外源水楊酸對鹽脅迫下小白菜幼苗生理的影響
2020-06-09 02:35:08黃程文小梅唐殷郭磊周江世杰代其林
江蘇農業科學 2020年7期
黃程 文小梅 唐殷 郭磊周 江世杰 代其林
摘要:采用葉面噴施法探討水楊酸(SA)對不同時間段NaCl脅迫下小白菜(Brassica campestris L.)幼苗生理的影響。結果表明,隨著鹽脅迫時間的延長,SA預處理的小白菜幼苗葉片葉綠素含量、相對含水量、脯氨酸含量、可溶性糖含量、可溶性蛋白質含量、過氧化物酶(POD)活性和超氧化物歧化酶(SOD)活性以及相關基因的表達量均高于對照處理;而丙二醛含量和相對電導率均低于對照處理。在整個鹽脅迫過程中,相對含水量、葉綠素含量、可溶性蛋白含量、可溶性糖含量以及脯氨酸含量呈不斷下降的趨勢;丙二醛含量和相對電導率呈不斷上升的趨勢;POD活性以及SOD活性呈先上升后下降的趨勢,峰值均出現在3 d。POD基因與SOD基因的相對表達量呈先上升后下降的趨勢,且峰值出現在3 d。分析結果表明,外源水楊酸可以通過提高葉綠素含量、相對含水量、滲透調節物質含量來上調抗氧化酶基因的表達,增強抗氧化酶活性,并且減輕細胞膜損傷來緩解鹽脅迫對小白菜幼苗的傷害,從而提高小白菜幼苗的抗鹽能力。
關鍵詞:水楊酸;小白菜;鹽脅迫;生理生化;基因表達
中圖分類號: S634.301文獻標志碼: A
文章編號:1002-1302(2020)07-0147-05
據統計,地球鹽漬土地面積約占陸地總面積的1/3,而土壤鹽漬化問題給農業發展帶來了諸多負面影響,給農作物的生產帶來了極大的阻礙。目前,也出現了許多土壤改良劑、土壤調理劑來改善土壤中出現的各種問題,但并不能從根本上解決土壤鹽漬化對植物造成的傷害[1-2]。土壤鹽分降低了土壤資源價值,給生產性農業帶來巨大損失,對環境造成威脅,從而加大了對經濟和環境的危害[3]。當植物體處于鹽脅迫環境下時,會導致體內活性氧的大量產生并且導致活性氧清除系統之間的失衡。活性氧的累積會引起膜質過氧化反應并且傷害細胞膜[4],而植物體內產生的活性氧可以通過抗氧化酶之間的協同作用來清除[5-6]。桑利敏研究發現,當甜菜幼苗處于鹽脅迫條件下時,其出芽率、葉綠素含量、超氧化物歧化酶(SOD)、過氧化物酶、過氧化氫酶和抗壞血酸過氧化物酶(APX)的活性等各項生理指標均呈現下降趨勢,植物生長受到限制[7]。師晨娟等論述了多胺、細胞分裂素、乙烯、脫落酸等激素對鹽脅迫下植物的緩解作用與作用機制[8],如吲哚乙酸對鹽脅迫下的番茄幼苗具有緩解作用,脫落酸(ABA)可以提高鹽脅迫下小麥的產量等[9-10]。水楊酸(SA)可以提高煙草對病毒的抗性,并且在黃瓜體內可以作為介導系統獲得性抗性(SAR)傳遞的內源信號[11-12],可以提高植物體的抗逆性[13-14]。
目前,關于SA對小白菜幼苗抗鹽生理方面的研究較少,探究SA對鹽脅迫下小白菜幼苗生理的影響,有利于為日后小白菜在鹽脅迫環境的栽培生產提供一定的理論參考。本研究以小白菜為試驗材料,研究鹽脅迫下SA對小白菜幼苗生理生化過程的影響。
1 材料與方法
1.1 材料
供試小白菜品種為溫州快菜,購自綿陽龍門農貿市場。溫州快菜生長周期短,生育期60 d,我國南北各地均可種植。
1.2 方法
選取飽滿的小白菜種子,用1% NaClO溶液消毒處理10 min,用蒸餾水清洗后在25 ℃的溫室中浸種2 d,然后置于25 ℃組織培養室內催芽2 d。選取長勢一致的小白菜出芽種子,均勻地栽種在盛有石英砂的塑料盤中(規格為120 cm×30 cm),共12盤,然后加入清水,放入(25±1) ℃、光/暗周期12 h/12 h、光流密度200 μmol/(m2·s)條件下的培養室中培養。當小白菜幼苗第1張葉張開時,開始添加MS培養液,培養至3葉1心期。把小白菜幼苗分成2組,一組小白菜幼苗葉片上均勻噴施濃度為0.5 mmol/L的水楊酸溶液,另一組小白菜葉片噴施蒸餾水作為對照。靜置1 d后,去掉盤中原來的MS培養液,再將噴施了SA和蒸餾水的小白菜幼苗置于含有200 mmol/L 的NaCl溶液中培養1 d,從上往下數取小白菜幼苗的第3張幼葉為試驗材料,每個試驗均重復3次,取平均值。
1.3 指標測定方法
按照丙酮乙醇混合法[15]測定葉綠素含量;按照浸泡法[16]測定相對電導率;按照王學奎的方法[17]測定可溶性糖含量;按照茚三酮法[18]測定脯氨酸含量;按照陳建勛等的方法[19]測定相對含水量;用NBT光還原法測定SOD活性;用Omran法[20]測定POD活性;按照Lowry法[20]測定可溶性蛋白含量;按照硫代巴比妥酸(TBA)比色法[18]測定丙二醛含量。
2 結果與分析
2.1 SA對鹽脅迫下小白菜幼苗葉片生理干旱情況的影響
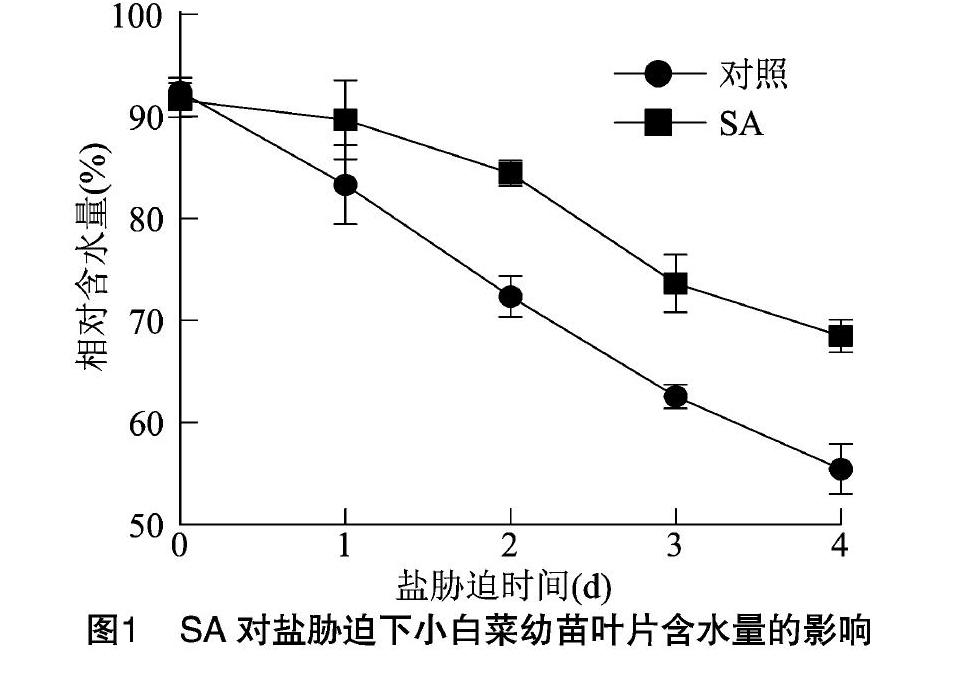
植物葉片的相對含水量可以反映植物對鹽脅迫所帶來的生理干旱的抵抗能力。在一定范圍內,植物葉片含水量高,說明其體內的生理生化反應可以正常進行,能夠保證正常的新陳代謝[21-22]。由圖1可知,隨著鹽脅迫時間的延長,小白菜幼苗葉片含水量明顯下降,在鹽脅迫過程中,經SA預處理的小白菜幼苗葉片含水量均高于對照處理(蒸餾水處理),在鹽脅迫的4 d時,SA處理的小白菜和對照處理的小白菜的含水量差達到最大。表明噴施SA有利于減緩小白菜幼苗在鹽脅迫下含水量的下降,緩解了小白菜幼苗的生理干旱,增強了小白菜的鹽脅迫抗性。
2.2 SA對鹽脅迫下小白菜幼苗質膜的影響
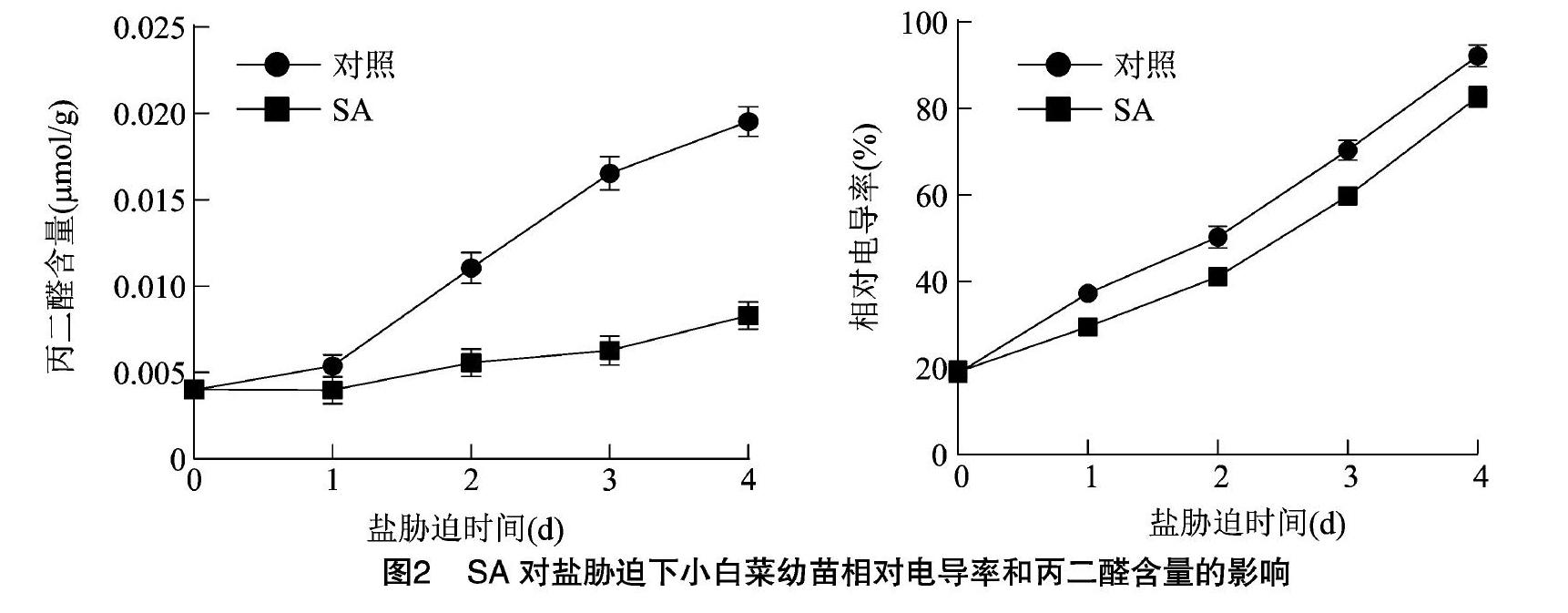
當植物處于非生物脅迫下時,其質膜會受到傷害,相對電導率和丙二醛含量增加[21-22]。由圖2可知,相對電導率和丙二醛含量均隨著鹽脅迫時間的延長而增加,經SA預處理的小白菜幼苗相對電導率和丙二醛含量均低于對照處理。鹽脅迫4 d時,SA處理和對照處理小白菜的相對電導率差值達到最大;在鹽脅迫的4 d時,SA處理和對照處理的小白菜丙二醛含量差值達到最大。總體而言,在鹽脅迫的過程中,SA處理的小白菜相對電導率和丙二醛含量均低于對照處理,表明噴施SA有利于緩解小白菜在鹽脅迫下的膜質過氧化反應,增強了小白菜的鹽脅迫抗性。
2.3 SA對鹽脅迫下小白菜幼苗葉片光合作用的影響
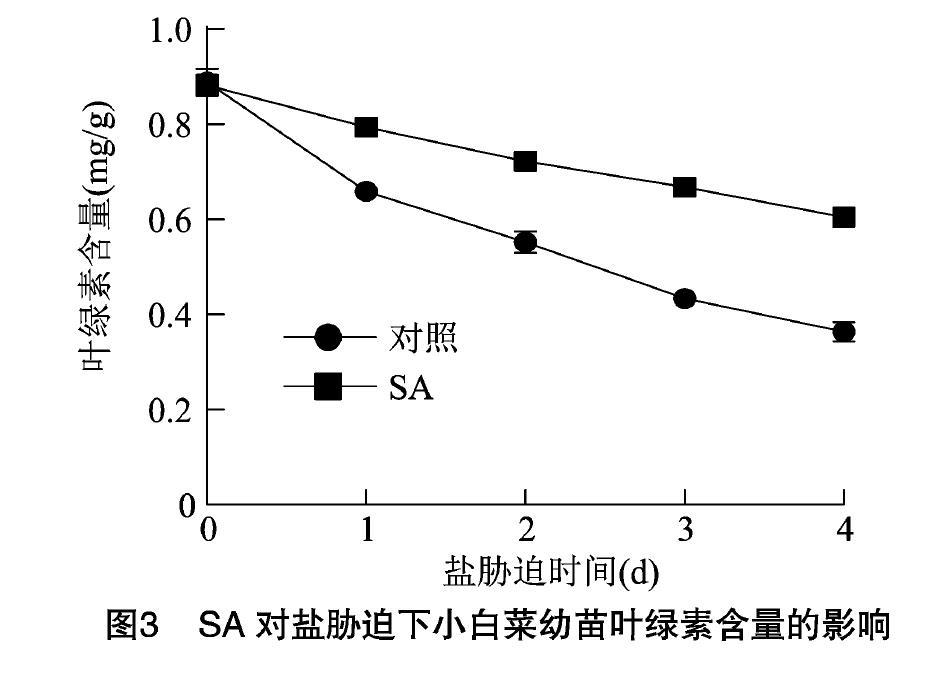
葉綠素是光合作用重要的色素分子,決定著光合作用的效率。因此,葉綠素含量是體現植物抵抗非生物脅迫能力的重要指標[21-22]。由圖3可知,隨著鹽脅迫時間的延長,小白菜幼苗葉片葉綠素含量明顯下降,而SA預處理的小白菜葉幼苗綠素含量下降速度比對照更慢。在鹽脅迫4 d時,SA處理小白菜和對照處理小白菜葉綠素含量差值最大。表明噴施SA有利于減緩鹽脅迫下小白菜葉綠素的降解,提高光合作用效率,有助于增強小白菜的鹽脅迫抗性。
2.4 SA對鹽脅迫下小白菜幼苗葉片滲透勢的影響
可溶性糖、可溶性蛋白和脯氨酸是植物體內重要的滲透調節物質,可以降低細胞滲透壓,減少失水,降低植株受到的脅迫傷害[20-21]。由圖4可知,隨著鹽脅迫時間的延長,小白菜幼苗葉片可溶性糖、可溶性蛋白和脯氨酸3種滲透調節物質的含量均呈上升趨勢。在鹽脅迫的同一時間內,SA處理小白菜幼苗葉片的可溶性糖、可溶性蛋白和脯氨酸的含量均高于對照處理。鹽脅迫剛開始時,SA和對照處理的小白菜幼苗滲透調節物質含量差異不明顯,隨著脅迫時間的延長,SA處理的小白菜幼苗葉片滲透調節物質明顯高于對照處理。脅迫處理3 d時,SA處理和對照處理可溶性糖含量差達到最大。脯氨酸含量和可溶性蛋白含量均在脅迫的4 d時達到最大差值。結果表明,噴施SA可以提高鹽脅迫下小白菜幼苗的滲透物質含量,有利于降低細胞的水勢,減少小白菜失水,降低小白菜受到的鹽脅迫傷害,增強了小白菜抵抗鹽脅迫的能力。
2.5 SA對鹽脅迫下小白菜幼苗抗氧化酶的影響
植物受到非生物脅迫時,體內會產生大量氧自由基,對植物體造成傷害。在長期進化過程中,植物體形成了具有清除活性氧自由基能力的抗氧化防御體系,其中SOD和POD是重要的保護酶類[20]。由圖5可知,隨著鹽脅迫時間的延長,SOD和POD活性均呈先上升后下降的趨勢,且均在鹽脅迫3 d后開始下降。SA處理和對照處理小白菜SOD、POD活性的最大差值均出現在脅迫3 d時。整個脅迫過程中,SA處理的小白菜抗氧化酶活性均高于對照處理,表明噴施SA有利于保護鹽脅迫下小白菜的抗氧化酶系統,增強了小白菜抵抗鹽脅迫的能力。
2.6 SA對鹽脅迫下小白菜幼苗抗氧化酶基因表達的影響
為探究小白菜幼苗的耐鹽機制,用實時熒光定量PCR研究POD和SOD基因在NaCl脅迫下的表達。由圖6可知,NaCl脅迫1 d以后,小白菜幼苗POD和SOD基因表達水平與對照相比均存在明顯差異。其中,未進行NaCl處理時,POD和SOD基因有少量表達;在NaCl處理后,小白菜幼苗POD和SOD基因的相對表達水平均有不同程度的提高,POD和SOD基因的表達量均在NaCl脅迫3 d時達到最高水平,與它們的酶活性變化具有一致的趨勢。表明外源水楊酸通過上調鹽脅迫下小白菜幼苗抗氧化酶基因的相對表達量來增強其抗氧化酶活性,進而增強了小白菜抵抗鹽脅迫的能力。
3 結論與討論
在植物體生長的不同時期,其鹽脅迫抗性存在一定的差異,植物在苗期對鹽脅迫更為敏感[23-24],因此選取了苗期的小白菜進行試驗。
當處于鹽脅迫下時,植物體內的生理特性發生巨大的變化[25]。由于植物自身的抗逆機制,植物體內會迅速積累可溶性蛋白、可溶性糖、脯氨酸等滲透調節物質。當脅迫達到一定程度時,植物自身抗逆機制會受到不可逆轉的傷害,進而導致植物體滲透調節物質含量的下降[26]。同時鹽脅迫也會引起植物體內氧自由基的大量產生,其中POD和SOD等抗氧化酶組成的抗氧化酶系統可以清除植物體內的氧自由基[20]。本研究中沒有鹽脅迫時,SA處理和對照處理的各項生理影響并沒有明顯差別,表明在正常的生長環境下,噴施SA對小白菜體內的生理生化反應并無明顯影響,這與富春元等的研究結果[27]一致。鹽脅迫對小白菜造成的傷害會通過某種信號傳遞的方式來影響小白菜的生理生化指標,而生理生化指標的變化會體現在這種信號傳遞完成之后。隨著鹽脅迫的進行,對照處理的小白菜POD和SOD指標先升高后降低,與杜世章等的研究結果[28]一致。表明在沒有噴施SA的情況下,小白菜幼苗在面對鹽脅迫時,自身也會有一種抗逆機制,進而使抗氧化酶活性在一定范圍內上升。當脅迫時間繼續延長之后,這種抗逆機制會受到破壞,進而使抗氧化酶活性下降,甚至可能會低于正常生長的小白菜抗氧化酶活性,直至小白菜死亡。當鹽脅迫時間足夠久時,將會對小白菜本身的抗逆機制造成不可逆轉的傷害,但具體機制有待進一步的考證與研究。
鹽脅迫下小白菜幼苗相關抗氧化酶基因的相對表達量發生了變化。在沒有使用NaCl處理之前,SOD基因與POD基因僅有微量表達。在使用NaCl處理之后,2種抗氧化酶基因的表達量均有不同程度的上調,且均在脅迫3 d時達到峰值,之后表達量均有下調。與代其林等關于豇豆在鹽脅迫下抗氧化酶活性以及相關基因表達的研究結果[20]吻合。顯然,這2種抗氧化酶基因都是誘導表達型基因,均存在過量表達的可能性。
參考文獻:
[1]史慶華,朱祝軍,Khalidal-Aghabary K,等. 等滲鹽脅迫對番茄抗氧化酶和ATP酶及焦磷酸酶活性的影響[J]. 植物生理與分子生物學學報,2004,30(3):311-316.
[2]付立東,王 宇,隋 鑫,等. 氮肥運籌對濱海鹽堿地水稻生育及產量的影響[J]. 沈陽農業大學學報,2010,41(3):327-330.
[3]Long X H,Zhao J,Liu Z P,et al. Applying geostatistics to determine the soil quality improvement by Jerusalem artichoke in coastal saline zone[J]. Ecological Engineering,2014,70(4):319-326.
[4]Sairam R K,Vasanthan B,Arora A,et al. Calcium regulates Gladiolus flower senescence by influencing antioxidative enzymes activity[J]. Acta Physiologiae Plantarum,2011,33(5):1897-1904.
[5]孫彩霞,劉志剛,荊艷東. 水分脅迫對玉米葉片關鍵防御酶系活性及其同工酶的影響[J]. 玉米科學,2003,11(1):63-66.
[6]Ennajeh M,Vadel A M,Khemira H,et al. Osmoregulation and osmoprotection in the leaf cells of two olive cultivars subjected to severe water deficit[J]. Acta Physiologiae Plantarum,2009,31(4):711-721.
[7]桑利敏. 甜菜幼苗對中性鹽和堿性鹽脅迫的生理應答特性[D]. 哈爾濱:東北農業大學,2017.
[8]師晨娟,劉 勇,荊 濤. 植物激素抗逆性研究進展[J]. 世界林業研究,2006,19(5):21-26.
[9]劉拴成,曹興明,穆俊祥,等. 外源激素對鹽脅迫下番茄種子萌發及幼苗生長的影響[J]. 種子,2016,35(12):94-98.
[10]李曉宇,穆春生. 鹽堿脅迫及外源植物激素對小麥和羊草生長發育的影響[J]. 草地學報,2017,25(2):257-260.
[11]Malamy J,Carr J P,Klessig D F,et al. Salicylic acid:a likely endogenous signal in the resistance response of tobacco to viral infection[J]. Science,1990,250(4983):1002-1004.
[12]Métraux J P,Signer H,Ryals J,et al. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber[J]. Science,1990,250(4983):1004-1006.
[13]Durner J,Shah J,Klessig D F. Salicylic acid and disease resistance in plants[J]. Trends in Plant Science,1997,2(7):266-274.
[14]Delaney T P,Uknes S,Vernooij B,et al. A central role of salicylic Acid in plant disease resistance[J]. Science,1994,266(5188):1247-1250.
[15]徐國瑞,劉濟明,閆國華,等. 羅甸小米核桃葉綠素含量測定方法研究[J]. 山地農業生物學報,2010,29(5):419-423.
[16]陳愛葵,韓瑞宏,李東洋,等. 植物葉片相對電導率測定方法比較研究[J]. 廣東教育學院學報,2010,30(5):88-91.
[17]王學奎. 植物生理生化實驗原理和技術[M]. 2版.北京:高等教育出版社,2006.
[18]李合生. 植物生理生化實驗原理和技術[M]. 北京:高等教育出版社,2000.
[19]陳建勛,王曉峰. 植物生理學實驗指導[M].廣州:華南理工大學出版社,2006.
[20]代其林,王金玲,馬明莉,等. NaCl脅迫處理對豇豆幼苗抗氧化酶活性的影響[J]. 江蘇農業科學,2015,43(10):193-196.
[21]Hou Y D,Guo Z F,Yi Y,et al. Effects of cold acclimation and exogenous pytohormone abscisic acid treatment on physiological indicators of winterness wheat[J]. Journal of Plant Sciences,2010,5(2):125-136.[HJ1.95mm]
[22]王連榮,薛擁志,常美花,等. 外源激素對杏扁抗寒生理指標的影響[J]. 核農學報,2016,30(2):396-403.
[23]孫小芳,鄭青松,劉友良. NaCl脅迫對棉花種子萌發和幼苗生長的傷害[J]. 植物資源與環境學報,2000,9(3):22-25.
[24]沈禹穎,王鎖民. 鹽脅迫對牧草種子萌發及其恢復的影響[J]. 草業學報,1999,42(3):54-60.
[25]Jini D,Joseph B. Physiological mechanism of salicylic acid for alleviation of salt stress in rice[J]. Rice Science,2017,24(2):97-108.
[26]付 艷,高樹仁,楊克軍,等. 鹽脅迫對玉米耐鹽系與鹽敏感系苗期幾個生理生化指標的影響[J]. 植物生理學報,2011,47(5):459-462.
[27]富春元,張淑娟,魚昭君,等. PEG模擬干旱脅迫對葉緣裂刻小白菜生理特性的影響[J]. 西北農業學報,2014,23(5):139-145.
[28]杜世章,代其林,奉 斌,等. 不同濃度NaCl脅迫處理下豇豆幼苗抗氧化酶活性的變化[J]. 基因組學與應用生物學,2011,30(3):351-356.
