膜電位的產生和傳導常見誤區辨析
2020-04-22 20:41:22謝仁榮申定健吳志強房杰
中學生物學 2020年1期
謝仁榮 申定健 吳志強 房杰

摘要 細胞膜電位主要由膜內外K+、Na+濃度梯度和膜對Na+和K+的通透性共同決定,靜息電位并不等同于K+平衡電位,動作電位是膜電位的變化過程,動作電位的傳導是局部電流刺激未興奮部分的Na+通道打開,引起該處產生同樣的動作電位。進一步分析了膜外K+、Na+濃度對靜息電位和動作電位的影響。
關鍵詞:鈉鉀泵,靜息電位,動作電位,膜電導
中圖分類號G633.91文獻標志碼B
人教版生物學教科書《必修3·穩態與環境》中對靜息電位的形成及興奮的產生和傳導過程介紹較為簡略,也沒有介紹動作電位,師生常會產生誤解,如把靜息電位直接等同于K+平衡電位,動作電位等同于Na+平衡電位,誤認為動作電位就是以局部電流的形式進行傳導,亦難以理解K+、Na+濃度對靜息電位和動作電位大小的影響。下面簡要介紹膜電位的形成和變化,分析動作電位的傳導原理及K+、Na+濃度對靜息電位和動作電位峰值的影響。
1 靜息電位是否等同于K+平衡電位
1.1 鈉鉀泵的生電效應對靜息電位的直接貢獻
鈉鉀泵普遍分布于哺乳動物細胞膜上,是具有ATP酶活性,能夠主動運輸Na+和K+的蛋白質載體。鈉鉀泵每消耗1個ATP,可以運出3個Na+,運進2個K+,從而膜內積累了高濃度的K+,并保持著很低的Na+濃度,并使細胞外積累較多的陽離子。因此,細胞膜上的鈉鉀泵具有使膜外積累正電荷的生電效應。但這不是靜息電位形成的主要原因,靜息電位的形成主要來自離子濃度差本身所儲存的電勢能量,鈉鉀泵對膜電位的直接貢獻大約為20%。當膜外K+濃度升高或膜內Na~濃度升高,會引起鈉鉀泵的活動加強,而膜內K+濃度的變化及膜外Na+濃度的變化則不會引起鈉鉀泵的活動強度變化。
1.2 鈉鉀泵活動形成的Na+、K+濃度差及膜對Na+和K+的通透性是決定靜息電位的主要因素
哺乳動物神經元細胞內、外液中主要離子的濃度和靜息狀態下膜對離子通透性見表1。由于鈉鉀泵的活動,神經細胞內K+濃度比膜外約高30倍,假設細胞膜只對K+具有通透性,K+將在它的濃度梯度作用下向膜外被動擴散(通過漏鉀通道),而細胞內液中的負離子不能出去,膜外將逐漸積累正電荷,建立起一個外高內低的電位差,而膜的電位梯度又驅使K+重新回到細胞,故K+的跨膜擴散受到濃度差和電位差兩個驅動力的作用,其代數和稱為電一化學驅動力。
隨著K+不斷向外擴散,其電位差的排斥力也越大,當二力達到平衡時,即電一化學驅動力為零時,K+的靜通量為零,此時的跨膜電位為K+平衡電位。利用Nernst公式可以計算出此時K+的平衡電位(&):Ek=RT/ZF(ln[K+]o/[K+]i)。
上述計算式中,Ek為K+的平衡電位,R為氣體常數,T為絕對溫度,Z為原子價,F為法拉第常數,[K+]o和[K+]i分別為K+在細胞外液和細胞內液中的濃度。以哺乳類動物神經元為例,若體溫為37℃,則EK=-90mV。同樣,可以利用Nernst公式計算出Na+平衡電位(ENa)約為+60mV,CF-平衡電位(Ecl)約為-70mV。這里的“+”“-”僅表示膜內電位與膜外電位的相對關系,而不具有代數意義。當膜電位由“-90mW”變為“-60mV”時,可以認為膜電位變小了,這與其數學數值上的意義增大正好相反。每種離子的跨膜滲透都是相互獨立的,這種現象又稱為離子運動的獨立性原則。
細胞膜在靜息狀態時主要對Na+和K+、Cl-具有通透性,而對Ca2+、Mg2+、A-幾乎是不通透的。Na+和K+濃度梯度是鈉鉀泵消耗ATP的主動轉運形成,而Cl-平衡電位雖然很接近于靜息電位,但膜對Cl-不存在原發性主動轉運,Cl-在膜的兩側的分布是被動的,是膜電位決定它在膜內的濃度,而Cl-濃度梯度并不能決定膜電位。故靜息電位主要是由K+和Na+的平衡電位Ek和ENa共同決定,其他離子如Ca2+、Cl-、Mg2+等在大多數細胞中對靜息電位并無直接貢獻。需要注意的是,靜息電位并不是Ek和ENa簡單的代數和,在槍烏賊巨大神經纖維中,實際測得的膜電位為“-77mV”,只比Nernst公式計算的Z真理論值“-90mV”略小,而遠遠偏離Na+的平衡電位“+60mV”,其原因是在靜息狀態下,細胞膜對K+的通透性是Na+的50~75倍,由于膜對K+的通透性遠遠大于對Na+的通透性,故Ek在決定靜息電位時的權重遠大于ENa的權重,靜息電位就非常接近于EK。
膜電位的計算公式為:膜電位(Em)=GkxEk(Gk+CNa)+GNaxENa/(Gk+CNa)。上述公式中,Gk為鉀電導,GNa為鈉電導。利用電壓鉗技術固定細胞膜兩側的電壓,可以測出跨膜離子流,根據歐姆定律,可以計算出細胞膜對帶電離子的電阻,膜電阻的倒數即為膜電導,因而膜電導反映了細胞膜對帶電離子的通透性,膜電導越大,膜對某種離子通透性越大。故影響靜息電位大小的因素除了鈉鉀泵的生電效應外,還有Ek和ENa、膜對K+和Na+的相對通透性(即鉀電導和鈉電導比值)。
綜上所述,靜息電位并不等同于K+平衡電位,只是通常比較接近K+平衡電位。當膜外K+濃度急性降低時,膜內外K+濃度梯度變大,鉀離子平衡電位(Ek)也會增大,靜息電位也相應增大;反之,當膜外K+濃度較高時,Ek減小,靜息電位也減小。由于靜息狀態時鈉電導遠遠小于鉀電導,ENa在決定靜息電位時的權重太小,當膜外Na+濃度變化時,靜息電位幾乎沒有變化。
由于靜息膜電位(-77mV)并沒有真正達到K+的平衡電位(-90mV),不足以完全阻止K+的外流;而這個外正內負的靜息電位和Na+的濃度差都促使Na+內流,因而在靜息狀態,K+會不斷的向外滲漏,而Na+會不斷的向內滲漏,同時需要鈉鉀泵不斷的消耗ATP,將膜內的Na+搬運出去,將膜外的K+搬運進來,維持細胞內鉀高鈉低的狀態。在靜息狀態時,鈉鉀泵需要消耗掉細胞20%~30%的能量。
2 動作電位是否等同于Na+平衡電位
2.1 動作電位的產生
以典型的哺乳動物神經細胞為例,當神經纖維受到一個較小的刺激(如一個電刺激),使細胞膜電位變小(去極化),部分對電壓敏感的鈉離子通道(電壓門控鈉離子通道)打開,Na+順濃度梯度內流。但是,同時由于膜電位的變小,K+外流也增強,抵消了少量Na+內流對膜電位的影響,使電位在產生一個小的波動后又重新回到靜息電位,這個小的電位波動,稱之為局部電位或分級電位。
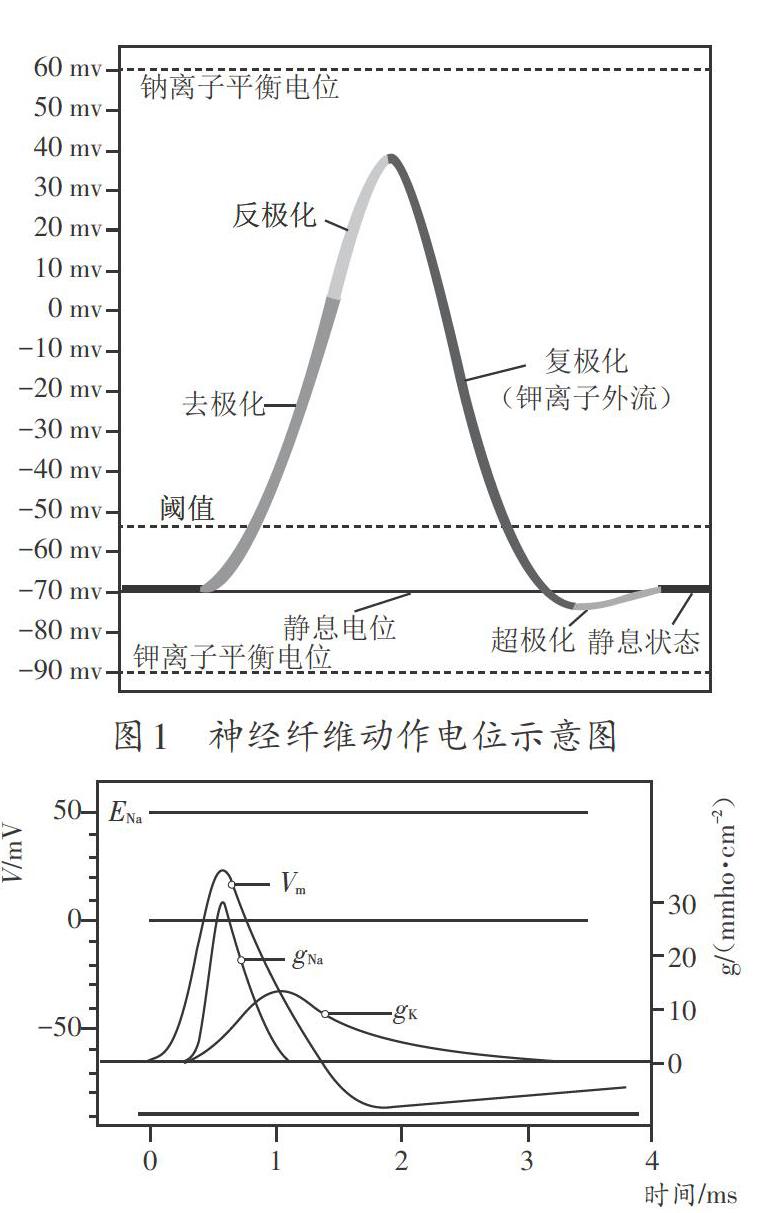
當刺激加大到一定程度(閾刺激),使膜電位減小到閾電位(如-55mV)以下,使較多的電壓門控Na+通道打開,Na+的內流超過因電位減小而增加的K+外流時,就會使膜電位進一步去極化,進而引起更多的鈉離子通道打開,形成一個正反饋,最終使所有的鈉離子通道都打開,使膜對Na+的通透性大大增大(GNa增大)。同時,電壓門控鉀離子通道也打開,膜對K+的通透性也增大(Gk增大),但是鈉離子通道打開的速度很快,而鉀離子通道打開較慢,有一定的延遲,因而GNa增大比Gk增大要快得多,使膜在0.5ms內對Na+的通透性就超過靜息時的500倍,內流的Na+遠超過外流的K+,這導致膜電位迅速反轉,變為內正外負,產生動作電位的上升支(圖1),膜電位快速向Na+的平衡電位移動,迅速到達一個+35mV左右的電位。但這個電位也未到達Na+的平衡電位(+60mV),這是因為在GNa增大的同時,Gk也增大了(圖2),但此時GNa要比Gk大得多,動作電位的峰值更偏向ENa。
若膜外的Na+濃度增大,將使細胞內外的Na+濃度差變大,ENa增大,動作電位的峰值也隨之更大;反之,降低膜外的Na+濃度,動作電位的峰值隨之變小。而由于動作電位上升支時GNa遠大于Gk,Ek對膜電位的影響很小,K+濃度變化對動作電位的峰值影響也很小,K+濃度主要影響靜息電位大小。因此,膜外鈉離子濃度對動作電位的幅度影響大,而膜外鉀離子濃度對動作電位影響很小。
2.2 動作電位的下降
鈉離子通道在打開后造成鈉離子大量內流后,迅速進入關閉狀態(0.5ms內),且暫時不能再打開(失活狀態)。隨著鈉離子通道的迅速關閉,GNa急劇減小,而鉀離子通道還在繼續打開,GK還在增加(圖2),進而K+外流超過少量內流的Na+,使膜電位又迅速向K+的平衡電位移動,構成動作電位的下降支(圖1)。隨即鉀離子通道逐漸關閉,GK逐漸變小。由于鉀離子門控通道關閉較慢,而漏鉀通道始終是開放的,膜對鉀離子的通透性(GK)還略大于靜息時期(僅通過漏鉀通道),有較多的K+外流,從而使膜電位更偏向于K+的平衡電位,產生一個比靜息電位更大的電位,即正后電位(圖1)。此外,鈉鉀泵的生電效應也對正后電位有一定貢獻。隨后,由于電壓門控鉀離子通道逐漸關閉,GK下降到靜息水平,膜電位也由正后電位逐漸被推到靜息電位。產生一次動作電位后,由于膜內Na+濃度及膜外K+濃度較靜息時有所上升,鈉鉀泵加強活動,將膜內多余的Na+的搬出細胞,將膜外K+的運進細胞,使離子分布也恢復到靜息狀態。
綜上可知,動作電位是可興奮性細胞受到刺激時在靜息電位的基礎上產生的可擴布的電位變化,表現了膜電位規律性快速去極化和復極化的變化過程。需要特別指出的是,動作電位具有“全或無”的特點:當刺激小于閾刺激時,只產生一個局部電位,不產生動作電位;當刺激大于或等于閾刺激時,使膜去極化達到或超過閾電位,便會爆發一個幅度完全相同的動作電位。通常認為,動作電位由峰電位和后電位共同組成,峰電位是動作電位的主要部分,常以峰電位來代表動作電位。因此,動作電位不能視為Na+平衡電位。
3 動作電位的傳導方式
神經纖維發生興奮的部分(內正外負)與未興奮部分(內負外正)會形成一個電位差,使電荷發生移動,形成一個局部電流。這個局部電流會使未興奮部分發生去極化,使膜電位減小到閾電位以下,進而引發未興奮部分的電壓門控Na+通道和K+通道開放,產生一個同樣的動作電位。神經纖維上的興奮通過這種方式不斷向前傳導下去。由于剛興奮過的部分的鈉離子通道會暫時還處于失活狀態不能重新打開,因此對局部電流的刺激不會再發生興奮,興奮會只向未興奮部位單向傳導下去。動作電位的峰值主要受膜內外Na+濃度差的影響,在傳導過程中,其幅度始終一樣,并不會發生衰減。
綜上分析,動作電位的傳導是由于局部電流在相鄰未興奮部分誘發產生一個新的興奮,而不是直接以局部電流的形式進行傳導。
4 結論
綜上所述,細胞內外的K+、Na+濃度差決定了Ek和ENa的大小,而膜對K+和Na+的通透性的比值決定膜電位更偏向于EK或更偏向于ENa,這是影響膜電位大小的主要原因。靜息狀態時GK遠大于GNa,膜電位就接近K+平衡電位EK,膜外K+濃度影響EK,對靜息電位影響大,而Na+濃度對靜息電位影響很小;動作電位是膜電位規律的變化過程,包括峰電位和后電位,峰電位上升時GNa遠大于GK,其峰值較接近Na+平衡電位ENa,膜外Na+濃度影響ENa,對動作電位影響大,而K+對動作電位影響很小。興奮部分與未興奮部分間的局部電流誘導后者產生新的興奮,這是動作電位傳導的方式。
