HBx基因對HepG2.2.15細胞MICA-A5.1表達及侵襲、遷移的影響
2020-04-23 09:18:02李沛,劉宇
臨床肝膽病雜志 2020年4期
關鍵詞:肝癌
李 沛, 劉 宇
南華大學附屬第一醫院 a.檢驗科; b.急診科, 湖南 衡陽 421001
肝癌是世界范圍內發病率第三位的惡性腫瘤疾病[1],發病率髙,預后差,病情進展快,易侵襲轉移,生存期短。據報道[2-3],在全球每年新增肝癌病例中,約一半發生在我國,已成為威脅我國人群健康安全的主要原因。研究[4]表明,HBV感染是引發肝癌的重要因素,HBV攜帶者罹患肝癌的風險極大提高,HBx蛋白作為HBV基因編碼的7個蛋白之一,可調節相關癌基因、抑癌基因表達,從而誘導肝癌發生[5],HBx蛋白可通過下調miR-200a-3p表達而調控HBV感染患者肝癌細胞活力和細胞周期,進而促進其增殖及侵襲轉移[6]。主要組織相容性復合體(MHC)Ⅰ類鏈相關基因A(MHC classⅠ chain-related gene A,MICA)是人類MHC基因Ⅰ類亞區的一種基因亞型[7],MICA-A5.1是其第5外顯子的等位基因,可轉錄翻譯產生可溶性MICA(soluble MICA,sMICA)分子,其與肝癌發生相關,sMICA水平高的肝硬化患者發生肝癌的風險最高[8],有研究[9]表明,sMICA可促使人骨肉瘤的侵襲及免疫逃逸,抑制其表達,可恢復抗腫瘤免疫,并抑制腫瘤進展,但HBx基因對HepG2.2.15細胞MICA-A5.1表達及侵襲、遷移的影響目前尚不清楚,本研究通過體外培養HepG2.2.15細胞系實驗對此進行探討。
1 材料與方法
1.1 試劑與材料 人肝癌細胞HepG2.2.15(寧波明舟生物科技有限公司,貨號:MZ-1350);HBx過表達質粒、HBx空載質粒、HBx siRNA、HBx siRNA陰性對照由上海吉瑪基因有限公司構建合成;MEM培養基(武漢純度生物科技有限公司,貨號:CD-100044GM);胎牛血清FBS、胰蛋白酶、青鏈霉素(美國Gibco公司,貨號分別為:10099-141、15050057、10378016);sMICA 酶聯免疫吸附法試劑盒(上海聯碩寶為生物科技有限公司,貨號:AE90082Hu);兔源GAPDH、HBx、E-cadherin、MICA及Vimentin一抗、羊抗兔二抗(美國Abcam公司,貨號分別為:ab181602、ab39716、ab76319、ab62540、ab193555、ab150077);伊紅染色液、蛋白裂解液、CKK-8試劑盒、BCA試劑盒(上海碧云天公司,貨號分別為:C0109、P0013K、C0037、P0011)。
1.2 主要儀器 Olympus CKX41顯微鏡(德國Leica公司);Centrifuge 5424R低溫高速離心機(德國 Eppendorf 股份公司);Model 680酶標儀、1659001蛋白電泳儀、Trans-Blot SD轉膜儀、CFX96 Touch Deep Well熒光定量PCR儀(美國Bio-Rad公司);凝膠成像儀(日本尼康公司)。
1.3 倫理學審查 本研究方案經由南華大學附屬第一醫院倫理委員會審批通過(批號:LL201700008)。
1.4 方法
1.4.1 細胞培養及分組處理 將購買的人肝癌細胞HepG2.2.15快速解凍復蘇,以完全培養基(含10%胎牛血清、100 U/ml青鏈霉素的MEM培養基)重懸,接種于25 cm2培養瓶中,并置于37 ℃、5%CO2、95%濕度的恒溫培養箱中培養,待細胞長至85%左右時,以胰蛋白酶消化后傳代培養。
傳代培養的細胞接種于24孔板中,隨機分為對照組、HBx過表達質粒組、HBx空載質粒組、HBx siRNA組、HBx siRNA陰性對照組,置于恒溫培養箱中培養24 h后,參照LipofectamineTM2000的說明書步驟,將轉染質粒及siRNA(質粒及siRNA濃度參照各自說明書)分別以50 μl Opti-MEM 減血清培養基溶解混勻,同時以50 μl Opti-MEM減血清培養基溶解混勻1 μl LipofectamineTM2000,均靜置20 min,將 LipofectamineTM2000溶液分別與質粒及siRNA溶液合并后輕輕混勻,然后按組別分別處理細胞,以Opti-MEM減血清培養基培養細胞6 h后,將其更換為完全培養基繼續培養。
1.4.2 CKK-8實驗 取傳代培養的HepG2.2.15細胞以0.6×105個的細胞密度接種于96孔板中,按照1.4.1中的方法分組處理,分別在12 h、24 h、48 h后加入CKK-8試劑,繼續培養2 h,于全自動酶標儀中振蕩混勻后檢測450 nm波長下各孔吸光度(optical density,OD),計算各組細胞活力,實驗重復3次。公式為:細胞活力(%)=實驗組 OD 值/對照組 OD 值 ×100%。
1.4.3 Transwell侵襲實驗 取傳代培養的HepG2.2.15細胞接種于24孔板中,按照1.4.1中的方法分組處理,于12 h后收集各組細胞,加入無血清的MEM培養基分別重懸后計數,將細胞液濃度調整為5×105個/ml,取200 μl加入基質膠包被過的Transwell上室,在其下室加入含有10%FBS的完全培養基,繼續培養24 h后,取出小室,以PBS漂洗細胞后,加入4%多聚甲醛于室溫下固定,然后以棉簽輕輕擦除基質膠及上室內細胞,加入500 μl 0.5%伊紅染液,于室溫下染色,最后用Olympus CKX41顯微鏡觀察穿膜細胞,并任選5個視野拍照,對其計數并記錄,以此作為評判細胞侵襲能力的指標。
1.4.4 細胞劃痕實驗 取傳代培養的HepG2.2.15細胞以5×105個/ml的密度接種于6孔板中,按照1.4.1中的方法分組處理,于12 h后以200 μl槍頭在六孔板中心劃一條直線(以無菌直尺作為標尺),使用PBS將劃痕中的細胞漂洗干凈,然后加入完全培養基繼續培養24 h,在Olympus CKX41顯微鏡下觀察遷移細胞數目,任選5個視野拍照,對其計數并記錄,以此作為評判細胞遷移能力的指標。
1.4.5 免疫印跡實驗 取傳代培養的HepG2.2.15細胞接種于24孔板中,按照1.4.1中的方法分組處理,于12 h后收集各組細胞,以蛋白裂解液及BCA試劑盒提取各組細胞總蛋白并測定其濃度,具體操作步驟參照說明書,制備SDS凝膠,各組取含相同質量蛋白的樣品液進行電泳,然后濕轉將分離蛋白轉移至硝酸纖維膜上,以5%的脫脂奶粉室溫封閉1 h后,分別經兔源HBx、E-cadherin、Vimentin、MICA、GAPDH一抗4 ℃孵育過夜,羊抗兔二抗室溫孵育2 h后,最后以增強型化學發光法顯色,使用凝膠成像儀觀察結果并拍照,以Image Lab軟件對條帶進行分析。
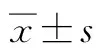
2 結果
2.1 HBx基因對細胞活力的影響 與對照組比較,HBx過表達質粒組細胞活力在24 h、48 h均升高,差異均有統計學意義(q值分別為8.268、12.680,P值均<0.001);HBx siRNA組細胞活力在24 h、48 h均降低,差異均有統計學意義(q值分別為4.365、7.523,P值分別為0.036、<0.001);為保證侵襲遷移實驗的準確性,避免HBx基因對細胞活力的影響,故選擇藥物作用時間為12 h(表1)。

表1 HBx基因對細胞活力的影響(%)
注:1)與對照組比較,P<0.05。
2.2 各組細胞HBx、MICA-A5.1蛋白表達及細胞培養基中sMICA水平的檢測 與對照組比較,HBx過表達質粒組細胞HBx及MICA蛋白表達水平、細胞培養基中sMICA水平升高,差異均有統計學意義(q值分別為9.427、6.697、5.042,P值分別為<0.001、0.001、0.006);HBx siRNA組細胞HBx及MICA蛋白表達水平、細胞培養基中sMICA水平降低,差異均有統計學意義(q值分別為8.133、8.828、7.673,P值均<0.001)(圖1,表2)。

注:A,對照組;B,HBx空載質粒組;C,HBx過表達質粒組;D,HBx siRNA陰性對照組;E,HBx siRNA組。

表2 各組細胞HBx、MICA-A5.1蛋白相對表達水平及細胞培養基中sMICA水平
注:1)與對照組比較,P<0.05。
2.3 HBx基因對細胞遷移的影響 與對照組比較,HBx過表達質粒組遷移的細胞數目升高,差異有統計學意義(q=22.740,P<0.001);HBx siRNA組遷移的細胞數目降低,差異有統計學意義(q=5.391,P=0.007)(圖2,表3)。




注:a,對照組;b,HBx空載質粒組;c,HBx過表達質粒組;d,HBx siRNA陰性對照組;e,HBxsiRNA組。
圖2HBx基因對細胞遷移的影響

表3 HBx基因對細胞遷移的影響
注:1)與對照組比較,P<0.05。
2.4 HBx基因對細胞侵襲的影響 與對照組比較,HBx過表達質粒組侵出Transwell小室的細胞數目升高,差異有統計學意義(q=15.720,P<0.001);HBx siRNA組侵出Transwell小室的細胞數目降低,差異有統計學意義(q=7.694,P<0.001)(圖3,表4)。
2.5 HBx基因對細胞上皮間質轉化(EMT)標志蛋白E-cadherin、Vimentin的影響 與對照組比較,HBx過表達質粒組細胞E-cadherin表達降低,Vimentin表達升高,差異均有統計學意義(q值分別為5.258、10.500,P值分別為0.008、<0.001);HBx siRNA組細胞E-cadherin表達升高,Vimentin表達降低,差異均有統計學意義(q值分別為6.226、7.616,P值分別為0.002、<0.001)(圖4,表5)。





注:a,對照組;b,HBx空載質粒組;c,HBx過表達質粒組;d,HBx siRNA陰性對照組;e,HBxsiRNA組。
圖3HBx基因對細胞侵襲的影響

表4 各組侵出Transwell小室的細胞數目比較
注:1)與對照組比較,P<0.05。

注:A,對照組;B,HBx空載質粒組;C,HBx過表達質粒組;D,HBx siRNA陰性對照組;E,HBx siRNA組。
圖4EMT標志蛋白E-cadherin、Vimentin的免疫印跡檢測結果

表5 各組細胞EMT標志蛋白E-cadherin、Vimentin的相對表達水平
注:1)與對照組比較,P<0.05。
3 討論
肝癌的治療以手術后放化療為主,但因早期腫瘤的轉移及術后復發,其治療效果不能令人滿意,提高對肝癌發生及侵襲轉移分子機制的理解對于肝癌的臨床治療具有重要意義。肝癌的發生機制復雜,是病毒感染、飲食習慣、遺傳等多因素長期相互作用的綜合結果[10-12],其中,HBV感染是肝癌的主要致病因素[13],合并HBV感染的肝癌患者病情進展快、生存期極短[14]。研究[15]發現,肝癌細胞Huh-7感染HBV后,HBV基因編碼的HBx蛋白在Huh-7細胞中表達,且其遷移侵襲能力增強,IFNα可通過促進其表達抗病毒蛋白而減少細胞遷移及侵襲,進而減輕HBV相關性肝癌的發生發展。因而,HBx基因可能調控HBV感染的HepG2細胞HepG2.2.15的侵襲、遷移。本研究以插入HBV全基因組并持續表達的HepG2細胞系HepG2.2.15作為研究對象,探討HBx基因對HepG2.2.15細胞侵襲、遷移的影響,以CKK-8檢測HBx基因對HepG2.2.15細胞活力的影響,結果顯示,過表達HBx基因可提升細胞活力,下調HBx基因可降低細胞活力,在HBx siRNA處理HepG2.2.15細胞24 h后其細胞活力約為59.36%,為保證侵襲遷移實驗的準確性,避免HBx siRNA對細胞活力的影響,故本研究選擇藥物作用時間為12 h進行后續實驗。研究結果表明,EMT是癌細胞發生轉移,向機體別處侵襲的關鍵病理過程[16],期間細胞極性逐漸消失,上皮細胞表型逐漸轉化為具有較高遷移與侵襲能力的間質細胞表型,同時上皮細胞標志蛋白E-cadherin表達降低,間質細胞標志蛋白Vimentin表達升高[17]。本研究以HBx過表達質粒上調HBx基因可使細胞Vimentin蛋白表達、遷移細胞數目、侵出Transwell小室的細胞數目升高,E-cadherin表達降低;以HBx siRNA下調HBx基因可使細胞Vimentin蛋白表達、遷移細胞數目、侵出Transwell小室的細胞數目降低,E-cadherin表達升高,表明HBx基因可介導HepG2.2.15細胞的侵襲轉移,抑制其表達,能夠降低HBV感染的HepG2.2.15細胞侵襲轉移能力,進一步證實了HBV基因能夠促進肝癌細胞遷移侵襲及癌癥進展。
MICA基因含有6個外顯子,MICA-A5.1作為其第5外顯子等位基因,可使蛋白質翻譯提前結束,從而相比其他外顯子更易形成sMICA分子,當其被腫瘤、病毒或細菌等刺激時,可表達上調,產生更多的sMICA分子。研究[18-19]發現,MICA基因型及sMICA水平與慢性丙型肝炎相關肝細胞癌的發生顯著相關,降低sMICA水平對治療索拉非尼耐藥的肝細胞癌有效。此外,sMICA水平在胰腺癌組織中的表達水平異常且與腫瘤的侵襲轉移有關,可作為判斷胰腺癌預后的重要生物標志物[20],因此,MICA-A5.1基因是調控HepG2.2.15細胞侵襲轉移的潛在作用靶點,但HBV基因是否可調控其表達,進而影響HepG2.2.15細胞侵襲的轉移,目前還未見研究。本研究對此進行了探討,結果顯示,上調HBx基因可升高HepG2.2.15細胞HBx、MICA蛋白表達水平及細胞培養基中sMICA水平,促使HBV感染的HepG2.2.15細胞侵襲轉移;下調HBx基因可降低細胞HBx、MICA蛋白表達水平及細胞培養基中sMICA水平,抑制HepG2.2.15細胞侵襲轉移,表明HBx基因可上調HepG2.2.15細胞中MICA-A5.1基因表達,進而促進其侵襲轉移,揭示了HBx基因促進HBV感染的HepG2.2.15細胞的侵襲遷移的作用機制可能是下調MICA-A5.1基因。
總之,HBx基因可調控HBV感染的HepG2.2.15細胞中MICA-A5.1基因表達,介導細胞侵襲轉移,上調該基因可促進HepG2.2.15細胞表達MICA-A5.1基因,進而增強其侵襲轉移能力,表明調控MICA-A5.1基因表達可能是HBx基因介導HBV感染的HepG2.2.15細胞侵襲轉移的分子機制之一。但本研究只是進行了初步研究,未通過上調及下調MICA-A5.1基因進行對照驗證,也未涉及HBx調控MICA基因的具體分子機制,需要后續進一步深入研究。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
