騰格里沙漠東南緣藻結皮與蘚結皮放線菌多樣性及其潛在代謝功能
2020-04-22 01:38:10李靖宇張肖沖靳新影馬志山蘇亮文
生態學報 2020年5期
李靖宇,張肖沖,陳 韻,靳新影,馬志山,金 多,蘇亮文
北方民族大學生物科學與工程學院, 銀川 750021
生物土壤結皮(Biological Soil Crusts,BSCs)廣泛分布于全球干旱、半干旱地區,主要由藍藻、綠藻、地衣、蘚類和微生物以及其他生物體通過菌絲體、假根和分泌物等與土壤表層顆粒膠結形成十分復雜的地表覆蓋體,在防風固沙,改善表層土壤生態功能方面發揮著重要作用[1- 3]。土壤微生物作為BSCs的重要組成部分,可通過分解植物殘體并將其轉化成土壤有機質和土壤腐殖質來改善土壤質量,通過胞外多糖、金屬螯合劑及其分泌的肽將鈉、鉀、鎂、鈣、錳、鐵、鎳、銅和鋅等土壤養分進行保持和濃縮,造成BSCs和BSCs下層化學性質的巨大差異[4-5]。自養和異養微生物參與甚至主導BSCs中的生物地球化學循環和能量流動[6]。不同發育演替階段BSCs的微生物組成差異會導致固碳、固氮效率等方面存在明顯不同[6- 8]。Liu等[9]研究了不同恢復年限的BSCs對騰格里沙漠土壤微生物生物量的影響,結果表明人工植被固沙區不同類型BSCs均可顯著提高土壤微生物生物量碳和氮,隨著固沙年限的推移微生物生物量碳和氮含量也在增加,表明BSCs的定殖和發展有利于土壤微生物特性和質量的改善[10]。
放線菌廣泛分布于不同生態系統中,其中干旱生境是發現放線菌新菌種最豐富的生態系統之一[11]。宏基因組學研究結果表明騰格里沙漠東南緣蘚結皮土壤微生物組主要類群為放線菌[12]。BSCs作為沙漠極端環境中的生命存在形式之一,可以通過生物化學反應路徑固定大氣中的氮和二氧化碳等物質,進而推動形成復雜的微生物組[12]。微生物組的每一個成員都是復雜代謝網絡的載體,輸入到其中的物質與能量經過代謝網絡的轉化形成復雜的代謝物庫。放線菌是臨床有用天然產物化學多樣性的最重要來源之一,也是新化學結構發掘的重要寶庫[13]。在自然環境中,放線菌生活的環境條件和生態相互作用可能影響天然產物的生物合成[14]。目前,我們對這些環境條件的理解以及天然產物的生態作用尚處于起步階段[14]。棲息在沙漠中的放線菌能夠在極度干燥的條件下生長,并且包含著能夠合成獨特抗菌活性化合物的基因簇[11]。通過對文獻數據的分析,Mohammadipanah等人提出了開發適應干旱環境的放線菌以及探索干旱生態系統放線菌分布的必要性[11]。Sun等[15]人通過高通量測序研究表明,放線菌是巴丹吉林沙漠和騰格里沙漠主要的微生物類群,在屬水平上Arthrobacterspp.和Kocuriaspp.是騰格里沙漠的優勢類群;通過分離培養的方法發現Streptomycesspp.是騰格里沙漠可培養放線菌的主要類群。
基于已報道文獻,關于騰格里沙漠東南緣廣泛分布的藻結皮與蘚結皮的放線菌群落組成及其潛在代謝功能的比較研究還較少。本文利用宏基因組學技術通過對藻結皮和蘚結皮土壤微生物組的測序,較為深入的分析了兩種不同類型的BSCs中放線菌種群的分布特征、組成及其潛在代謝功能。可為開發利用極端環境條件下放線菌種資源、挖掘特定放線菌株的基因資源提供重要線索,也為深入評估放線菌資源的生物學特性、潛在功能和應用奠定基礎。
1 材料與方法
1.1 研究區域與樣品采集
研究區域設在寧夏回族自治區中衛市沙坡頭地區騰格里沙漠東南緣。該地區年均氣溫10.0℃,低溫極值-25.1℃,高溫極值38.1℃,全年日照時數3264 h,年均降水量186.2 mm(1956—2012年),年潛在蒸發量3000 mm,年均風速2.9 m/s,年均沙暴天數59 d[16]。1956年,為確保包頭-蘭州鐵路順利通過騰格里沙漠東南緣,通過設置沙障以及草方格,在此建立了長16 km,寬0.7 km(鐵路北側約0.5 km,南側約0.2 km)的植被保護帶。隨著沙子表面穩定后,在草方格中種植旱生灌木,如油蒿(Artemisiaordosica)、檸條錦雞兒(Caraganakoshinskii)和細枝巖黃芪(Hedysarumscoparium)等。到目前為止,該地區的局部環境得到改善,大量的BSCs覆蓋了80%以上的土壤表面,包括藍藻結皮約占10%,地衣結皮約占50%—60%以及蘚結皮約占20%—30%[17]。

圖1 采樣點位置圖Fig.1 Sampling sites X1—X6代表蘚結皮土壤樣品,X1—X6 represent the moss crust samples;Z1—Z6代表藻結皮土壤樣品,Z1—Z6 represent the cyanobacteria crust samples
取樣之前對騰格里沙漠東南緣進行現場考察后結合衛星圖像,實驗預設了藻結皮6個重復,蘚結皮6個重復,并按照空間交叉布局,即沿著樣帶按照藻結皮-蘚結皮交替預設取樣點,相鄰兩樣點之間的距離大約為1 km。2017年9月17日選取沙坡頭地區BSCs形成帶藻結皮與蘚結皮預設樣點附近發育良好的位點進行取樣,每個樣點按照五點取樣法進行取樣,取樣深度為各種結皮形成的自然厚度(藻結皮厚度范圍約1—6 mm,蘚結皮厚度范圍約11—22 mm),將土樣帶回實驗室,過2 mm篩子,將處理后的12個樣品冷凍保存,用于后續宏基因組DNA提取分析。藻結皮樣品總有機碳含量范圍為4.42—7.90 g/kg,總氮含量范圍為0.31—0.71 g/kg,總磷含量范圍為0.20—0.32 g/kg,總鉀含量范圍為13.58—15.22 g/kg;蘚結皮樣品總有機碳含量范圍為8.17—11.91 g/kg,總氮含量范圍為0.57—0.77 g/kg,總磷含量范圍為0.25—0.43 g/kg,總鉀含量范圍為12.99—15.35 g/kg。取樣點見圖1。
1.2 土壤宏基因組DNA提取和Illumina高通量測序
按照FastDNA Spin Kit for Soil (MP Biomedicals, Santa Ana, CA, USA)提取試劑盒說明書提取土壤宏基因組DNA,利用1%瓊脂糖凝膠電泳檢測抽提的基因組DNA。將純化后的宏基因組DNA破碎成片段,按照TruSeqTMDNA Sample Prep Kit試劑盒說明書構建PE(paired-end)文庫。然后根據HiSeq 3000/4000 PE Cluster Kit試劑盒說明書進行橋式PCR擴增。隨后按照Illumina公司提供的標準實驗流程,通過HiSeq 2000高通量測序平臺(Illumina)進行測序。
1.3 放線菌多樣性及其功能分析
首先對原始測序數據進行質量控制,使用軟件Seqprep(https://github.com/jstjohn/SeqPrep)剪切序列3′端和5′端的adapter序列,使用軟件Sickle(https://github.com/najoshi/sickle)去除質量剪切后長度小于50 bp、平均質量值低于20以及含N堿基的reads,保留高質量的pair-end reads。質控后得到的短片段序列,利用IDBA-UD(http://i.cs.hku.hk/~alse/hkubrg/projects/idba_ud/)軟件針對不同測序深度短reads進行從頭拼接組裝,從小k-mer開始到大k-mer不斷迭代處理,每次迭代根據設定閾值從低到高依次刪掉短的和低深度的contigs,逐步完成低深度和高深度短reads的拼接組裝。使用MetaGene (http://metagene.cb.k.u-tokyo.ac.jp/)對拼接結果中的contigs進行ORFs(open reading frame)預測。將所有樣品預測出來的基因序列,用CD-HIT軟件(http://www.bioinformatics.org/cd-hit/)進行聚類(默認參數為:95% identity、90% coverage),每個類取最長的基因作為代表序列,構建非冗余基因集。使用SOAPaligner 軟件(http://soap.genomics.org.cn/),分別將每個樣品的高質量reads與非冗余基因集進行比對(默認參數為:95% identity),統計基因在對應樣品中的豐度信息。使用BLASTP(BLAST Version 2.2.28+,http://blast.ncbi.nlm.nih.gov/Blast.cgi)將非冗余基因集與NR數據庫進行比對(BLAST 比對參數設置期望值e-value為1e-5),并通過NR庫對應的分類學信息數據庫獲得物種注釋結果。使用BLASTP(BLAST Version 2.2.28+,http://blast.ncbi.nlm.nih.gov/Blast.cgi)將非冗余基因集序列與KEGG的基因數據庫(GENES)進行比對,BLAST 比對參數設置期望值e-value為1e-5。根據比對結果使用KOBAS 2.0(KEGG Orthology Based Annotation System)進行功能注釋。使用KO、Pathway、EC、Module 對應的基因豐度總和計算該功能類別的豐度。最后通過上海美吉生物醫藥科技有限公司開發的I-Sanger云平臺(www.i-sanger.com)挑選放線菌的物種注釋信息和KEGG注釋信息,并進行Circos作圖分析、PCA分析(Principal Component Analysis)、相似性分析(ANOSIM)、組間顯著性差異檢驗分析以及物種與功能貢獻度分析。
2 結果與分析
2.1 藻結皮與蘚結皮放線菌群落組成
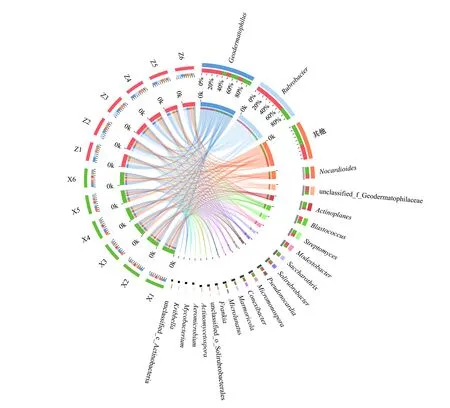
騰格里沙漠東南緣藻結皮與蘚結皮土壤微生物組主要形成以地嗜皮菌屬(Geodermatophilus)、紅色桿菌屬(Rubrobacter)、類諾卡氏菌屬(Nocardioides)、游動放線菌屬(Actinoplanes)、芽生球菌屬(Blastococcus)、鏈霉菌屬(Streptomyces)、貧養桿菌屬(Modestobacter)、糖絲菌屬(Saccharothrix)、土壤紅桿菌屬(Solirubrobacter)、假諾卡氏菌屬(Pseudonocardia)、小單孢子菌屬(Micromonospora)、康奈斯氏桿菌屬(Conexibacter)、大理石雕菌屬(Marmoricola)、小月菌屬(Microlunatus)以及弗蘭克氏菌屬(Frankia)等為主要類群的放線菌群落結構特征(圖2)。主成分分析結果表明主成分1可以解釋藻結皮與蘚結皮放線菌群落差異的36.01%,主成分2可以解釋27.86%,主成分3可以解釋9.45%。ANOSIM分析結果表明放線菌群落在藻結皮與蘚結皮之間存在顯著差異(圖2)。

圖2 藻結皮與蘚結皮放線菌門各屬的Circos圖, 主成分分析, ANOSIM比較分析和顯著性差異分析Fig.2 Circos diagram, principal component analysis, ANOSIM comparative analysis and significant difference analysis for different genera in Actinobacteria phylum between cyanobacteria crust and moss crustGeodermatophilus地嗜皮菌屬;Rubrobacter 紅色桿菌屬;Nocardioides類諾卡氏菌屬;unclassified_f_Geodermatophilaceae 未分類的地嗜皮菌科屬;Actinoplanes 游動放線菌屬;Blastococcus芽生球菌屬;Streptomyces 鏈霉菌屬;Modestobacter貧養桿菌屬;Saccharothrix 糖絲菌屬;Solirubrobacter 土壤紅桿菌屬;Pseudonocardia 假諾卡氏菌屬;Micromonospora 小單孢子菌屬;Conexibacter 康奈斯氏桿菌屬;Marmoricola 大理石雕菌屬;Microlunatus小月菌屬;Frankia弗蘭克氏菌屬;unclassified_o__Solirubrobacterales 未分類的土壤紅桿菌屬;Actinomycetospora 放線產孢菌屬;Aeromicrobium氣微菌屬;Mycobacterium 分支桿菌;Kribbella 韓國生工菌屬
地嗜皮菌屬作為該地區藻結皮與蘚結皮的優勢放線菌類群之一,在藻結皮中其相對豐度達到26.60%,而蘚結皮中其相對豐度為20.41%,二者之間存在顯著差異。紅色桿菌屬是藻結皮中相對豐度最高的放線菌類群,其值達到27.55%;而該類群在蘚結皮中的相對豐度為10.94%,顯著低于藻結皮中的相對豐度。類諾卡氏菌屬在蘚結皮(7.61%)中的相對豐度要顯著高于藻結皮(4.23%);游動放線菌屬在蘚結皮(5.81%)中的相對豐度要顯著高于藻結皮(1.42%);鏈霉菌屬在蘚結皮(3.66%)中的相對豐度要顯著高于藻結皮(2.87%);糖絲菌屬在蘚結皮(3.73%)中的相對豐度要顯著高于藻結皮(0.93%);小單孢子菌屬在蘚結皮(2.89%)中的相對豐度要顯著高于藻結皮(0.92%);大理石雕菌屬在蘚結皮(2.43%)中的相對豐度要顯著高于藻結皮(1.21%);土壤紅桿菌屬在蘚結皮(2.04%)中的相對豐度要顯著高于藻結皮(1.56%);小月菌屬在蘚結皮(2.96%)中的相對豐度要顯著高于藻結皮(0.47%)(圖2)。
2.2 藻結皮與蘚結皮放線菌群落代謝特征
基于KEGG數據庫對騰格里沙漠東南緣藻結皮與蘚結皮土壤微生物組中放線菌群落的宏基因組數據進行了代謝功能注釋,結果表明相對豐度平均值(n=12)高于1%的代謝功能分類主要包括碳代謝(6.11%)、氨基酸生物合成(5.21%)、氧化磷酸化(4.32%)、嘌呤代謝(3.50%)、嘧啶代謝(3.60%)、氨基糖與核苷酸糖代謝(2.49%)、糖酵解/糖異生(2.04%)、原核生物中的碳固定途徑(2.07%)、丙酮酸代謝(1.93%)、三羧酸循環(1.80%)、脂肪酸代謝(1.89%)、甘氨酸/絲氨酸和蘇氨酸代謝(1.87%)、丙酸代謝(1.48%)、乙醛酸和二羧酸鹽代謝(1.67%)、肽聚糖生物合成(1.27%)、丙氨酸/天門冬氨酸和谷氨酸代謝(1.46%)、丁酸代謝(1.51%)、淀粉與蔗糖代謝(1.18%)、甲烷代謝(1.35%)、2-羰基羧酸代謝(1.28%)、果糖與甘露糖代謝(1.52%)、磷酸戊糖途徑(1.48%)、半乳糖代謝(1.37%)、半胱氨酸和蛋氨酸代謝(1.45%)、脂肪酸生物合成(1.44%)、纈氨酸/亮氨酸與異亮氨酸降解(1.06%)、精氨酸與脯氨酸代謝(1.07%)、脂多糖生物合成(1.08%)、甘油磷脂代謝(1.08%)和葉酸合成(1.02%)等(圖3)。
相對豐度前5的代謝功能,包括碳代謝、氨基酸生物合成、氧化磷酸化、嘌呤代謝以及嘧啶代謝,在藻結皮和蘚結皮之間沒有顯著差異。氨基糖與核苷酸糖代謝、原核生物中的碳固定途徑、丁酸代謝、丙酸代謝、丙氨酸/天門冬氨酸和谷氨酸代謝、甲烷代謝、2-羰基羧酸代謝、肽聚糖生物合成、淀粉與蔗糖代謝以及纈氨酸/亮氨酸與異亮氨酸降解在蘚結皮中顯著高于藻結皮(圖3)。


圖3 藻結皮與蘚結皮放線菌門各屬代謝功能分類Circos圖和組間顯著性差異分析Fig.3 Circos diagram and significant difference analysis for metabolism functional classification of different genera in Actinobacteria phylum between cyanobacteria crust and moss crustCarbon metabolism碳代謝;Biosynthesis of amino acids氨基酸生物合成;Oxidative phosphorylation 氧化磷酸化;Purine metabolism嘌呤代謝;Pyrimidine metabolism嘧啶代謝;Amino sugar and nucleotide sugar metabolism氨基糖與核苷酸糖代謝;Glycolysis/Gluconeogenesis糖酵解/糖異生;Carbon fixation pathways in prokaryote原核生物中的碳固定途徑;Pyruvate metabolism丙酮酸代謝;Fatty acid metabolism 脂肪酸代謝;Glycine, serine and threonine metabolism甘氨酸、絲氨酸和蘇氨酸代謝;Citrate cycle (TCA cycle) 三羧酸循環;Glyoxylate and dicarboxylate metabolism乙醛酸和二羧酸鹽代謝;Fructose and mannose metabolism果糖與甘露糖代謝;Butanoate metabolism丁酸代謝;Propanoate metabolism丙酸代謝;Pentose phosphate pathway磷酸戊糖途徑;Alanine, aspartate and glutamate metabolism丙氨酸、天門冬氨酸和谷氨酸代謝;Cysteine and methionine metabolism半胱氨酸和蛋氨酸代謝;Fatty acid biosynthesis 脂肪酸生物合成;Galactose metabolism半乳糖代謝;Methane metabolism 甲烷代謝;2-Oxocarboxylic acid metabolism 2-羰基羧酸代謝;Peptidoglycan biosynthesis肽聚糖生物合成;Starch and sucrose metabolism淀粉與蔗糖代謝;Lipopolysaccharide biosynthesis脂多糖生物合成;Glycerophospholipid metabolism甘油磷脂代謝;Arginine and proline metabolism精氨酸與脯氨酸代謝;Valine, leucine and isoleucine degradation纈氨酸、亮氨酸與異亮氨酸降解;Folate biosynthesis葉酸合成;Carbon fixation in photosynthetic organisms光合生物中的固碳;Nicotinate and nicotinamide metabolism煙酸和煙酰胺代謝;Ubiquinone and other terpenoid-quinone biosynthesis泛醌及其他萜類醌生物合成;Arginine biosynthesis精氨酸生物合成;Photosynthesis光合作用;Biotin metabolism生物素代謝;Tryptophan metabolism色氨酸代謝;Lysine biosynthesis賴氨酸生物合成;Valine, leucine and isoleucine biosynthesis纈氨酸、亮氨酸與異亮氨酸生物合成;Pentose and glucuronate interconversions戊糖和葡糖醛酸鹽相互轉化;Histidine metabolism組氨酸代謝
2.3 藻結皮與蘚結皮放線菌群落KEGG模塊功能分析
KEGG模塊是手動定義的功能單元的集合,用于測序基因組的注釋和生物學解釋。基于KEGG數據庫對騰格里沙漠東南緣藻結皮與蘚結皮土壤微生物組中放線菌群落的宏基因組數據進行了模塊功能注釋,結果表明細菌核糖體模塊的相對豐度最高,蘚結皮中相對豐度為14.63%,藻結皮中相對豐度為10.51%,且兩種結皮類型存在顯著差異。其次豐度最大的為原核生物NADH-醌氧化還原酶功能模塊,蘚結皮中相對豐度為4.70%,藻結皮中相對豐度為4.44%,兩種結皮類型之間不存在顯著差異。相對豐度平均值(n=12)高于1%的其他功能模塊主要包括三羧酸循環、氨酰-tRNA生物合成(真核生物)、ABC轉運系統、ABC- 2型轉運系統、肌苷一磷酸生物合成、F型ATP酶(原核生物和葉綠體)、細胞色素C氧化酶(原核生物)、脂肪酸生物合成(延伸)、脂多糖轉運系統、尿苷一磷酸生物合成、核黃素生物合成、磷酸戊糖途徑、賴氨酸生物合成、分泌系統、磷酸鹽饑餓反應雙組分調節系統、糖酵解、脂多糖生物合成、肽/鎳轉運系統、煙酰胺腺嘌呤二核苷酸生物合成、異亮氨酸生物合成、三酰甘油生物合成、dTDP-L-鼠李糖生物合成、Ⅱ型分泌途徑、RNA聚合酶(細菌)、DNA聚合酶Ⅲ復合物(細菌)等。其中,三羧酸循環、氨酰-tRNA生物合成(真核生物)、ABC- 2型轉運系統 、dTDP-L-鼠李糖生物合成以及DNA聚合酶Ⅲ復合物(細菌) 等功能模塊在蘚結皮中顯著高于藻結皮(圖4)。
2.4 藻結皮與蘚結皮放線菌門優勢屬與功能貢獻度分析
基于樣本中物種和功能的對應關系,進行物種與功能相對豐度之間的關聯分析,找出特定物種的功能貢獻度。選擇了物種相對豐度前10和功能豐度前10的KEGG功能分類水平- 3進行了物種和功能對應分析。結果表明,在碳代謝功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到61.45%,而在蘚結皮中為38.69%;在氨基酸生物合成功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到61.37%,而在蘚結皮中為36.43%;在ABC轉運體功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到46.36%,而在蘚結皮中為15.38%;在嘌呤代謝功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到60.51%,而在蘚結皮中為36.79%;在群體感應功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到51.11%,而在蘚結皮中為29.92%;在嘧啶代謝功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到60.67%,而在蘚結皮中為37.11%;在氧化磷酸化功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到56.71%,而在蘚結皮中為33.74%;在乙醛酸和二羧酸鹽代謝功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到60.78%,而在蘚結皮中為38.01%;在丙酮酸代謝功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到61.08%,而在蘚結皮中為40.14%;在原核生物的碳固定途徑功能分類中,地嗜皮菌屬和紅色桿菌屬的相對貢獻在藻結皮中達到61.04%,而在蘚結皮中為39.15%(圖5)。從藻結皮和蘚結皮這兩種類型來看,地嗜皮菌屬和紅色桿菌屬對以上10中功能分類的貢獻度在蘚結皮中顯著低于藻結皮,而在蘚結皮中,類諾卡氏菌屬、芽生球菌屬、貧養桿菌屬、游動放線菌屬、鏈霉菌屬、假諾卡氏菌屬和糖絲菌屬等對這些功能的相對貢獻具有重要作用(見圖5)。
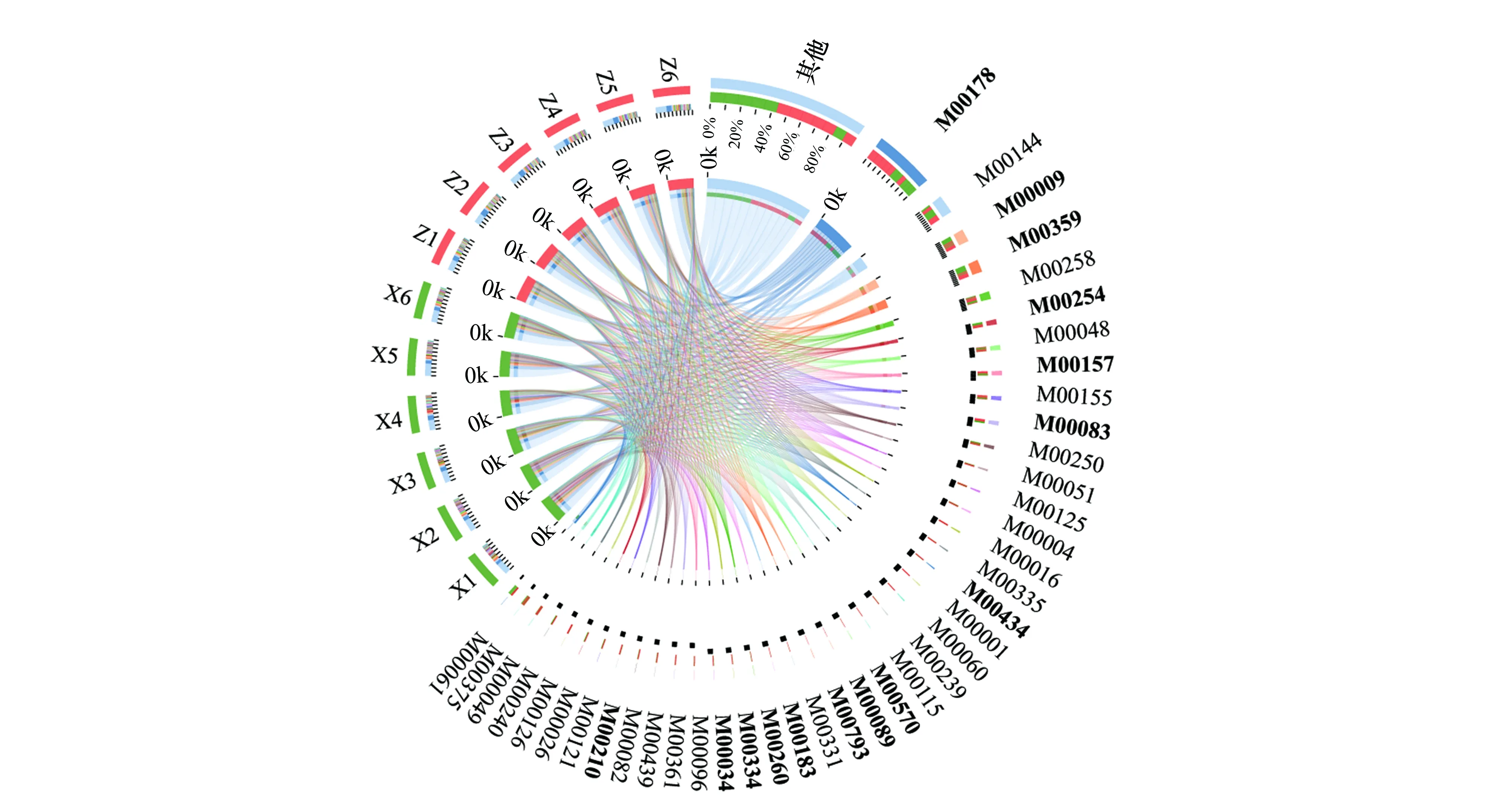
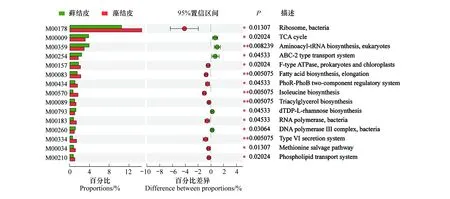
圖4 藻結皮與蘚結皮放線菌門各屬的Module功能分類Circos圖和組間顯著性差異分析Fig.4 Circos diagram and significant difference analysis for Module functional classification of different genera in Actinobacteria phylum between cyanobacteria crust and moss crustM00178:核糖體(細菌) Ribosome(bacteria);M00144:NADH:醌氧化還原酶(原核生物) NADH:quinone oxidoreductase(prokaryotes); M00009:三羧酸循環 Citrate cycle;M00359:氨酰-tRNA生物合成(真核生物) Aminoacyl-tRNA biosynthesis(eukaryotes);M00258:公認的ABC轉運系統 Putative ABC transport system;M00254:ABC- 2型轉運系統 ABC- 2 type transport system;M00048:肌苷一磷酸生物合成Inosine monophosphate biosynthesis;M00157:F型ATP酶(原核生物和葉綠體) F-type ATPase(prokaryotes and chloroplasts);M00155:細胞色素C氧化酶(原核生物) Cytochrome c oxidase(prokaryotes);M00083:脂肪酸生物合成(延伸) Fatty acid biosynthesis(elongation);M00250:脂多糖轉運系統Lipopolysaccharide transport system;M00051:尿苷一磷酸生物合成Uridine monophosphate biosynthesis;M00125:核黃素生物合成Riboflavin biosynthesis;M00004:磷酸戊糖途徑Pentose phosphate pathway;M00016:賴氨酸生物合成Lysine biosynthesis;M00335:分泌系統secretion system;M00434:磷酸鹽饑餓反應雙組分調節系統 PhoR-PhoB two-component regulatory system;M00001:糖酵解 Glycolysis;M00060:脂多糖生物合成Lipopolysaccharide biosynthesis;M00239:肽/鎳轉運系統Peptides/nickel transport system;M00115:煙酰胺腺嘌呤二核苷酸生物合成NAD biosynthesis;M00570:異亮氨酸生物合成Isoleucine biosynthesis;M00089:三酰甘油生物合成Triacylglycerol biosynthesis;M00793:dTDP-L-鼠李糖生物合成dTDP-L-rhamnose biosynthesis;M00331:Ⅱ型分泌途徑Type II general secretion pathway;M00183:RNA聚合酶(細菌) RNA polymerase(bacteria);M00260:DNA聚合酶Ⅲ復合物(細菌) DNA polymerase III complex(bacteria);M00334:Ⅵ型分泌系統Type VI secretion system;M00096:C5類異戊二烯生物合成C5 isoprenoid biosynthesis;M00361:核苷酸糖生物合成(真核生物) Nucleotide sugar biosynthesis(eukaryotes);M00439:寡肽轉運系統 Oligopeptide transport system;M00082:脂肪酸生物合成(起始) Fatty acid biosynthesis(initiation);M00210:磷脂轉運系統Phospholipid transport system;M00121:血紅素生物合成Heme biosynthesis;M00026:組氨酸生物合成Histidine biosynthesis;M00126:四氫葉酸生物合成Tetrahydrofolate biosynthesis;M00240:鐵配合物轉運系統Iron complex transport system;M00049:腺嘌呤核糖核酸生物合成Adenine ribonucleotide biosynthesis;M00375:羥基丙酸酯-羥基丁酸酯循環 Hydroxypropionate-hydroxybutylate cycle;M00061:葡萄糖醛酸降解D-Glucuronate degradation
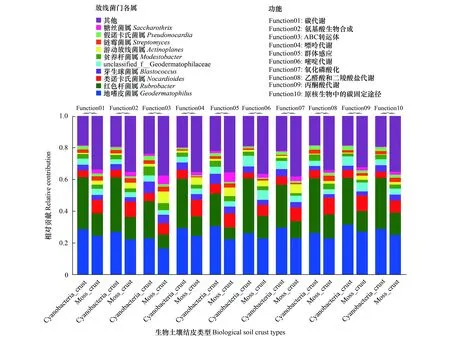
圖5 藻結皮與蘚結皮放線菌門各屬的物種與功能貢獻度分析Fig.5 Species and functional contribution analysis for different genera in Actinobacteria phylum between cyanobacteria crust and moss crust
3 討論
放線菌多樣性是騰格里沙漠東南緣藻結皮和蘚結皮土壤微生物組的重要組成部分,是BSCs進行生物地球化學循環的重要參與者。放線菌各屬在藻結皮和蘚結皮中的分布情況對于全面、深入認識BSCs的發育、演替提供了物種多樣性數據。物種多樣性以生命的形式將眾多指導生物化學反應的基因多樣性整合在一起,構建起了復雜的代謝網絡,為放線菌適應極端環境、推動物質循環和能量流動提供了保障,而這些復雜的代謝網絡成為了臨床有用天然產物化學多樣性挖掘、開發、利用的潛在重要“生產線”。這可能有助于對BSCs中放線菌生態功能的深入理解和認識。從藻結皮到蘚結皮的演替過程中,放線菌多樣性及其群落結構變化明顯,優勢類群差異明顯,特別是在碳代謝等具體生態功能方面,優勢類群地嗜皮菌屬和紅色桿菌屬的貢獻較大,這對于深入理解放線菌在生物土壤結皮演替過程中的生態功能具有重要的科學意義。
地嗜皮菌屬作為藻結皮和蘚結皮的優勢屬之一。該屬微生物廣泛分布于干旱極端環境條件下,對于能夠產生活性氧的輻射、紫外線以及干燥等環境脅迫具有很強的抗性[18]。通過比較發現,藻結皮土壤微生物組中地嗜皮菌屬相對豐度顯著高于蘚結皮。隨著BSCs從藻結皮向蘚結皮演替的過程中,放線菌的優勢屬發生了顯著變化,局部環境的改善減少了地嗜皮菌屬的相對豐度。這也間接說明了地嗜皮菌屬在藻結皮適應極端干旱環境過程中具有更加重要的生態功能。同屬于地嗜皮菌科的屬主要包括地嗜皮菌屬(模式屬)、芽生球菌屬和貧養桿菌屬,這三個屬都呈現出藻結皮土壤微生物組中的相對豐度顯著高于蘚結皮。地嗜皮菌科不僅與石灰巖退化過程密切相關,甚至能夠腐蝕巖石,而且對干旱、高溫、離子輻射和低營養等惡劣環境具有較強的耐受性[19]。地嗜皮菌屬的模式種GeodermatophilusobscurusLuedemann 1968全基因組測序以及生理特性研究結果表明,其基因組(G+C) 含量為74%,基因多拷貝,可大量產生細胞外功能性細菌淀粉樣蛋白,菌落呈現黑色[20]。有研究者從突尼斯和埃及的沙漠石頭和紀念碑上分離到地嗜皮菌屬菌株12株、芽生球菌屬菌株5株以及貧養桿菌屬菌株3株,對這些菌株的酯酶酶學性質研究表明,地嗜皮菌科具有豐富且獨特的酯酶活性,這些酯酶的活性與極度干旱、高溫、強電離輻射及營養供給匱乏等為特征的嚴酷生態環境直接相關[21]。
紅色桿菌屬作為藻結皮相對豐度最大的類群,在蘚結皮中相對豐度顯著下降。紅桿菌屬最早由Suzuki等人于1988年建立,他們從日本的一個放射性溫泉中分離出一種高度抗伽馬輻射的細菌,即Rubrobacterradiotolerans[22]。從此以后,科學家們相繼從工業排污熱水、溫泉、地熱流以及火山口周圍土壤等高溫環境或非高溫環境(如教堂墻壁生物膜、海綿)中分離出嗜熱、耐鹽或對γ射線具有高度抗性的紅桿菌屬菌株,命名為Rubrobacterxylanophilus、Rubrobactertaiwanensis、Rubrobactercalidiflumis、Rubrobacternaidicus、Rubrobacterbracarensis、Rubrobacteraplysinae、Rubrobacterbracarensis[23]。紅色桿菌屬放線菌廣泛分布于阿塔卡馬沙漠、哈薩克斯坦極端干旱沙漠土壤、澳大利亞干旱土壤、中國騰格里沙漠以及巴丹吉林沙漠等干旱環境中[15,24- 26]。Rubrobacterradiotolerans菌株 RSPS- 4對電離輻射有很強的抵抗力,全基因組測序結果表明其基因組(G+C) 含量為66.91%,含有3個環狀質粒,分別為190889 bp,149806 bp和51047 bp,含有3214個預測的蛋白質編碼基因,46個tRNA基因和一個rRNA操縱子,該基因組擁有參與DNA修復的RecFOR途徑的所有基因[27]。基于以上分析,騰格里沙漠東南緣藻結皮和蘚結皮中廣泛分布的紅色桿菌屬對于適應沙漠干旱、強輻射等極端環境具有重要作用,在以后的研究中需要加強對該類微生物的分離、純化,獲得純培養,以便進一步深入探究該類微生物適應極端環境的分子機理。
隨著BSCs的演替,蘚結皮土壤微生物組中類諾卡氏菌屬、游動放線菌屬、鏈霉菌屬、糖絲菌屬、小單孢子菌屬、大理石雕菌屬、土壤紅桿菌屬、小月菌屬、康奈斯氏桿菌屬相對豐度顯著高于藻結皮。類諾卡氏菌屬成員廣泛分布于土壤、海水、火山灰、海膽、沙漠、地下水、沙灘、冰川、山地土、森林土、沉積物、藥用植物(麻瘋樹屬)以及銀杏和玉米的根部等環境中[28]。游動放線菌屬具有球形、圓柱形、指狀、淺裂、瓶狀或燒瓶狀或非常不規則的孢子囊特征,其成員已逐漸被認為是次生代謝產物的重要來源[29]。鏈霉菌屬普遍存在于土壤中,它們可以形成耐干燥和饑餓的孢子,可以降解纖維素和幾丁質等復雜有機分子,在陸地碳循環中起著重要作用[30]。該屬最顯著的特性是其產生次生代謝物的能力,其中許多具有重要的工業和藥用價值,這些次生代謝物之間還存在著相當大的結構多樣性,能夠大量產生微生物來源的抗生素、抗真菌劑以及腫瘤抑制化合物[31]。當饑餓或環境變化導致營養生長結束時,伴隨著基因表達的一系列復雜變化,抗生素的產生通常會被激活,導致次生代謝產物的產生[32]。糖絲菌屬的成員主要來源于土壤,包括沙漠土壤,是新抗生素的潛在生產者,因能夠產生對細菌和真菌具有活性的新型特殊代謝物而備受關注[33-34]。小單孢子菌屬的成員主要來源于土壤、昆蟲、海洋沉積物、植物、固氮根瘤、葉和根等環境中,該屬逐漸被認為是次生代謝產物的重要來源,已從該屬中分離出許多抗生素[35-36]。小單孢菌屬中的內生放線菌作為潛在拮抗劑的來源,為發現新型生物活性天然產物帶來了巨大希望[36]。
4 結論
騰格里沙漠東南緣藻結皮和蘚結皮土壤微生物組中放線菌多樣性高,是兩種BSCs的優勢類群。主要形成以地嗜皮菌屬和紅色桿菌屬為主要類群的放線菌群落結構特征。其中,藻結皮中以紅色桿菌屬為優勢類群,而在蘚結皮中該類群放線菌相對豐度顯著下降;而蘚結皮以地嗜皮菌屬為優勢類群,而在藻結皮中該類群放線菌顯著下降。在藻結皮向蘚結皮演替的過程中,放線菌群落結構發生了明顯的變化。特別是在蘚結皮中,一些具有潛在產生重要次生代謝產物的放線菌類群的相對豐度顯著高于藻結皮,這可能會為開發、利用放線菌資源多樣性提供重要參考。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
