蘇干湖濕地植被覆蓋度時空變化格局
2020-06-11 11:51:44康滿萍趙成章楊俊倉
生態學報 2020年9期
關鍵詞:生長
康滿萍,趙成章,*,白 雪,楊俊倉
1 西北師范大學地理與環境科學學院,甘肅省濕地資源保護與產業發展工程研究中心, 蘭州 730070 2 甘肅省地質環境監測院, 蘭州 730050
濕地是一種生態水文過程獨特、生態服務功能巨大的生態系統,氣候因子和土壤理化屬性的空間異質性是濕地植被生物量和覆蓋度形成的主要驅動力[1],對濕地植被群落的組成和優勢物種的空間分布具有重要影響[2]。植被覆蓋度是植被投影與該地域面積之比,不僅反映了植物群落的外貌和性質,而且維持著濕地植被群落結構和功能的穩定性[3],對于保持濕地景觀多樣性和棲息地環境安全性具有重要意義[4]。內陸鹽沼濕地是干旱和半干旱氣候下地表過濕或季節性積水、土壤鹽漬化并長有鹽生植物的沼澤濕地[5],在時間尺度上氣溫和降水等因子影響著植物群落的生態學過程和植物的生長周期[6],在空間上濕地植被的生長發育和優勢種群的分布更多地受到土壤全鹽量和地下水位埋深的影響[7],而植被生長狀態變化又將改變陸地地表下墊面屬性,進一步影響氣候調節、水土保持以及整個濕地生態系統的穩定性。內陸鹽沼濕地的植被覆蓋度在空間上具有明顯的斑塊化分布特征,傳統的群落學調查方法具有時空尺度上的局限性,遙感技術在小尺度區域存在精度誤差[8],將濕地群落學調查和遙感技術相結合能夠較好的刻畫鹽沼濕地植被群落蓋度的斑塊化和空間異質性,有助于認識內陸鹽沼濕地植物覆蓋度的時空變化及其成因,對于理解鹽沼濕地植物群落空間分布格局的形成機制具有重要意義。
蘇干湖濕地位于柴達木盆地西北部阿爾金山和祁連山系交會處的內陸盆地,是全球氣候變化的敏感區,植被覆蓋度對蘇干湖內陸沼澤濕地的生態環境的形成具有重要的作用。目前,像元二分模型已廣泛地應用于植被蓋度空間異質性的研究[9- 11],許多學者圍繞內陸鹽沼濕地的種群分布格局[12]、生物量分配[5]進行了深入研究,對蘇干湖濕地的群落組成[13]、鳥類多樣性[14]、生態脆弱性評價[15]、水循環的影響[16]等方面做了大量的研究。關于內陸鹽沼濕地植被覆蓋度空間分異規律及其成因研究還比較薄弱,特別是對蘇干湖濕地植被覆蓋度時空分布格局及其形成機理認識尚不清晰。鑒于此,本研究在濕地群落學調查的基礎上,基于landsat數據采用像元二分模型來估算蘇干湖濕地1987—2017年植被覆蓋度,分析了蘇干湖濕地各等級植被覆蓋度的變化格局及其影響因子。試圖明晰:(1)蘇干湖濕地近30年植被覆蓋度的年際變化特征及其與氣候因子間的關系;(2)不同等級植被覆蓋度濕地的空間分布格局及其地下水、土壤全鹽量間的關系,為提高蘇干湖濕地候鳥保護區生態環境的安全性提供理論依據。
1 研究區概況
蘇干湖自然保護區位于甘肅省酒泉市阿克塞哈薩克族自治縣阿勒騰鄉境內(93°47′53″—94°04′26″E, 38°50′07″—38°56′27″N),處于阿爾金山、黨河南山與賽什騰山之間的花海子-蘇干湖盆地的西北端,海拔2795—2808 m,大蘇干湖(咸水湖)水面面積約101.6 km2,小蘇干湖(微咸淡水湖)水面面積約11.6 km2。屬內陸高寒半干旱氣候,夏季短而涼爽,冬季長且寒冷,西北風盛行,日溫差較大,年平均氣溫3—6℃,年均降水量35.5—70.6 mm,蒸發量1228—2495 mm,該區大風天氣盛行,沙塵暴發生頻率高,干燥度為30,無霜期90 d左右。蘇干湖水系屬于柴達木內流水系中西北端的一個獨立水系,發源于黨河南山的大、小哈兒騰河出山口潛流于地下,在海子盆地成泉涌露,匯成河網,流入大蘇干湖和小蘇干湖,最終消耗于泉水溢出帶的蒸發蒸騰和湖面蒸發。土壤主要草甸土、草甸沼澤土及鹽堿土等,主要植物有:蘆葦(Phragmitesaustralis)、賴草(Leymussecalinus)、風毛菊(Saussureajaponica)、西伯利亞蓼(Polygonumsibiricum)、披針葉黃華(Thermopsislanceolata)、矮藨草(Scirpuspumilus)、海韭菜(Triglochinpalustre)、水麥冬(Triglochinmaritimum)、鹽角草(Salicorniaeuropaea)、乳苣(Mulgediumtataricum)、中間型荸薺(Heleocharisintersita)、苔草(Carexspp)、嵩草(Kobresiamyosuroides)等。
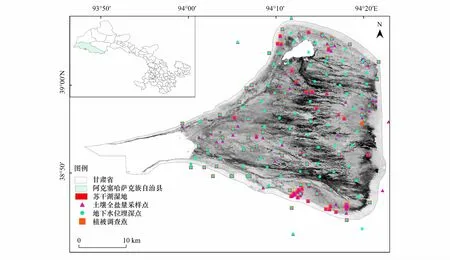
圖1 蘇干湖濕地實地調查數據采樣點Fig.1 Sampling points of field survey data of Sugan Lake wetland
2 數據來源與研究方法
2.1 數據來源
2.1.1濕地群落學調查
2017年8月下旬至9月上旬(群落生物量達到最大時)進行濕地群落學調查,在前期實地踏勘的基礎上,沿南北方向設置5條調查樣帶,在每條樣帶內間隔500 m設置1塊10 m×10 m的樣地,在每個樣地上設3個1 m×1 m小樣方,測定樣方內所有植物的種類、蓋度、高度和密度,重復3次。

2.1.2遙感數據
傳統的群落學調查具有時間和空間尺度上的局限性,在2017年濕地群落學調查的基礎上,結合遙感技術對該區域植被覆蓋度進行估算。本文選取蘇干湖濕地1987—2017年的遙感數據,來源于地理空間數據云(www.gscloud.cn)Landsat(Landsat- 5和Landsat- 8)衛星,研究區影像均采用9—10月植被生長旺盛季節、云量小于10%的數據,合計7景,其空間分辨率為30 m×30 m。利用ENVI 5.1對原始影像進行輻射定標,將其DN值轉化為像元輻射亮度值;以FLAASH模型對影像進行大氣校正,將輻射亮度值轉化為地表真實反射率;對影像進行幾何精校正,校正誤差控制在0.1個像元以內;采用Landsat- 5中的4、3、2波段,Landsat- 8中的5、4、3波段進行標準假彩色波段的融合,有助于歸一化植被指數(Normalized Difference Vegetation Index,NDVI)的計算,采用像元二分模型計算每期的植被覆蓋度。
2.1.3氣象數據
氣溫、降水資料均來源于中國氣象科學數據共享服務網(http://cdc.cma.gov.cn/),蘇干湖濕地數據來源周邊地區現有冷湖、敦煌、大柴旦、德令哈、肅北等5個國家氣象站的降水量、月平均氣溫等氣象數據作為研究數據。根據各氣象站點經緯度信息,采用ArcGIS 10.3軟件Geostatistical Analyst模塊對氣象數據進行Kriging空間插值,最終獲取每期氣象數據柵格圖像,通過數據掩膜,剪取蘇干湖濕地年平均溫度和年降水量的柵格圖像,獲取該區域年均氣溫和年均降水量的數據。
2.1.4地下水位埋深
地下水位埋深資料來源于甘肅省地質環境研究院2016—2017年在蘇干湖地區選取具有代表性的167個地下水位點(圖1),本文在ArcGIS 10.3軟件下將各點位的地下水埋深值進行Kriging空間插值,獲取地下水位埋深數據的柵格圖像,通過數據掩膜剪取蘇干湖濕地地下水位埋深的柵格圖像。
2.2 研究方法
2.2.1歸一化植被指數(NDVI)
歸一化植被指數(NDVI)是植被生長狀態及植被覆蓋度最佳指示因子,被定義為近紅外波段與可見光紅波段數值之差與這兩個波段數值之和的比值[17]。計算公式:
NDVI=(NIR-R)/(NIR+R)
(1)
式中,NIR和R分別為遙感圖像中近紅外和紅波段的反射率。
2.2.2植被覆蓋度的計算
基于混合像元分解理論的像元二分法能夠在較大程度上消除土壤等背景因素的影響,能高精度提取干旱地區純植被像元[18],客觀有效地反映植被冠層情況,采用像元二分模型,公式:
FVC=[(NDVI-NDVIsoil)/(NDVIveg-NDVIsoil)]
(2)
式中,FVC為研究區的植被覆蓋度,NDVI為歸一化植被指數,NDVISoil為完全是裸土或無植被覆蓋區域的NDVI值(即取累計頻率為0.5%的NDVI值為NDVISoil),NDVIVeg則代表完全被植被所覆蓋的像元的NDVI值(取累計頻率為99.5%的NDVI值為NDVIVeg),即純植被像元的NDVI值。
2.2.3植被覆蓋度的等級劃分
根據國家《土地利用現狀調查技術規程》和《濕地資源調查技術規程(2008)》,結合濕地群落蓋度、濕地類型和濕地植物組成等野外濕地調查資料,制定了蘇干湖濕地植被覆蓋度標準,將研究區分為5個蓋度組(表1)。
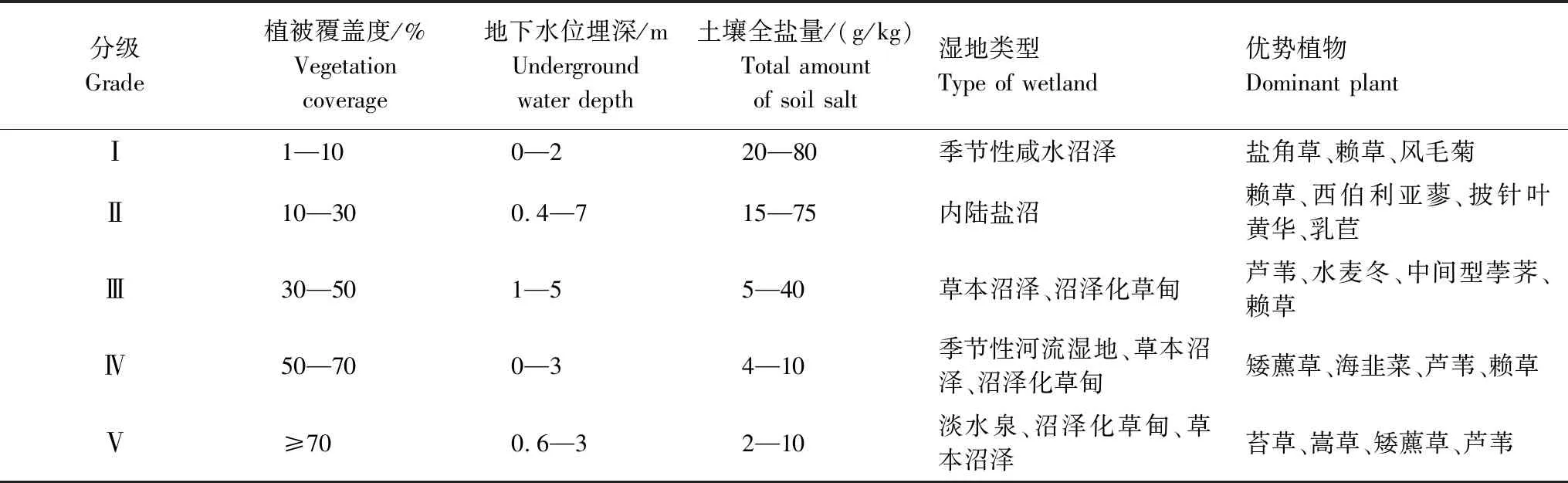
表1 蘇干湖濕地調查植被覆蓋度分級
Ⅰ:極低植被覆蓋度;Ⅱ:低植被覆蓋度;Ⅲ:中植被覆蓋度;Ⅳ:中高植被覆蓋度;Ⅴ:高植被覆蓋度
3 結果分析
3.1 植被覆蓋度的擬合度檢驗
根據landsat衛星遙感監測數據和群落學調查的植被覆蓋度實測值,建立植被覆蓋度的線性回歸模型,通過擬合程度來檢驗像元二分模型,圖2可以看出遙感反演結果與實測值趨于y=0.9108x+8.5696,其決定系數R2為0.822,相關性程度均較為滿意,線性模型擬合程度最高;表明基于遙感數據NDVI通過像元二分模型模擬得到的植被覆蓋度在蘇干湖內陸鹽沼濕地具有較高的模擬精度和可靠性。
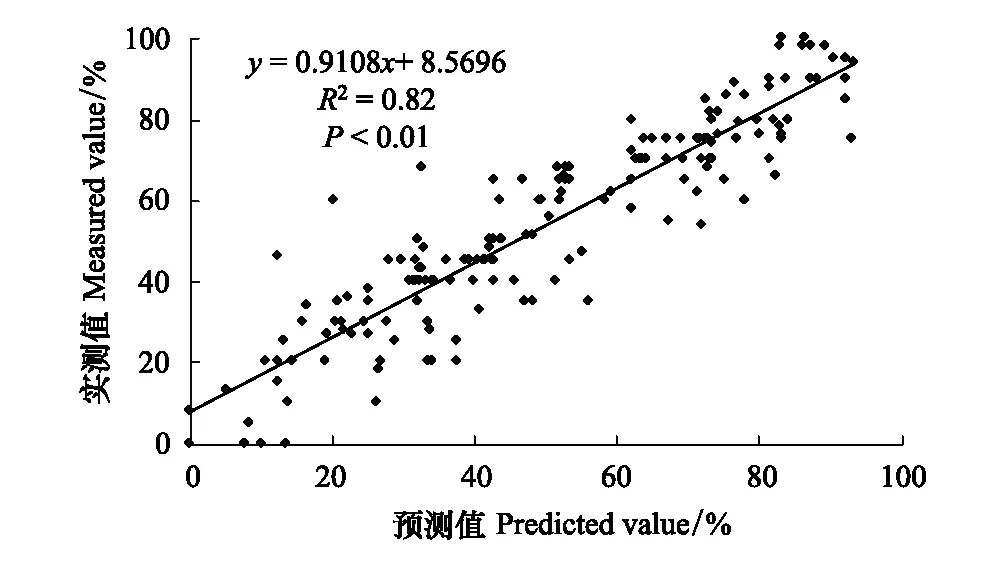
圖2 2017年植被覆蓋度實測值和預測值Fig.2 Measured and predicted vegetation coverage in 2017
3.2 植被覆蓋度的時間變化及其對氣候的響應
3.2.1植被覆蓋度年際變化
蘇干湖濕地植被覆蓋度的年際變化總體上逐年呈上升趨勢(圖3),其多年平均值為19.34%,年際變化率為0.162%5a-1,植被覆蓋度的最大值和最小值出現在2002年和2017年,分別為11.20%和31.22%。1987—2017年蘇干湖濕地植被覆蓋度的年際變化分三個階段,在1987—2002年間植被覆蓋度呈波動下降的變化,自2003年植被覆蓋度呈波動上升變化,尤其在2013—2017年間植被覆蓋度增長幅度較大。
3.2.2植被覆蓋度對氣溫、降水的響應
1987—2017年蘇干湖保護區的氣溫和降水呈上升趨勢(圖3),年際變化率分別為0.2℃5a-1和0.85 mm5a-1。多年平均氣溫為5.76℃,其波動范圍在4.5—6.5℃之間;多年平均降水量為36 mm,其年均降水量波動變化范圍在35.5—70.6 mm間,1987到2000年降水量呈波動減少,且低于多年平均降水量;2000年之后降水量波動增加且幅度較大,且高于多年平均水平,降水量最大值出現在在2002年為65 mm。
蘇干湖保護區氣溫與植被覆蓋度呈顯著正相關關系(P<0.05)(表2),相關系數為0.64,在高寒的氣候背景下,氣溫上升有利于植被的生長發育;植被覆蓋度與降水呈顯著正相關關系(P<0.05),相關系數為0.69,降水量的增加為干旱荒漠區的植被生長提供了必需的水分,利于植被覆蓋度的增加;降水量與植被覆蓋度的相關性系數大于氣溫,由此可知蘇干湖濕地的植被覆蓋度更多地受降水的影響更大。

圖3 1987—2017年蘇干湖濕地植被覆蓋度、氣溫和降水的年際變化Fig.3 Interannual variations of vegetation coverage, temperature and precipitation in Sugan Lake wetland from 1987 to 2017
表2 植被覆蓋度與氣溫、降水相關系數
Table 2 Coefficient of correlation between vegetation coverage and temperature, precipitation

相關系數Correlation coefficient植被覆蓋度/%Vegetation coverage氣溫/℃Temperature降水/mmPrecipitation植被覆蓋度Vegetation coverage/%1氣溫Temperature/℃0.63?1降水Precipitation/mm0.69?0.931
*表示通過置信度為0.05的顯著檢驗
3.3 植被覆蓋度的空間分布及其與影響因子間的關系
3.3.1植被覆蓋度空間分布

圖4 2017年蘇干湖濕地植被覆蓋度空間分布 Fig.4 Spatial and temporal changes of vegetation coverage in Sugan Lake wetland in 2007
蘇干湖保護區不同等級植被覆蓋度濕地呈斑塊化鑲嵌分布(圖4),2017年極低植被覆蓋度濕地的面積最小為39.36 km2,占比為3.92%,主要分布在微咸水沼澤濕地,在山地洪積扇的邊緣地帶也有少量分布;低植被覆蓋度濕地面積為437.58 km2,占比最大為43.53%,主要分布在大、小蘇干湖周邊、水鴨子河南的河流灘地及濕地中部的積水洼地;中植被覆蓋度濕地面積為242.26 km2,所占面積較大,呈條帶狀主要分布在泉水、河流周邊區域;中高植被覆蓋度所占面積較小,所占比為7.78%,呈零星分布在小蘇干湖以南和水鴨子河上游的周邊灘地;高植被覆蓋度濕地面積為101.42 km2,呈斑塊狀分布在小蘇干湖以南的淡水區和濕地的泉水出露地帶和水鴨子河下游的灘地。
3.3.2植被覆蓋度對地下水、土壤全鹽量的響應
地下水位的高低直接影響濕地植物的分布和物種多樣性,從根本上影響植物群落的分布格局[20]。蘇干湖濕地植被覆蓋度與地下水位埋深間呈較低負相關性,相關系數為-0.03,各等級濕地植被覆蓋度與地下水位間的相關性各有差異(圖5),其中:極低植被覆蓋度與地下水位埋深間呈極顯著正相關(P<0.01),相關系數為0.73;低植被覆蓋度與地下水位埋深間極顯著負相關(P<0.01),相關系數為-0.62;中、中高、高植被覆蓋度與地下水位埋深間呈較低的正相關性,相關系數分別為0.04、0.03和0.31,地下水位埋深在2 m左右,適宜植被的生長,整體上地下水位埋深對植被覆蓋度的影響較小。
蘇干湖濕地植被覆蓋度與土壤全鹽量呈極顯著負相關(P<0.01),相關系數為-0.73,隨著土壤全鹽量的增加濕地植被覆蓋度呈減少的趨勢,但各等級植被覆蓋度與土壤全鹽量間的相關性存在明顯差異(圖5)。其中:極低、低植被覆蓋度濕地分布區與土壤全鹽量呈極顯著負相關(P<0.01),相關系數分別為-0.6和-0.61,較高的土壤全鹽量抑制植被的生長。中、中高、高植被覆蓋度與土壤全鹽量間相關性較低,相關系數分別為-0.35、-0.48和-0.23,地表的土壤全鹽量受稀釋作用,土壤鹽漬化較輕,對植被的生長影響相對較小,植被覆蓋度越好。

圖5 植被覆蓋度與地下水位埋深、土壤全鹽量間的相關系數圖Fig.5 Coefficient of correlation between vegetation coverage and groundwater table depth and soil salinity in wetland content ***表示通過置信度為0.01的顯著檢驗,*表示通過置信度為0.05的顯著檢驗
4 討論
植被的生長發育和分布與其所處環境條件息息相關,氣候和環境因子在時空尺度上的穩定性,能夠反映植被群落的分布格局和植物生物量的分配模式[19]。鹽沼濕地植被對氣候變化的響應非常敏感,全球氣候變化影響植被的生長動態以及植被的結構和功能,地下水和土壤因子是植被覆蓋度空間分布差異的驅動因子[20]。為避免各因子間存在共線性,植被覆蓋度與環境因子間的偽相關的情況,采用偏相關的方法,剔除共線性因子的影響,通過皮爾遜相關分析方法得到該區域植被覆蓋度與各環境影響因子間的關系(如表3),本研究發現:蘇干湖濕地近30年植被覆蓋度的年際變化隨著氣溫和降水的增加呈上升趨勢,氣溫和降水與植被覆蓋度呈正相關關系,相關系數分別為0.43和0.58,且降水的相關系數高于氣溫;植被覆蓋度與地下水埋深的相關性較低,主要受該區域微地形、土壤質地等因素的復雜的水體的影響;土壤全鹽量是影響植被覆蓋度空間異質性分布的主要因子,與土壤全鹽量呈極顯著的負相關(P<0.01),相關系數為-0.826,不同等級植被覆蓋度濕地受地下水位埋深、土壤全鹽量的影響呈斑塊狀鑲嵌分布,其相關性具有差異性。

表3 植被覆蓋度與各環境因子間的相關系數
**相關性在0.01上顯著,*相關性在0.05上顯著
4.1 植被覆蓋度年際變化與氣溫、降水間的關系
氣溫和降水是決定植被空間分布及其變化的主要非生物因素,影響著植物群落的生態學過程和植物的生長周期[21]。蘇干湖保護區的多年平均氣溫為5.76℃,降水量為36 mm,屬于大陸性高寒氣候,為適應嚴酷的氣候環境,濕地植被的外在結構和主要植物的內在生理功能具有相對應的耐受性和抗逆性策略。近30年植被覆蓋度的呈波動增加趨勢(圖3),與氣溫、降水量呈顯著正相關(表2),主要原因有:(1)氣溫升高加快了周邊山地冰川消融,匯集地勢低平的盆地,稀釋土壤鹽分,有利于植被的生長;另外隨著氣溫上升,為濕地植被提供了充足的熱量,植物用于光合作用的光化學能增加,利于碳水化合物的合成和運輸,同時促進了根系對水肥的吸收和輸導功能,進而加快了植物的生長速率,提高了植被覆蓋度。(2)降水對干旱荒漠區的植被生長至關重要,降水量的增加提高了植被生長所需水資源的供給水平,有利于植被蓋度和生物量的增加;另一方面,蘇干湖濕地主要分布在花海子盆地,地表水和地下水的補給靠冰川消融、積雪融水、大氣降水、盆地東部、小哈爾騰河的強烈的入滲和兩側山區季節洪流的入滲補給以及基巖裂隙水的側向補給[16,22],隨著降水的增加,兩側山區的洪水和雨水在封閉的盆地聚集,為沼澤濕地的發育提供了豐富的水資源,促使濕生、鹽生植物大量繁衍,提高了濕地的植被覆蓋度。因此,近30年來蘇干湖濕地植被覆蓋度與氣溫、降水呈正相關關系,植被覆蓋度受降水的影響更大,這與李博等[9]的研究相一致。
4.2 植被覆蓋度空間異質性及其影響因子分析
4.2.1地下水埋深對植被覆蓋度空間異質性的影響
水是鹽沼濕地生態系統中最為敏感的環境因子,地下水位的高低直接影響植物分布和物種多樣性,從根本上影響植物群落的分布格局[23]。蘇干湖濕地植被覆蓋度與地下水位埋深間的相關性較低,不同等級植被覆蓋度與地下水位埋深間的相關性各有差異(圖5),極低植被覆蓋度濕地主要以分布在泉水出露地帶的季節性咸水沼澤濕地為主,鹽角草、風毛菊等鹽生植物是蘇干湖季節性咸水沼澤濕地的優勢物種(表1),因長時間受水淹的影響,植物生長期短、植株矮小,植被稀疏且分布零散,地下水位越淺,地表積水越多,植被覆蓋度越低;隨著地下水位埋深的增加,濕地表面靜水持留時間減少,生長狀況好轉,植被覆蓋度增加,使濕地植被覆蓋度與地下水位埋深之間形成了正相關關系(圖5)。低植被覆蓋度濕地主要分布在湖水、河流周邊灘地及積水洼地的內陸鹽沼濕地,該類濕地以賴草、西伯利亞蓼、披針葉黃華、乳苣等植被群落為主(表1、圖4),這些植被雖具有耐干旱、耐貧瘠的特性,但是隨著地下水位埋深的增加,植株根系吸水困難,植被的生長受到抑制,植被覆蓋度與地下水位間呈負相關。中、中高、高植被覆蓋度濕地主要分布在泉水、河流周邊的沼澤化草甸和季節性河流濕地,這些濕地呈斑塊狀分布,在地表流動性淡水的滋養下,以蘆葦、水麥冬、中間型荸薺、賴草、矮藨草、海韭菜、苔草和嵩草等植被群落為優勢種群,植被蓋度與地下水位埋深間呈不顯著相關(圖5)。該區域的植被隨年齡增長和種群密度、蓋度的增加,優勢種群對種內種間競爭可利用水資源的能力不斷提升,其生存能力不斷提高,種群規模不斷擴大,自身優勢地位得到了鞏固,其根莖較深、植株個體的高度、生物量較大,生長繁茂,蓋度相對較高[24-25]。因此,地下水位的高低直接影響植被的生長和物種的多樣性,地下水位埋深對植被的生長有適宜的閾值,水位埋深過淺或過深會威脅植被群落的正常生長,這與徐海量等[23]的研究相一致。
4.2.2土壤全鹽量對植被覆蓋度空間異質性的影響
干旱區地下水的動態變化以及強烈的蒸發作用,加快了鹽分在土壤中的移動和積累,形成了土壤鹽分的空間異質性,進而影響內陸鹽沼濕地植被的生長及其空間分布格局[3,26]。蘇干湖濕地植被覆蓋度與土壤全鹽量呈顯著負相關,總體而言,隨著土壤全鹽量的降低,植被覆蓋度呈增加趨勢(表1、圖5),但是不同等級植被覆蓋度濕地與土壤全鹽量間的相關系有較大差異。極低、低植被覆蓋濕地主要分布在湖水、河流周邊的灘地,由于季節性咸水沼澤濕地和鹽沼濕地區域的淺層地下水受地表較強蒸發力的影響,土壤鹽分在地表聚集,土壤鹽堿化嚴重,較高土壤全鹽量可能引起根系腐爛,造成植被吸水能力和給養能力的減弱,甚至導致植物體內水分外滲或者導致植物死亡[27],分布在該濕地區域的鹽生植物植株矮小、葉面積小,植被覆蓋度與土壤全鹽量間呈負相關(圖5)。中、中高、高植被覆蓋度濕地主要分布于泉水、河流周邊區域,由于濕地表面常年或大部分時間有流動的淡水,土壤含鹽量較低(表1),有利于植被生長,是蘇干湖保護區植被生長狀況最好的區域,濕地植被覆蓋度與土壤全鹽量間的存在相關關系(圖5)。全區濕地群落的蓋度隨著土壤鹽分含量的減少呈增加趨勢,受水鹽環境脅迫較小的區域,優勢種的生存能力不斷提高,種群規模不斷擴大,植株較高,生長茂盛,植被蓋度增加;競爭能力較弱的物種無法占據優良生境而衰退,只能在優勢種無法生存的區域定居,種群單一且其生長受水鹽等環境因子的脅迫,其植株矮小,覆蓋度相對較低[28-29]。因此,土壤全鹽量的高低直接影響植被生態系統的結構、組成和分布,土壤中鹽分過高時,引起植被根系腐爛造成對吸收水分和養分的能力減弱,從而對植被產生滲透脅迫,影響其正常的生長發育,這與李亮[27]的研究相一致。
5 結論
本研究發現,在高寒的氣候背景下,氣溫上升有利于植被的生長發育,而降水對植被的影響更大。植物的水鹽適應能力與植物形態結構和其空間分布格局密切相關,水、鹽的變化影響濕地群落的蓋度、種群間競爭能力和植株之間的鄰體干擾。蘇干湖濕地植被覆蓋度空間分布受地下水位埋深和土壤全鹽量空間異質性的影響呈斑塊狀分布,與地下水位埋深、土壤全鹽量呈負相關;不同等級植被覆蓋度與地下水位埋深、土壤全鹽間的相關性各不相同。地下水位埋深對植被的生長有適宜的閾值,其過淺或過深會威脅植被群落的正常生長;土壤全鹽量是影響該區域植被分布的主要影響因子,土壤全鹽量較高植被根系腐爛造成對吸收水分和養分的能力減弱,鹽生植物植株矮小。內陸鹽沼濕地植被覆蓋度與氣候、水分、土壤鹽分等因子在時間和空間上表現出相對的穩定性和空間異質性,今后應分區討論各植被群落及其植被覆蓋度的空間格局與其他環境要素間的關系,對未來分析評價蘇干湖濕地的生態環境具有重要現實意義。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
