不同草田輪作模式土壤養分及細菌群落組成特征
2020-04-22 01:40:56尹國麗李亞娟張振粉李小龍張曉燕師尚禮
生態學報 2020年5期
尹國麗,李亞娟,張振粉,李小龍,張曉燕,師尚禮,*
1 甘肅農業大學草業學院, 蘭州 730070 2 草業生態系統教育部重點實驗室, 蘭州 730070
以紫花苜蓿(MedicagosativaL.)為代表的多年生豆科牧草是我國主要種植加工的一大類飼草作物,在現代養殖業發展中有著不可或缺的作用,但是連作障礙一直是多年生牧草種植的瓶頸問題。近些年已有大量學者進行了探索,證實連作障礙的發生不僅與土壤理化性質改變有關,還與土壤微生物組成有關[1-2],土壤微生物作為土壤中的活性因子,是反應土壤質量最敏感的指標,近年來已有大量的研究表明在土壤微生物與連作障礙之間存在密切關系[3-4]。
草田輪作是農業生產中消減連作障礙最廣泛的應用措施,輪作通過增加作物組分豐富了地上、地下生物多樣性,改變了土壤微生態環境[5],大量試驗證明,輪作不僅可以協調不同作物之間養分吸收的局限性,避免養分失衡,提高土壤養分的有效性和酶活性,還可以通過根系分泌物調整根圍微生物群落組成,減少土傳病蟲害的發生,提高經濟效益[6-7],但不同輪作模式對土壤理化性質的影響不盡相同。土壤微生物作為土壤生態系統重要組成部分,其群落結構和多樣性主要受植物物種組成和土壤養分狀況的影響[8],不同作物釋放的根系分泌物存在差別,也影響了土壤微生物群落生長代謝和群落結構[9-11]。細菌是土壤微生物中分布最廣,數量最多的部分[12],占土壤微生物總數的70%—90%,是土壤養分變化的敏感指標之一[13]。研究微生物群落組成及多樣性一直是揭示植物-微生物互作關系機制的熱點問題[14-15]。目前高通量測序技術的不斷發展,提高了研究者對環境中微生物群落組成和功能的認知水平[16-17]。
科學合理的輪作模式可有效緩解連作障礙引起的土壤環境惡化,使土壤中微生物活性增強、多樣性增加,為作物生長提供健康穩定的土壤生態環境[18-19]。目前關于紫花苜蓿草田輪作的研究報道相對較多,但前人的研究多集中在輪作對作物田土壤肥力及產量的影響[20-22],缺乏紫花苜蓿-作物輪作系統中土壤細菌組成方面的研究報道。鑒于此,本試驗分析5齡紫花苜蓿草地和不同作物輪作模式下,土壤養分因子含量和土壤細菌群落結構的組成及其相互關系,旨在較深層次上探索紫花苜蓿連作障礙的形成機制,為采用輪作措施減輕紫花苜蓿連作障礙提供實踐依據,并為半干旱地區草田輪作和草地生態農業的可持續發展提供理論基礎。
1 材料與方法
1.1 試驗地概況
試驗地位于在甘肅省武威市涼州區甘肅農業大學黃羊鎮牧草站(103°5′E,37°30′N,海拔1776 m)。位于甘肅河西走廊東端,屬冷溫帶干旱區,典型的大陸性氣候,日照充足,春季多風沙,夏季有干熱風。降水年際變化不大,但季節變化較大,多年平均降水量160 mm左右,主要集中在7、8、9月份,冬春季干旱,降水無法滿足作物生長的需要。年蒸發量2400 mm,年平均氣溫7.2 ℃,一月最低氣溫 -27.7 ℃,七月最高氣溫34.0 ℃,0 ℃積溫3513.4 ℃,10 ℃積溫2985.4 ℃。全年無霜期156 d,絕對無霜期118 d,年日照時數2945 h。土壤以荒漠灌淤土為主,粉沙壤質,土層深厚。
1.2 供試材料
供試材料小麥品種為雜交種春寧4號(TriticumaestivumL.cv.Chunning No.4),玉米品種為雜交種隴單4號(ZeamaysL.cv.Longdan No.4),紫花苜蓿品種為甘農3號(MedicagosativaL.cv.Gannong No.3)。小麥及玉米種子市售,紫花苜蓿種子由甘肅農業大學草業學院提供。
1.3 試驗設計
選取地勢平坦的紫花苜蓿草地進行草田輪作試驗。設計如表1所示,設4個處理。試驗小區面積為4 m×5 m,不同小區間設1 m隔離帶,每個處理3個重復,不同小區的土壤肥力及管理方式相同。

表1 紫花苜蓿-玉米/小麥不同輪作處理
1.4 土樣采集
于2017年7月中旬采集土壤樣品,五點法取樣,用內徑40 mm的土壤取樣器取0—30 cm土層(耕作層)土樣。每次采集后用70%的酒精對土壤采樣器進行消毒處理,以避免不同樣品之間的污染。采得土樣去除植物殘體和石礫等雜物后放入滅菌袋,低溫運輸至實驗室,盡快進行土壤DNA的提取和其余操作。
1.4.1土壤養分測定
土壤有機質含量采用重鉻酸鉀法測定;土壤全氮采用凱氏定氮法測定;土壤堿解氮采用堿解擴散法;土壤全鉀采用火焰光度法;土壤有效鉀含量先用醋酸銨浸提,再用火焰光度法測定;土壤全磷采用HClO4-H2SO4消煮法;土壤速效磷采用碳酸氫鈉浸提-鋁銻抗比色法測定[23]。
1.4.2土壤微生物DNA提取和 16S rRNA 基因高通量測序
將采集的樣品揀除雜質后,同一樣品充分混合使之均勻,準確稱取1.00 g新鮮土樣。采用OMEGA土壤DNA提取試劑盒提取土壤總DNA,提取步驟參考試劑盒說明書。1%的瓊脂糖電泳和紫外分光光度法檢測所提取DNA的質量和濃度,檢測合格后利用細菌16S rRNA基因的通用引物314F (5′-CCTACGGGNGGCWGCAG- 3′)
- 805R(5′-GACTACHVGGGTATCTAATCC- 3′)擴增土壤微生物16S rRNA V3+V4區基因片段。PCR反應體系如下:Phusion Master Mix(1X)15 μL,DNA模版(1 ng/μL)10 μL,正、反向引物(0.2 μmol/L) 各3 μL,ddH2O補至30 μL。PCR 擴增條件為:95℃,3 min;(95 ℃,30 s; 55℃,30 s;72 ℃,30 s),25個循環;16 ℃10 min[24]。每個樣品3個重復。瓊脂糖凝膠電泳法檢測擴增結果,檢測合格后送交至上海生工生物科技公司進行后續實驗和高通量測序。
1.5 序列分析

1.6 數據統計
采用Microsoft Excel 2013軟件對數據進行處理和作圖;用SPSS 24.0進行單因素方差分析(one-way ANOVA),采用最小顯著差異法(LSD)比較數據組間的差異,用Person相關系數評價不同因子間的相關性。用SPSS 24.0統計軟件完成相關關系分析。
2 結果與分析
2.1 不同輪作模式對土壤養分的影響
如表2所示,紫花苜蓿輪作作物1年時,A5C1處理的有機質含量為12.3 g/kg,A5W1處理為11.85 g/kg,輪作第2年,無論后茬作物是玉米還是小麥,有機質含量較輪作第1年均呈現下降趨勢,輪作玉米處理第2年比第1年下降3.66%,而小麥處理下降了7.68%。其余指標均呈現出相似的規律,就玉米輪作組而言,第2年與第1年相比,全氮下降了2.17%,全磷下降了9.28%,全鉀、堿解氮、速效磷、速效鉀呈現不同程度的下降,其中,下降幅度最大的是速效磷,下降了25.8%,下降幅度最小的是堿解氮,下降了0.47%。輪作小麥和輪作玉米表現出了相似的規律,都是輪作第2年各指標較第1年下降,下降幅度最大的是速效磷,下降了38.37%,下降幅度最小的是速效鉀,為2.53%。

表2 不同輪作模式下的土壤養分含量
同列不同字母表示處理間差異顯著(P<0.05)
2.2 不同輪作模式下土壤測序結果及深度
測序得到的土壤細菌群落序列讀數和OTUS數(表3),在相似度大于97%的水平上4個輪作模式共獲得高質量目標序列42743條。Clean Reads Rate值均高于80%,說明測序質量可以反映土壤細菌群落組成情況。不同輪作模式統計分析表明,土壤細菌OTUS種類數無顯著差異(P>0.05)。

表3 不同輪作模式下土壤細菌序列讀數及OTUS數
大寫字母表示同行之間比較, 小寫字母同列之間比較, 不同字母在 0.05 水平下差異顯著
2.3 不同輪作模式對土壤細菌群落Alpha多樣性的影響
由表4所示,除ACE指數外,其余3個指數都呈現出A5C2>A5W1>A5W2>A5C1的趨勢,ACE指數雖為A5W2>A5W1,但差異不顯著。綜上,不同指數反映出了基本一致的趨勢。

表4 不同輪作模式下土壤細菌群落Alpha多樣性比較
2.4 不同輪作模式對土壤細菌組成的影響
在門水平上,除極少部分細菌(0.02%—0.05%)未被分類外,4種輪作模式下土壤細菌主要分布在23個細菌門類。由圖1可知,優勢菌門為變形菌門(Proteobacteria)(25.30%—41.90%)和放線菌門(Actinobacteria)(14.50%—19.80%)以及厚壁菌門(Firmicutes)(3.70%—23.70%)、酸桿菌門(Acidobacteria)(5.40%—13.90%)。其次,擬桿菌門(Bacteroidetes)、綠彎菌門(Chloroflexi)、浮霉菌門(Planctomycetes)和芽單胞菌門(Gemmatimonadetes)的豐度也相對較高,其余菌門在各處理中均有分布,相對豐度因處理不同而存在差異,但差異不顯著(P>0.05)。另外,優勢菌在不同輪作模式下的組成也不同,A5W2中變形菌門(Proteobacteria)含量很高,而厚壁菌門(Firmicutes)含量卻較低,而這兩種菌在其他輪作模式下含量較為穩定。

圖1 不同輪作模式土壤在門水平上的細菌類群比較Fig.1 Comparison of bacteria groups at phylum level in the soils under different rotation sequencesA5W1: 5a紫花苜蓿- 1a小麥;5a紫花苜蓿- 2a小麥;A5C1: 5a紫花苜蓿- 1a玉米;A5C2: 5a紫花苜蓿- 2a玉米

圖2 不同輪作模式土壤在屬水平上的細菌類群比較Fig.2 Comparison of bacterial groups in the soils at the genus level under different rotation patterns
在屬的分類水平,除未被分類的細菌屬外(41.31%—53.78%),各輪作模式土壤細菌相對豐度排序前13的細菌菌屬分布見圖2。不同輪作模式下土壤優勢屬基本相同,其中丙酸桿菌屬(Propionibacterium)、芽孢桿菌屬(Bacillus)、鏈球菌屬(Streptococcus)以及奈瑟氏球菌屬(Neisseria)為優勢屬。但不同輪作模式下土壤各細菌屬所占相對豐度大小存在差異,如芽孢桿菌屬(Bacillus)在A5W1中相對豐度最高,但在A5W2中卻相對較低。
2.5 不同輪作模式下土壤細菌屬相對豐度的主成分分析
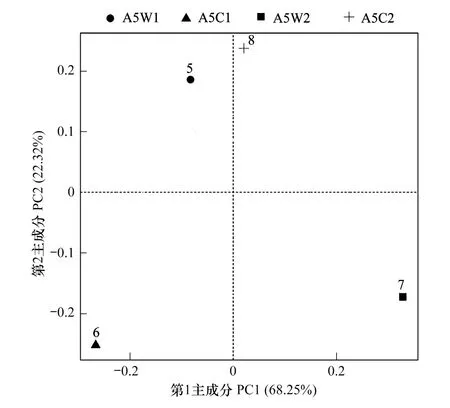
圖3 不同輪作模式土壤在屬水平細菌相對豐度主成分分析 Fig.3 Principal component analysis of relative abundance of soil bacteria at genus level under different rotation patternss
由圖3可知,4種輪作模式下土壤細菌物種相對豐度共提取4個主成分,累積貢獻率達96.82%,其中第1主成分(PC1) 和第2主成分(PC2)貢獻率分別為68.25%和22.32%。從圖3中可以看出4種輪作模式中A5W1和A5C1分布在PC1軸負方向,A5W2和A5C2分布在正方向。A5W1和A5C2序列分布在PC2軸正方向,A5C1和A5W2分布在負方向。
載荷因子反映主成分與細菌屬水平豐度的相關系數,載荷因子越高,表示該細菌屬對主成分的影響越大[12]。由表5可知,與第一主成分(PC1)具有較高相關性的細菌屬主要有固氮菌屬Azotobacter、硝化螺旋菌屬Nitrospira、小梨形菌屬Pirellula、Iamia菌屬和芽孢桿菌屬Bacillus,與第二主成分(PC2)具有較高相關性的細菌屬則主要有Flavisolibacter菌屬、德沃斯氏菌屬Devosia、德沃斯氏菌屬、拜納蒙納斯屬Balneimonas、Adhaeribacter菌屬(表5)。

表5 屬水平豐度的主成分載荷因子
表中帶的為兩個主成分中相關性前5的細菌屬
2.6 基于土壤細菌組成的不同輪作模式聚類分析
由圖4可見,在屬水平上,A5W1和A5C1的組成最接近,其次是A5C2,A5W2的組成與前兩者相差較大,說明作物輪作的時間越長,細菌組成差異越大。

圖4 不同輪作模式土壤在屬水平細菌組成聚類分析 Fig.4 Cluster analysis of genus level composition of soil bacteria under different rotation patterns
2.7 土壤養分與土壤細菌優勢屬間的相關關系
由表6可知,土壤養分因子與各細菌屬的相關性不同,有機質與丙酸桿菌屬Propionibacterium及固氮菌屬Azotobacter呈現顯著相關關系(P<0.05),而與芽孢桿菌屬Bacillus為極顯著正相關關系(P<0.01)。全鉀與鏈球菌屬Streptococcus呈現顯著相關關系(P<0.05),土壤堿解氮與丙酸桿菌屬Propionibacterium呈顯著正相關關系(P<0.05),與固氮菌屬Azotobacter呈現極顯著負相關關系(P<0.01);而速效磷與丙酸桿菌屬Propionibacterium、芽孢桿菌屬Bacillus及固氮菌屬Azotobacter均呈顯著正相關關系(P<0.05)。
3 討論
3.1 輪作模式和土壤細菌多樣性
土壤細菌作為土壤中最重要的活性組分,其多樣性水平也是反映土壤環境的最重要指標。通過本研究結果可以看出,后茬作物種類和輪作時間對土壤細菌群落結構都有明顯的影響。其他學者的研究也發現了相似的規律,如王芳等[25]利用Illumina Mi Seq技術對不同連作及輪作方式下大豆根際細菌進行研究,發現連作時間、輪作作物種類和輪作順序3個因素對對土壤細菌多樣性都會產生影響。梁志婷[26]在甘肅隴東地區進行了不同糧草輪作模式對土壤微生物多樣的研究結果也與本研究相似。本研究的結果顯示,輪作作物和輪作時間2個因子對土壤細菌多樣性的影響是普遍的,不同作物上的研究均支持這一結論。因此,在利用輪作改善土壤微生態時,應充分考慮2個因子,使其發揮最佳效果。另一方面,本研究也提出了新的科學問題,即在輪作體系中,作物種類和輪作時間這2個因子,哪一個對土壤微生物多樣性的組成影響更大,這將是值得深入研究的新課題。

表6 不同養分因子和幾種優勢細菌屬之間的相關性分析
*P<0.05;**P<0.01
3.2 不同輪作模式下土壤細菌群落組成
土壤細菌群落結構受種植作物的影響[27],作物種類不同其土壤微生物所處環境不同,細菌組成和豐度大小有差異,不同的農田土壤中存在共有和特異的細菌種群[28]。本研究發現變形菌門(Proteobacteria)和放線菌門(Actinobacteria)以及厚壁菌門(Firmicutes)、酸桿菌門(Acidobacteria)為各輪作順序豐度較高的菌,這一結果與Sapp等[29]、Liu等[30]和王伏偉等[31]對土壤細菌的研究結果一致。在屬的水平,芽孢桿菌屬(Bacillus)在耕層豐度較高,該屬為不產氧光合細菌,其功能與禾本科作物聯合固氮作用緊密相關[32],這與4個輪作順序取樣階段均為禾本科作物相符。研究表明,某些植物能夠與土壤微生物產生互作作用,對植物病害控制產生一定效果,同時也可能會引起某種作物感染病害[33]。本研究發現,5a紫花苜蓿- 1a玉米序列土壤中芽孢桿菌屬(Bacillus)和奈瑟氏球菌屬(Neisseria)豐度較高,該菌屬能夠通過產生幾丁質酶或細菌素的方式對土壤線蟲發育產生抑制作用,有生防效果[34-35]。研究結果在揭示在河西走廊灌區連作苜蓿后輪作禾本科作物在改善土壤肥力的同時,對胞土壤害蟲防治效果較好。輪作后Fimbriimonas菌屬豐度較高,此菌具有固氮和脫氮特征,在維持氮平衡方面起重要作用[36]。
3.3 輪作時間與土壤細菌群落
細菌群落是土壤微生物中最主要的一類微生物,是土壤微生物多樣性的重要指標。應用高通量測序分析技術,可以比較準確地反映細菌群落特征[37- 38]。本研究通過高通量測序技術對比研究了不同輪作模式下土壤細菌多樣性,結果發現,不同的輪作模式下細菌群落結構具有一定的差異。但是主要優勢菌群保持穩定,都以變形菌門(Proteobacteria)豐度最高,土壤中每一特定性的菌類均具有不同的功能特性,變形菌是一類適應性很強的細菌,該類菌中即包含動植物的病原菌也存在抑制致病菌的有益菌[39- 40],但是不同輪作模式的細菌組成也有不同,輪作1年的比輪作2年的芽單胞菌門的豐度要高。芽單胞菌類是有益于作物生長的細菌,輪作1年時,該類菌群數量明顯高于第2年,這可以從土壤微生物特定菌類的增加來解釋輪作栽培有益于作物生物生長。輪作栽培由于不同的作物對養分和水分吸收以及根系分泌物不同,因而能夠改變土壤微生物的生態環境。吳宏亮對砂田土壤進行輪作栽培研究,發現輪作改變砂田土壤微生物區系結構,提高微生物多樣性[41]。本研究中,無論后茬作物玉米還是小麥,輪作2年實際上又造成了禾本科作物的連作,因此,其土壤微生態的組成要劣于輪作1年的。
4 結論
在多年種植苜蓿地和不同禾本科作物復種輪作模式下,不同輪作模式對土壤養分各指標有不同程度的影響,后茬作物無論是玉米還是小麥,輪組第2年土壤有機質、全氮、全磷、全鉀、堿解氮、速效磷和速效鉀的含量都較第1年有明顯的下降。4種輪作模式下占優勢的菌門為變形菌門、放線菌門和厚壁菌門。土壤細菌群落屬組成中以丙酸桿菌屬(Propionibacterium)、芽孢桿菌屬(Bacillus)、鏈球菌屬(Streptococcus)以及奈瑟氏球菌屬(Neisseria)為優勢屬等為主。土壤細菌多樣性以A5C2輪作模式下最高,A5C1最低;聚類分析顯示A5W1和A5C1的組成最接近,其次是A5C2,A5W2的組成與前兩者相差較大。土壤養分因子含量與幾種優勢屬呈現了不同程度的相關性。研究結果揭示了河西走廊灌溉區種植多年紫花苜蓿后復種小麥和玉米改善土壤肥力和防治某些土傳植物病害微生物的機理。
