撫育間伐對栓皮櫟種群空間分布格局的影響
2020-05-29 09:06:14閆東鋒賀文馬瑞婷楊喜田
生態環境學報 2020年3期
關鍵詞:生長
閆東鋒,賀文,馬瑞婷,楊喜田
河南農業大學林學院,河南·鄭州 450002
林木種群空間分布格局是林木種群在水平空間上的排列方式,可以反映種群對外來干擾和環境要素的適應策略(吳盼等,2019;Gang et al.,2007),對于理解植物群落構建和適應機制具有重要意義。空間分布點格局和空間關聯性是研究種群空間分布格局的兩個主要內容(Muhamed,2019)。點格局分析能夠量化特定林分內林木個體在不同空間尺度上的空間分布和種間共存格局,在林木空間分布格局研究中廣泛應用(汪洋等,2016;涂洪潤等,2019)。空間關聯性是指不同類型的林木個體在空間上的相互關系(蘭航宇等,2019),往往是由生境差異或者外來干擾造成的,研究空間關聯性有助于理解種群與環境及不同生長階段林木個體之間的關系(Zhou et al.,2019),是揭示林分動態的重要手段。
撫育間伐是一種重要的改善林木生長條件的經營措施,它通過減小林分密度,調整林木個體競爭關系,改善影響林木生長的光照條件和土壤環境,進而促進保留木生長和天然更新(Cabon et al.,2018;Yang et al.,2017;閆東鋒等,2019)。目前,生產中的撫育間伐主要是以修枝、清灌、清理生長不良和低質低效林木個體為主的低間伐強度的作業方式。在研究撫育間伐對林分發育的影響時,諸如蓄積量、生長量等林木生長指標短時間內難以被準確評價(胡雪凡等,2019),而林分空間結構作為森林經營可以調控的因子,能夠較快的地反映出經營效果,可為制定科學的經營措施提供理論基礎。近年來,有關撫育間伐對林分的影響研究主要集中在林分生長(朱子卉等,2018)、物種多樣性(魏安然等,2019)、土壤性質(徐雪蕾等,2019;Cheng et al.,2012)、土壤微生物(Kim et al.,2019)、天然更新(閆東鋒等,2019)等方面,對林分空間結構的影響研究相對較少,且相關研究主要集中在使用空間結構參數(如混交度和角尺度)及傳統的空間格局診斷方法(如Moristita指數)(Kint et al.,2003;曹小玉等,2015),而基于大樣地定位數據研究撫育間伐對栓皮櫟種群空間分布格局的研究未見報道。
栓皮櫟(Quercus variabilis)人工林是河南山區典型的落葉闊葉林,科學的撫育方式將對栓皮櫟林生態防護功能持續發揮和大徑級木材生產起著舉足輕重的作用。本文以 42年生的撫育間伐(蓄積強度3.23%)栓皮櫟人工林為研究對象,以未間伐林分為對照,通過設置2個100 m×100 m的大樣地,使用點格局分析和空間關聯性分析方法研究了撫育間伐對栓皮櫟種群空間分布格局的影響,旨在探究在生產性撫育間伐背景下,栓皮櫟種群空間結構與格局變化的內在機制,為栓皮櫟人工林的定向培育提供方法和理論支持。
1 研究區概況
研究區位于河南省國有登封林場清涼寺林區,屬暖溫帶大陸性季風氣候,冬季寒冷干燥,夏季高溫多雨,季風性顯著;全年平均溫度14.2 ℃,無霜期238 d左右;平均降水量614 mm,降水量主要分布在夏季(6—8月),占全年降水總量的 33.8%左右。境內土壤以褐土和棕壤為主,主要樹種有栓皮櫟、麻櫟(Quercus acutissima)、槲櫟(Quercus aliena)、側柏(Platycladus orientalis)、油松(Pinus tabuliformis)、刺槐(Robinia pseudoacacia)、黃櫨(Cotinus coggygria)、化香(Platycarya strobilacea)、椿樹(Ailanthus altissima)、榆樹(Ulmus pumila)等。
2 研究方法
2.1 樣地設置與調查
結合登封林場日常經營活動,2016年在栓皮櫟人工林內開展以間伐為主的生產性撫育間伐作業,主要是通過割灌、清理劣質木、伐除沒有培養前途的林木個體來改善林分結構。栓皮櫟間伐及未間伐林分年齡均為 40年生,郁閉度約 0.90,林分密度分別為 2200 plant·hm-2和 2211 plant·hm-2,平均胸徑分別為10.01 cm和10.10 cm,平均樹高分別為7.98 m和8.06 m。在每木檢尺的基礎上,根據一元材積表計算林分蓄積量,間伐前后樣地蓄積量分別為 96.864 m3·hm-2和 93.697 m3·hm-2,間伐株數強度為31.55%,蓄積強度為3.23%;未間伐樣地林分蓄積量為115.615 m3·hm-2。間伐前后及未間伐林分基本情況見表1。
在間伐林分內選取有代表性的地塊,同時選擇與間伐作業前林分的林分結構、生長階段、立地條件以及經營歷史一致的未間伐地塊內各設置 1塊100 m×100 m的樣地,研究區及樣地位置示意圖見圖1。
生產性撫育間伐作業2年后,利用全站儀(南方測繪,NTS-332R)對100 m×100 m樣地內胸徑≥2.0 cm的喬木個體進行每木定位,分別采用圍尺和Haglof激光測高儀測定胸徑和樹高,同時記錄樹種名稱、胸徑和樹高等測樹因子,并采用一元材積表法計算林分蓄積量。經緩沖區分析后,間伐和未間伐樣地林分密度分別為1448 plant·hm-2和 2078 plant·hm-2,蓄積量分別為 96.282 m3·hm-2和 118.150 m3·hm-2。樣地具體情況見表 2。
2.2 數據處理
2.2.1 數據整理
分別撫育樣地和對照樣地,采用徑級結構代替年齡結構方法,根據研究區栓皮櫟種群生長現狀及調查樣地栓皮櫟種群林木直徑分布狀態(劉憲釗等,2017a;周建云等,2012;舒蘭等,2019),將栓皮櫟種群按照胸徑大小劃分為幼樹(2 cm≤DBH<5 cm)、小樹(5 cm≤DBH<13 cm)和大樹(DBH≥13 cm)3個生長階段,并整理出栓皮櫟種群3個生長階段林木個體坐標信息,用于點格局及空間關聯性分析。
2.2.2 點格局分析
點格局法是由Ripley首先提出的一種空間分布格局分析方法(Ripley,1977),它利用Ripley’sK函數及其衍生的g(r)函數對點數據集進行不同距離尺度的聚類程度研究。

表1 間伐與未間伐林分基本情況Table 1 Basic situation of thinned stand and unthinned stand

圖1 研究區及樣地位置示意圖Fig.1 Location map of study area and the plots

表2 樣地基本情況Table 2 Basic situation of the plots
Ripley’sK函數K(r)計算公式如下:

式中,A表示樣地面積;n表示種群個體總數;Wij表示以點i為圓心;uij為半徑的圓周長落在A內的比例,代表1個點被觀察到的概率,為消除邊緣效應的權重因子;uij表示兩隨機點i、j之間的距離;r表示空間尺度;Ir(uij)為指示函數。當uij≤r時,Ir(uij)=1.0;當uij>r時,Ir(uij)=0。
g(r)是K(r)函數的概率密度函數,以環代替K函數中的圓,它能夠較好地排除K(r)函數在小尺度上引起的累積效應(張俊艷等,2014;王敬等,2014),使空間分布格局的研究更加直觀準確,具有更廣的適用性(Zhao et al.,2012)。g(r)函數計算公式如下:

式中,dK(r)是函數K(r)的微分;dr是半徑r的微分。當g(r)=1.0時,個體分布格局呈完全空間隨機分布;當g(r)<1.0時,個體分布為均勻分布;當g(r)>1.0時,個體分布為聚集分布。
利用Programita軟件(2014版) (Wiegand et al.,2014),以樣地最短邊長的一半為最大空間距離尺度(本研究為50 m),步長值為1 m,利用完全隨機零假設模型,通過199次Monte-Carlo隨機模擬,得出由上下2條包跡線所構成的95%的置信區間和理論值。當g(r)觀測值位于置信區間之內,種群空間分布為隨機分布;當g(r)位于置信區間以上,為聚集分布;當g(r)位于置信區間以下,為均勻分布。
2.2.3 空間關聯性分析
空間關聯性分析可以描述種間或種內不同發育階段林木個體之間的關系(潘磊磊等,2019;舒蘭等,2019),成對相關函數K12(r)計算公式如下:

式中,A,Wij,uij,r,Ir(uij)含義同公式(1);n1和n2分別為變量1和變量2的個體總數(本研究為 2個不同生長階段的種群個體總數)。用g12(r)函數對K12(r)函數進行改進,成對相關g12(r)函數計算公式如下:

利用Programita軟件(2014版),基于先決條件零假設模型(Wiegand et al.,2004),空間距離尺度設為0—50 m,步長值1 m,Monte Carlo模擬199次,生成置信區間為95%的上下包跡線和理論值。當g12(r)位于下包跡線以下和上包跡線以上,分別表示這兩個變量在空間尺度r處呈顯著負關聯或顯著正關聯;若g12(r)位于上下包跡線內,則兩個變量之間無顯著關聯性。
3 結果與分析
3.1 撫育間伐對栓皮櫟種群空間分布點格局的影響
采用式(2)計算間伐樣地和未間伐樣地不同空間距離尺度條件下栓皮櫟種群g(r)函數觀測值、理論值及上下包跡線,結果如圖2所示。由圖可知,間伐樣地和未間伐樣地栓皮櫟種群空間點格局均與空間距離尺度有顯著的相關關系,其中,間伐樣地栓皮櫟種群在0—2 m距離尺度內呈現均勻分布,在4—16 m距離尺度內表現為聚集分布,在其他距離尺度內則呈現隨機分布;而未間伐樣地栓皮櫟種群在0—50 m尺度上基本上呈現聚集分布(除33—37 m表現為隨機分布外),但隨著距離尺度增加,其聚集強度有減弱的趨勢。間伐樣地和未間伐樣地栓皮櫟種群分別在距離尺度由0 m增加到4.5 m和1.5m 時,g(r)函數值達到最大,分別為 1.039和1.106;分別在16—50 m和34—50 m的空間距離尺度內,g(r)函數觀測值波動不大,整體上表現為隨機分布。在各個距離尺度上,間伐樣地的g(r)函數觀測值均小于未間伐樣地,這說明間伐調整了林分栓皮櫟種群的空間排列形式,使得在一定空間距離尺度內,林木個體間的競爭趨于緩和。
3.2 撫育間伐對不同生長階段栓皮櫟種群空間分布格局的影響
3.2.1 撫育間伐對栓皮櫟種群株數分布的影響
統計樣地內栓皮櫟種群不同生長階段株數及其比例(表 3)可知,間伐和未間伐樣地栓皮櫟幼樹、小樹和大樹株數比例關系分別為 8∶741∶699和261∶1134∶683。間伐樣地幼樹和小樹所占比例均小于對照樣地,而大樹所占比例高于對照樣地。這說明,生產性撫育間伐作為一種外來干擾,其作業活動本身可能直接造成幼樹及小樹數量的大幅度減少,進而顯著改變了不同生長階段栓皮櫟種群株數分布,從長期來看,這可能并不利于次林層的培育。
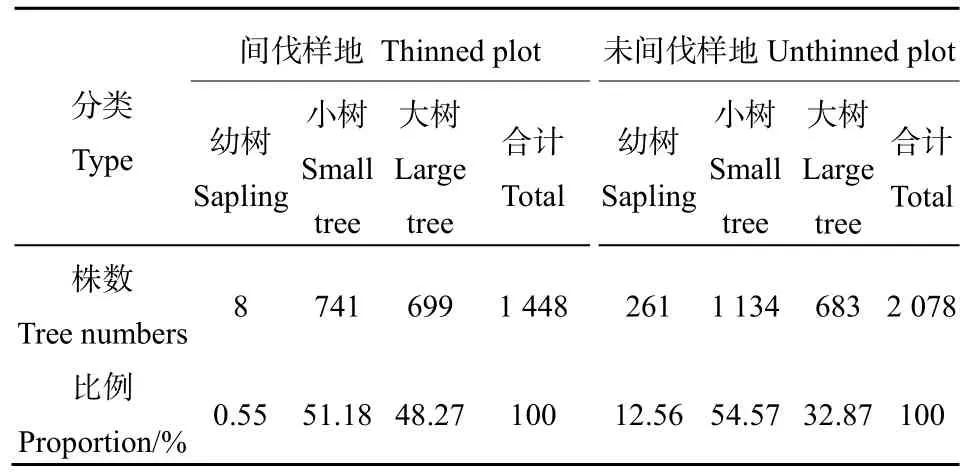
表3 間伐和未間伐樣地不同生長階段栓皮櫟種群株數分布Table 3 Tree numbers distribution of Quercus variabilis population at different growth stages in thinned and unthinned plots
3.2.2 撫育間伐對不同生長階段栓皮櫟種群空間分布點格局的影響
對間伐樣地和未間伐樣地,分別計算栓皮櫟幼樹、小樹和大樹不同距離尺度條件下的g(r)函數觀測值、理論值和上下包跡線,結果如圖3所示。由圖可知,栓皮櫟幼樹、小樹和大樹林木個體空間分布點格局隨距離尺度的增加而呈現不同的分布特征。在間伐樣地,栓皮櫟幼樹空間分布點格局在0—14 m和 17—24 m的距離尺度上表現為聚集分布,而在14—17 m和24—50 m的距離尺度上表現為隨機分布;小樹個體空間分布點格局在 0—1 m的小距離尺度內表現為隨機分布,在 1—16 m和16—50 m 的距離尺度內分別呈現出聚集分布和隨機分布特征;栓皮櫟大樹在不同距離尺度上呈現出多樣化的分布狀態,如在0—3 m距離尺度內為均勻分布,在3—6 m距離尺度內為隨機分布,6—9 m距離尺度內為弱的聚集分布,而在9—50 m的距離尺度上表現為隨機分布。

圖2 間伐和未間伐栓皮櫟種群空間分布點格局Fig.2 Spatial point distribution patterns of thinned and unthinned Quercusv ariabilis population

圖3 不同生長階段間伐和未間伐栓皮櫟種群空間分布點格局Fig.3 Spatial point distribution patterns of thinned and unthinned Quercusv ariabilis population at different growth stages
與間伐樣地相比,未間伐樣地不同生長階段栓皮櫟種群在不同空間距離尺度上呈現出不同的分布狀態,其規律性并不明顯。如栓皮櫟幼樹個體空間分布點格局分別在0—9、13—14、16—19、23—28 m空間距離尺度上表現為聚集分布,而分別在 9—13、14—16、19—23、28—50 m 距離尺度上表現為隨機分布;栓皮櫟大樹個體空間分布點格局在0—3 m距離尺度內呈均勻分布,分別在3—5、13—16、25—27、29—34、37—39 m 及 41—50 m距離尺度上呈隨機分布,分別在5—13、16—25、27—29、34—37、39—41 m 距離尺度內表現為弱的聚集分布。但是,栓皮櫟小樹個體空間分布點格局在0—21 m的很大空間距離尺度內表現為聚集分布,g(r)函數觀測值雖在21—50 m的距離尺度上有一定地波動,但均處于上下包跡線內,其點格局總體表現為隨機分布。
3.3 撫育間伐對不同生長階段栓皮櫟種群空間關聯性的影響

圖4 不同生長階段間伐和未間伐栓皮櫟種群空間關聯性Fig.4 Spatial association with different growth stages of thinned and unthinned Quercus variabilis population
為了研究撫育間伐對不同生長階段栓皮櫟種群空間關聯性的影響,采用雙變量g12(r)函數(式4),運用先決條件零假設模型,在0—50 m空間距離尺度內,分析間伐樣地和未間伐樣地內不同生長階段栓皮櫟種群之間的g12(r)函數觀測值變化規律,結果見圖4。由圖4可知,間伐樣地不同生長階段栓皮櫟種群之間在較小的空間距離尺度內呈現一定的相關性,但在較大的空間距離尺度上并沒有明顯的相關性,如栓皮櫟幼樹和小樹之間在0—6 m距離尺度上呈顯著正相關,而在6—50 m的距離尺度上無顯著關聯性;栓皮櫟幼樹和大樹之間,以及小樹和大樹之間,分別在0—5 m和0—2 m較小的距離尺度上呈顯著正關聯,而分別在5—50 m和2—50 m的距離尺度上無顯著關聯性。與間伐樣地相比,對照樣地不同生長階段栓皮櫟種群之間的空間關聯性呈現出不規則的變化規律,如幼樹和小樹之間分別在0—2、4—6、36—39 m及41—50 m的距離尺度上無顯著關聯性,而分別在2—4、6—36、39—41 m距離尺度上呈顯著正相關;幼樹和大樹之間分別在0—6、38—43、44—50 m的距離尺度上無顯著關聯性,而在6—38 m和43—44 m距離尺度上呈顯著正相關。
4 討論
點格局分析的基本原理是將林分內每株林木個體視作二維空間內的一個點,以全部林木個體在一定空間內所構成的二維空間點圖為基礎對空間分布格局進行分析(胡滿等,2019;張曉晨等,2018)。林木空間分布點格局受環境異質性、外部干擾(如人類經營活動)等各種因素影響,與林分自然稀疏過程以及生長環境變化密切相關(王曉榮等,2019),且在林分不同生長發育階段會呈現出不同的分布格局。林木空間分布點格局可分為聚集分布、隨機分布及均勻分布3種類型(Kanaji et al.,2017)。一般認為,小尺度上的種群分布格局大多是因為種子的擴散限制以及物種自身生物學特性所作用的結果,大尺度上的點格局則主要受環境異質性的影響(劉憲釗等,2017b)。
相關研究表明,撫育間伐通過降低林分密度,減緩林木競爭,進而促進保留木生長,這個過程必然對保留木空間分布格局產生影響(王蒙等,2016;段夢成等,2019)。本研究發現,在0—50 m距離尺度內,間伐樣地和未間伐樣地均呈現出由聚集分布向隨機分布過渡,這與有關遼東櫟和其他種群研究結果類似(伊力塔等,2008),這種現象被認為是與櫟類本身的生物學特性有關。相比于間伐樣地,未間伐樣地在更大尺度(16—33 m)內的呈現聚集分布,這可能與未間伐樣地過高的林分密度,尤其是幼樹密度(261 plant·hm-2)有關,小徑級的林木個體在小尺度上的聚集分布,被認為在一定程度上可以提高抵御外來干擾的能力。同時可以看出,空間距離尺度的選擇往往會影響點格局分布特征的判斷,大部分的研究由于樣地面積過小,只能選擇0—10 m或0—20 m的空間距離尺度,本研究基于大樣地數據,選擇了0—50 m的距離尺度,能夠在比較大的距離尺度內研究種群空間分布點格局,這有利于指導在大的林分尺度上制定更精細化的森林經營措施。
本研究發現,未間伐樣地栓皮櫟小樹個體在0—21 m距離尺度內表現為聚集分布,而間伐后在1—16 m距離尺度內表現為聚集分布,而在其他距離尺度上表現為隨機分布,這是因為櫟類依靠母樹的天然更新,會創造以結實量比較大的母樹為中心的生長環境,但是生產性的撫育間伐,可能會砍伐大量的劣質幼樹和小樹,使得在較大的距離尺度內表現為隨機分布,這種趨勢可以改善保留的優質幼樹小樹的生態位,促進對水分、養分等林地資源的充分利用(Podlaski,2019),有利于栓皮櫟種群的生存與發展。另外,撫育間伐方式的不同,對保留木種群空間分布格局的影響并不相同,如以目標樹作業法為作業方式的近自然林經營,因為在撫育過程中,注重保護幼苗幼樹和小樹,會造成間伐后幼苗幼樹的聚集分布更為明顯(周建云等,2012)。而在生產實際中,生產性撫育間伐較少考慮木材收獲,其主要作業方法仍然是以修枝、清灌、清理生長不良和低質低效林木為主的傳統作業方式,由于栓皮櫟幼樹和小樹很多是萌生起源的,其干形和生長狀態往往不佳,容易在作業時被作為劣質木砍伐掉。本研究發現,栓皮櫟幼樹和小樹密度在間伐樣地內僅為8 plant·hm-2和741 plant·hm-2,而在未間伐樣地為 261 plant·hm-2和1134 plant·hm-2。因此,雖然從長期來看,間伐會促進林下天然更新和次林層的形成,但在生產性撫育間伐作業后的短時間內,由于作業活動本身的影響,不同生長階段的林木個體株數比例差異較大,這必然導致不同生長階段的林木個體空間分布格局的差異,進而對其生長動態產生影響。
5 結論
本研究發現,撫育間伐后栓皮櫟幼樹和小樹之間、幼樹和大樹之間均在小尺度(0—6 m)上呈正關聯,在其余距離尺度無關聯,說明栓皮櫟種群幼樹和小樹、幼樹和大樹之間在小尺度上因對環境資源需求相近而聚集共存,在其余很大距離尺度內相對獨立,并無資源競爭關系。經撫育間伐后,栓皮櫟種群內不同生長階段林木個體的關聯性得到了弱化,這有利于保留木的快速生長。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
