AKR1B10基因沉默對肝癌細胞增殖和凋亡的影響
2020-04-21 13:08:54錢瑞坤馬長林
臨床肝膽病雜志 2020年4期
錢瑞坤, 馬長林, 喬 森
濟寧市第一人民醫(yī)院 肝膽外科, 山東 濟寧 272100
肝細胞癌(HCC)是肝臟中最常見的惡性原發(fā)性腫瘤,也是全球癌癥死亡的第二大主要原因。已發(fā)現(xiàn)醛酮還原酶家族1成員B10(aldo-keto reductase family 1 member B10,AKR1B10)在HCC、非小細胞肺癌、胰腺癌、乳腺癌中高表達[1-4]。
AKR1B10是醛酮還原酶(aldo-keto reductase,AKR)超家族的一員,AKR是一種以NADPH為輔酶的氧化還原酶基因超家族,可催化醛酮類物質轉化為相應的醇。AKR在不同的生物體內可參與多種生物學過程,包括羰基解毒、滲透調節(jié)、激素代謝、脂質合成、腫瘤的發(fā)生與治療[5]。AKR1B10在人類肝癌中高表達,但其調控機制仍不明確。為了解AKR1B10在肝癌中的致癌作用,本研究使用shRNA敲減方法進一步分析了AKR1B10沉默對肝癌生物表型的影響。
1 材料與方法
1.1 細胞培養(yǎng) HepG2及Huh7 HCC細胞株均在含有10%胎牛血清、青霉素(100 U/ml)和鏈霉素(100 g/ml)的DMEM培養(yǎng)基中培養(yǎng),放置于37℃、含有5%CO2的恒溫培養(yǎng)箱中。
1.2 AKR1B10敲減 將1×105Huh7及HepG2 HCC細胞接種在裝有2 ml新鮮完全培養(yǎng)基的6孔板中。過夜孵育后,除去培養(yǎng)基,加入2 ml含shRNA-AKR1B10慢病毒顆粒的培養(yǎng)基(含10 μg/ml polybrene)。24 h后,除去含有病毒的培養(yǎng)基,每孔加入2 ml新鮮完全培養(yǎng)基(DMEM+10%胎牛血清)。48 h后,用含有1 μg/ml嘌呤霉素的培養(yǎng)基篩選陽性細胞。
1.3 AnnexinV-APC/PI雙染法檢測細胞凋亡 穩(wěn)轉shRNA-scramble的HepG2及 Huh7細胞(對照組)及敲減AKR1B10的HepG2及 Huh7細胞(敲減組)消化后,收集5×105個細胞,AnnexinV-APC和PI(Abnova)染料雙染,利用流式細胞儀(BD Biosciences)進行檢測,具體的實驗步驟參考試劑盒說明書。
1.4 克隆形成實驗 取對數(shù)生長期的細胞,用胰酶消化后計數(shù)。在6孔板里進行,每孔鋪種Huh7細胞2000個,每孔2 ml培養(yǎng)液 (DMEM+10% FBS) 于培養(yǎng)箱中進行培養(yǎng),每3天更換1次培養(yǎng)液,培養(yǎng)10 d左右,棄培養(yǎng)液,用預冷的PBS洗滌2次之后,甲醇固定15 min,再用結晶紫染色10 min。在鏡下計數(shù),≥50個細胞形成的細胞團即為一個克隆。
1.5 檢測活性氧(ROS) 使用過氧化物敏感性熒光探針DCFH-DA檢測細胞內ROS的產生。 將細胞鋪在6孔板中,然后使用無血清的DMEM將DCFH-DA的終濃度調整為20 μmol/L,每孔加入1 ml DCFH-DA稀釋液,37 ℃孵育30 min,并用PBS洗滌3次。ROS水平通過流式細胞儀(BD Biosciences)測定。
1.6 患者肝癌組織及異體移植瘤模型 肝癌組織來自本院肝膽外科2018年1月-2018年6月收治的6例肝癌患者。本研究通過濟寧第一人民醫(yī)院倫理委員會審批[批號:2018倫審研第(017)號]及實驗動物倫理委員會審批[批號:2018倫審研第(045)號],所有患者均簽署知情同意書。實驗中使用了4~6周齡的BALB-c/Nude小鼠,購自上海南方模式生物科技股份有限公司。將AKR1B10敲減組以及對照組的Huh7細胞皮下注射到BALB-c/Nude雌性小鼠的左下肢(每只小鼠100 μl細胞懸液,包含4×106個細胞)。注射后第7天開始,每3天記錄1次腫瘤體積。
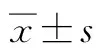
2 結果
2.1 AKR1B10在HCC細胞和HCC組織中的表達 CCLE數(shù)據(jù)庫提示AKR1B10 mRNA在HCC細胞中的表達明顯高于正常肝細胞(https://portals.broadinstitute.org/ccle)(圖1a)。Oncomine數(shù)據(jù)庫中的數(shù)據(jù)顯示,與癌旁組織(n=220)相比,HCC組織(n=225)中AKR1B10 mRNA的表達增加了10倍以上(圖1b)。使用UALCAN數(shù)據(jù)庫分析了HCC轉錄組數(shù)據(jù),提示AKR1B10 mRNA在HCC組織中高表達(圖1c)。同時,本研究在6例HCC患者中檢測到AKR1B10在HCC組織中的表達高于癌旁組織(圖1d)。
2.2 AKR1B10沉默對HCC細胞克隆、凋亡的影響 與對照組細胞相比,AKR1B10敲減的Huh7和HepG2細胞(使用不同的慢病毒敲減Huh7和HepG2細胞中的AKR1B10基因,分別標記為1#和2#)中AKR1B10的表達水平顯著下調(P值均<0.01)(圖 2a)。與對照組相比,敲減組中觀察到細胞集落形成的能力顯著下降(P值均<0.001)(圖2b,表1)。在AKR1B10敲減的情況下,肝癌細胞的凋亡率顯著增加(P值均<0.001)(圖2c,表1)。Western Blot表明,AKR1B10敲減后導致凋亡相關蛋白(包括剪切后的Caspase-3和RARP)的表達增加(圖3a)。

表1 敲減AKR1B10后HCC細胞克隆和凋亡的情況
注:與對照組比較,1)P<0.001。
2.3 AKR1B10敲減通過ROS超載引起廣泛的DNA損傷 如圖3b所示,AKR1B10敲減組細胞的ROS水平高于對照組。為了研究敲減AKR1B10的HCC細胞中升高的ROS水平是否引起DNA損傷,通過Western Blot檢測γ-H2AX和p-ATMser1981的水平以確定DNA損傷的存在。結果顯示,AKR1B10敲減可以導致p-ATMser1981和γ-H2AX的累積(圖3a)。

注:a,CCLE 數(shù)據(jù)庫中HCC細胞系中AKR1B10 mRNA水平; b,Oncomine數(shù)據(jù)庫中癌旁組織與HCC組織的AKR1B10 mRNA水平; c,UALCAN數(shù)據(jù)庫中HCC組織與癌旁組織的AKR1B10mRNA表達水平; d,Western Blot評估AKR1B10在本研究6例HCC患者癌旁組織(P)和癌組織(T)中的表達。
圖1AKR1B10在HCC中的表達情況
2.4 AKR1B10敲減可延遲腫瘤發(fā)生,降低HCC異種移植小鼠的腫瘤體積 將AKR1B10敲減組以及對照組的Huh7細胞皮下注射到BALB-c/Nude雌性小鼠的左下肢,28 d后拍照并剝離瘤體,如圖 4a~b 所示,AKR1B10敲減的HCC細胞致癌能力減弱,腫瘤體積減少。從異體移植后第13天開始,與對照組相比,AKR1B10敲減組腫瘤體積明顯縮小(P值均<0.05)(圖4c)。對照組瘤體石蠟包埋后,HE染色如圖4d所示。

注:a,Western Blot 檢測 AKR1B10敲減組細胞中AKR1B10蛋白水平; b,敲減組Huh7細胞集落形成的結果; c,流式細胞分析法檢測敲減組HepG2 以及Huh7 細胞的凋亡。
圖2敲減AKR1B10對HCC細胞克隆、凋亡的影響
3 討論
肝癌是全球第五大最常見的癌癥,已證實對公共健康構成嚴重威脅[6]。盡管當前的治療策略在過去幾十年中已有所改善,但長期生存率仍不令人滿意[7]。AKR1B10在HCC中過表達,在相鄰的肝組織中幾乎沒有表達。 HCC細胞中AKR1B10表達的沉默導致:(1)HCC細胞集落形成減少、細胞凋亡增加;(2)HCC細胞內氧化應激和DNA損傷增強;(3)延遲腫瘤發(fā)生和體內腫瘤負荷減少。這些結果均說明AKR1B10參與HCC的發(fā)生和發(fā)展。
為了闡明AKR1B10在HCC中的調控機制,筆者在GCBI網(wǎng)站(www.gcbi.com.cn)上預測AKR1B10參與多種代謝途徑,包括戊糖和葡萄糖醛酸酯相互轉化、果糖和甘露糖代謝、半乳糖代謝、甘油脂代謝、丙酮酸代謝。

注: a,流式細胞技術檢測AKR1B10敲減引起的細胞內ROS變化(空白組為Huh7細胞不與DCFH-DA共孵育,H2O2為陽性對照組); b,Western Blot檢測AKR1B10敲減的HCC細胞中DNA損傷和細胞凋亡相關蛋白表達水平。
圖3AKR1B10沉默后誘導ROS超載
AKR1B10可能通過調節(jié)相關代謝在腫瘤發(fā)生中起關鍵作用。前期研究[8-10]已經(jīng)報道了由于代謝過程中ATP耗竭引起ROS升高會導致癌細胞的凋亡增加。筆者推測,由AKR1B10敲減引起的ROS超負荷可能導致HCC細胞內的DNA發(fā)生氧化損傷。本研究中,通過DCFH-DA探針檢測發(fā)現(xiàn)AKR1B10敲減可誘導細胞ROS積累,從而導致DNA損傷。ATM是一種絲氨酸/蘇氨酸激酶,參與調節(jié)細胞周期檢查點控制、細胞凋亡和DNA修復[11]。在暴露的DNA雙鏈斷裂應力下,ATM在ser1981位點被自身磷酸化激活。γ-H2AX是一種變異的組蛋白,可響應DNA雙鏈斷裂而在ser139處被磷酸化激活,從而在斷裂點處形成病灶[12]。HCC細胞系中p-ATMser1981和γ-H2AX的增加更加確定了筆者的推論。據(jù)報道,與正常細胞相比,癌細胞的ROS水平較高[13]。穩(wěn)定的ROS水平還可以充當化學信使,激活NF-κB和p38 MAPK等信號通路,從而影響細胞增殖、分化和凋亡[14]。AKR1B10的高表達可以在HCC中維持相對穩(wěn)定的ROS水平。推測敲減AKR1B10引起異常代謝,然后增加氧化應激和廣泛的DNA損傷,導致細胞死亡。
綜上,本研究證明AKR1B10減少了氧化應激超負荷和DNA損傷,從而促進了HCC的發(fā)生和發(fā)展。AKR1B10是預后不良的有前途且具有高度潛力的生物標志物。基于這些發(fā)現(xiàn),AKR1B10是否為HCC的潛在新治療靶標有待于進一步驗證。


注:a~b,異體移植shRNA-AKR1B10和對照組小鼠的腫瘤體積情況; c,腫瘤生長曲線(與對照組相比,*P<0.05,**P<0.01,***P<0.001);d,Huh7皮下成瘤后的HE染色結果(×200)。
圖4敲減AKR1B10后異體移植HCC細胞成瘤體積的變化
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
天津醫(yī)科大學學報(2019年3期)2019-08-13 06:53:08
財經(jīng)(2017年2期)2017-03-10 14:35:35
財經(jīng)(2016年15期)2016-06-03 07:38:02
財經(jīng)(2016年3期)2016-03-07 07:44:46
財經(jīng)(2016年6期)2016-02-24 07:41:51
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫(yī)藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
