙版納熱帶季節雨林與橡膠林土壤呼吸的季節變化[J].應用生態學報,2009,20(10),2315-2322.
[19]Phillips JM,Hayman DS,1970.Improved procedures for clearing roots and staining parasitic and vesicular mycorrhizal fungi for rapid assessment of infection[J].Transaction of the British Mycological Society,55(1):158-161.
[20]周麗思,郭順星.云南西雙版納野生與栽培絞股藍根內叢枝菌根真菌的分子多樣性[J].應用生態學報,2013,24(9):2503-2510.
[21]Jakobsen I,Abbott LK,Robosen AD.External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L[J].New Phytologist,1992,120(3):371-380.
[22]Lee J,Lee S,Young JPW.Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi[J].FEMS Microbiology Ecology,2008,65(2):339-349.
[23]Simon L,Lévesque RC,Lalonde M.Identification of endomycorrhizal fungi colonizing roots by fluorescent single-strand conformation polymorphism-polymerase chain reaction[J].Applied and Environmental Microbiology,1993,59(12):4211-4215.
[24]Helgason T,Daniell TJ,Husband R,Fitter AH,Young JP.Ploughing up the wood-wide web?[J].Nature,1998,394(6692):431.
[25]pik M,Vanatoa A,Vanatoa E,et al.The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi(Glomeromycota)[J].New Phytologist,2010,188(1):223-241.
[26]Toljander JF,Santos-González JC,Tehler A,Finlay RD.Community analysis of arbuscular mycorrhizal fungi and bacteria in the maize mycorrhizosphere in a long-term fertilization trial[J].FEMS Microbiology Ecology,2008,65(2):323-338.
[27]Heuer H,Smalla K.Evaluation of community-level catabolic profiling using BIOLOG GN microplates to study microbial community changes in potato phyllosphere[J].Journal of Microbiological Methods,1997,30(1):49-61.
[28]Pietikinen A,Kytviita MM,Husband R.,Young JP.Diversity and persistence of arbuscular mycorrhizas in a low-arctic meadow habitat[J].New Phytologist,2007,176(3):691-698.
[29]Brundrett M,Abbott LK,Jasper DA.Glomalean mycorrhizal fungi from tropical Australia.I.Comparison of the effectiveness and specificity of different isolation procedures[J].Mycorrhiza,1999,8(6):305-314.
[30]譚鉅發,黃京華,扶蓉,等.接種AM真菌對四種藥用植物生長的影響研究初報[J].中藥材,2009,32(6),15-17.
[31]李香串.接種泡囊-叢枝菌根劑對人參產量的影響[J].中藥材,2003,26(7):475-476.
[31]任嘉紅,劉瑞祥,李云玲.三七叢枝菌根(AM)的研究[J].微生物學通報,2007,34(2):224-227.
[33]黃文麗,范昕建,嚴鑄云,等.三角葉黃連叢枝菌根真菌的多樣性研究[J].中草藥,2012,35(5):689-693.
[34]姜攀,王明元,盧靜嬋.福建漳州常見藥用植物根圍的叢枝菌根真菌[J].菌物學報,2012,31(5):676-689.
[35]于洋,于濤,王洋,等.接種后共培養時間對叢枝菌根喜樹幼苗喜樹堿含量的影響[J].生態學報,2012,32(5):1371-1377.
[36]王琚鋼.蒙古扁桃AMF多樣性及AM提高蒙古扁桃抗旱機制研究[D].呼和浩特:內蒙古大學,2011.
[37]程俐陶,郭巧生,劉作易.栽培及野生半夏叢枝菌根研究[J].中國中藥雜志,2010,35(4):405-410.
[38]王虹,李鶯,趙麗莉.VA菌根真菌對紫茉莉生長的影響[J].陜西農業科學,1999,45(1):22-23,41.
[39]王森,唐明,牛振川,等.山西歷山珍稀藥用植物AM真菌資源與土壤因子的關系[J].西北植物學報,2008,28(2):0355-0361.
[40]米芳珍.商洛地區8種藥用植物VA菌根真菌的資源調查[J].山西林業科技,2012,41(5):37-39.
[41]趙金莉,程學謙,顧曉陽,等.河北安國新“八大祁藥”根際AM真菌與土壤因子的關系[J].河南農業科學,2012,41(6):87-91.
[42]張霽,劉大會,郭蘭萍,等.不同溫度下叢枝菌根對蒼術根莖生物量和揮發油的影響[J].中草藥,2011,42(2):372-375.
[43]李熙英,黃世臣.叢枝菌菌根對1年生高山紅景天植株生長的影響[J].林業科技,2005,33(3):25-27.
[44]pik M,Metsis M,Daniell TJ,Zobel M,Moora M.Large-scale parallel 454 sequencing reveals host ecological group speci-city of arbuscular mycorrhizal fungi in a boreonemoral forest[J].New Phytologist,2009,184(2):424-437.
[45]Treseder KK,Cross A.Global distributions of arbuscular mycorrhizal fungi[J].Ecosystems,2006,9(2):305-316.
[46]趙昕.叢枝菌根真菌對喜樹幼苗的接種效應[D].哈爾濱:東北林業大學,2006.
(2020-02-10收稿?責任編輯:徐穎)
